Late Maastrichtian-Danian Mytilids and Pinnids
(Mollusca: Bivalvia) from Northern Patagonia, Argentina
Miguel Griffin1,2, Ana Parras1,2 and Silvio Casadío1,2
1Facultad de Ciencias Exactas y Naturales, Universidad Nacional
de La Pampa, Uruguay 151, 6300 Santa Rosa, La Pampa,
Argentina
2Consejo Nacional de Investigaciones Científicas y Técnicas,
Argentina. miguelgriffin@aol.com
Abstract. The Upper Cretaceous - lower Paleogene rocks from northern Patagonia yield a rich fauna of
marine mollusks that may be useful in elucidating paleobiogeographic and paleoenvironmental changes
taking place across the K/P boundary in the region. Most of the mollusk faunas remain poorly understood,
as the preservation is generally deficient. This paper describes three new species of mytilids
(Septifer mayai n. sp., Mytilaster ? edseli n. sp. and Gregariella amara n. sp.), and records four species of pinnids,
one of which is new (Plesiopinna pampeana n. sp.). All the material comes from the Maastrichtian
Jagüel Formation, and the Maastrichtian/Danian Roca Formation, exposed in the provinces of Río Negro,
Mendoza and La Pampa (Argentina). A survey of the stratigraphic distribution of the material described
shows that the only Maastrichtian species that survived into the Danian was Gregariella amara.
Resumen. Mytílidos y Pínnidos (Mollusca: Bivalvia) del Maastrichtiano Tardío - Daniano del Norte
de Patagonia, Argentina . Las rocas del Cretácico Superior-Paleógeno inferior expuestas en el norte de
Patagonia contienen una rica fauna de moluscos cuyo estudio es importante para comprender los cambios
paleobiogeográficos y paleoambientales producidos en la región durante el límite K/P. Sin embargo, una
porción considerable de esta fauna permanece poco estudiada, probablemente a causa de su pobre preservación.
En este trabajo se describen tres especies nuevas de mytílidos (Septifer mayai n. sp., Mytilaster ? edseli
n. sp. y Gregariella amara n. sp.) y cuatro especies de pínnidos de las cuales una es nueva (Plesiopinna
pampeana n. sp.). Todos los ejemplares provienen de las formaciones Jagüel (Maastrichtiano) y Roca
(Maastrichtiano/Daniano) expuestas en las provincias de Río Negro, La Pampa y Mendoza, Argentina.
Gregariella amara es la única especie que se registra en el Maastrichtiano y Daniano.
Key words. Bivalvia; Mytilids; Pinnids; Cretaceous/Paleogene boundary; Argentina.
Palabras clave. Bivalvia; Mytílidos; Pínnidos; Límite Cretácico/Paleógeno; Argentina.
Introduction
Upper Cretaceous and Paleogene marine faunas
from Patagonia are known since the end of the nineteenth
century and early part of the twentieth century,
particularly from Northern Patagonia
(Burckhardt, 1900, 1901; Ihering, 1903, 1907, 1914;
Böhm, 1903; Fritzsche, 1919; Weaver, 1931; and references
therein). However, most of the species were
based on insufficient material, whether because the
available specimens were few or else poorly preserved,
a fact that hinders taxonomic, paleobiogeographic,
and biostratigraphic inferences. Although
true that the material is generally poor, extensive collections
during the last few decades have allowed a
better understanding of the taxonomic composition
of the fauna and of its paleobiogeographic relationships
(Camacho, 1970; Casadío, 1998; Casadío et al.,
2005; Griffin et al., 2005; and references therein).
Micropaleontological studies - mainly on ostracods
and foraminifers - were published by Bertels (1968,
1969, 1970), Concheyro and Villa (1996), and
Malumián and Caramés (1995). On this occasion we
describe seven taxa collected at different localities of
the Neuquén Basin and belonging in the Order
Mytiloida, a group which had remained until now
largely unnoticed in the Cretaceous/Paleogene rocks
of the area. The material described comes from beds
of well constrained Maastrichtian and Danian age; at
some of the localities the K/P boundary is exposed.
Therefore, in addition to aiding in the elucidation of
the paleobiogeographic relationships of the fauna,
these taxa also become valuable tools in understanding
the faunal events occurring at the K/P boundary.
Geological setting
During the Cretaceous-Paleogene transition, wide
areas of northern Patagonia were covered by a shallow
sea developed as a consequence of relative sea
level changes related to subsidence along the
Atlantic margin.
In the Neuquén Basin, in northwestern Patagonia,
the K/P boundary is recorded within rocks included
in the Malargüe Group, which includes from bottom
to top the Loncoche, Allen, Jagüel, Roca, Pircala and
El Carrizo Formations. These units are well exposed
in southern Mendoza, northeastern Neuquén, western
La Pampa and northern Río Negro. Because these
rocks provide an excellent opportunity to study the
changes operated in mid-latitude continental and
marine ecosystems of the southern hemisphere at the
end of the Cretaceous, they have been lately subject
to renewed interest (Casadío et al., 2005).
The Cretaceous - Paleogene boundary coincides
with an important change in the composition of the
marine invertebrate assemblages, consisting in an increase
in the number of species from low latitudes,
and a decrease of the endemic taxa with austral
affinities. This modification in the faunal composition
was coeval with an increase of the relative sea
level and the beginning of carbonatic sedimentation
(Feldmann et al., 1995; Casadío et al., 2005).
The studied fossils come from beds referred to the
Jagüel and Roca formations exposed in southern
Mendoza, western La Pampa, and northern Río Negro
(figures 1 and 2). The Maastrichtian Jagüel Formation
comprises mainly siltstone, and transitionally
grades into the limestones of the Roca Formation,
which has been referred to the Maastrichtian-Danian
(Casadío et al., 1998; Parras et al., 1998). Overlying the
Roca Formation are evaporitic deposits of the El
Carrizo Formation, while in the Andean area it gradually
gives way to the continental sandstone of the
Pircala Formation.
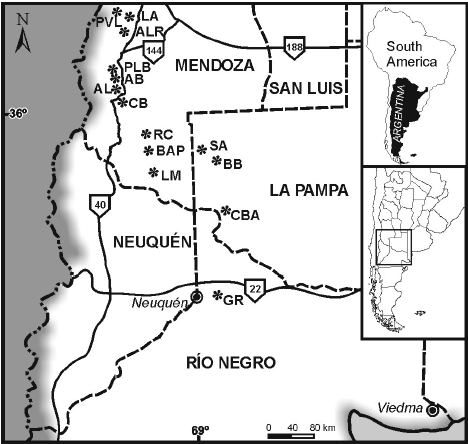
Figure 1. Map showing fossil localities / mapa mostrando las localidades fosilíferas. PVL, Portezuelo Vega del Loro; LA, Laguna
Amarga; ALR, Arroyo las Ramaditas; PLB, Puesto La Bebida; AB. Arroyo Brea; AL, Arroyo Loncoche; CB, Cerro Butaló; RC. Ranquil-Có; BAP, Bordo Alto del Payún; LM, Liu Malal; SA, Salitral de La Amarga; BB, Barda Baya; CBA, Cerros Bayos; GR, General Roca.
Figure 2. Stratigraphic sections at localities studied, showing lithology and fossil-bearing beds; vertical scale in meters / secciones estratigráficas de las localidades estudiadas, mostrando la litología y los niveles fosilíferos; escala vertical en metros.
The specimens studied are housed in the Departamento
de Ciencias Naturales of the Universidad
Nacional de La Pampa (GHUNLPam), Muséum
National d'Histoire Naturelle, Paris (MNHN-R), and
Naturhistoriska Riksmuseet, Stockholm (SMNH).
Systematic paleontology
Order MYTILOIDA Férussac, 1822
Superfamily MYTILOIDEA Rafinesque, 1815
Family MYTILIDAE Rafinesque, 1815
Subfamily MYTILINAE Rafinesque, 1815
Genus Septifer Recluz, 1848
Type species. Mytilus bilocularis Linné, 1758; subsequent designation
by Stoliczka, 1871.
Remarks. According to Soot-Ryen (1955), Septifer is
easily separated from the similar Hormomya Mörch,
1853 (type Mytilus exustus Linné, 1758), by the presence
of a deck or septum for the insertion of the anterior
adductor muscle. Except for differences in the
interior muscle insertion patterns, Septifer and
Hormomya share the same kind of ornamentation on
the outer shell surface, especially on the ventral area,
where the unilateral furcating ribs are readily distinct
from the bifurcating pattern seen in the otherwise externally
very similar Brachidontes Swainson, 1840
(type Modiola sulcata Lamarck, 1819).
Septifer is known to occur in rocks as early as
Triassic, and is nowadays widespread in tropical and
subtropical seas (Soot-Ryen, 1955; Beu and Maxwell,
1990). Recent species of Septifer inhabit upper subtidal
zones attached to rocks or shell debris (Selin and
Latypov, 2006).
Several taxa mentioned in the geological literature
as Mytilus Linné, 1758 (type Mytilus edulis Linné,
1758), may probably belong better in Septifer, such as
the occurrences reported by Bodenbender (1892),
who recorded Mytilus cuvieri Matheron, 1843, in the
limestones of the Roca Formation exposed in Arroyo
Pequenco, west of Malargüe. Similarly, Weaver
(1927, p. 430; 1931, p. 89) mentioned Mytilus sp. from
the same unit exposed southeast of Cerro Auca Ma-
huida (province of Neuquén) and along the eastern
limb of the Malargüe syncline (province of Mendoza);
this record is also probably referable to Septifer.
Septifer mayai n. sp.
Figures 3.1-3

Figure 3. 1-3, Septifer mayai n. sp. 1, right valve, holotype / valva derecha, holotipo, GHUNLPam 808; 2, right valve / valva derecha, GHUNLPam 809; 3, ventral view / vista ventral, GHUNLPam 806. 4, Mytilaster ? edseli n. sp. left valve, holotype / valva izquierda, holotipo, GHUNLPam 10143. 5-8, Gregariella amara n. sp. 5, right valve / valva derecha, GHUNLPam 593. 6, left valve / valva izquierda,
GHUNLPam 586. 7, right valve, holotype / valva derecha, holotipo, GHUNLPam 592. 8, right valve / valva derecha, GHUNLPam 606. Scale
bar 1 cm / escala gráfica 1 cm.
Diagnosis. Average-sized Septifer, with very strong
postumbonal keel and 35 to 45 simple, flat, quadratesectioned,
smooth ribs separated by deeply incised
interspaces showing scaly ornamentation produced
by commarginal growth lines.
Localities. Barda Baya (36º 54' S, 67º 55' W) and
Salitral de La Amarga (36º 33' S, 68º 9' W), both in
the province of La Pampa; Liu Malal (36º 45' S, 68º
43' W), Bordo Alto del Payún (36º 31' S, 69º 27 W),
Ranquil Có (36º 12' S, 69º 30' W), Puesto La Bebida
(35º 26' S, 69º 44' W), and Portezuelo Vega del Loro
(34º 44' S, 69º 39' W), all in the province of Mendoza.
Stratigraphic range. All the material comes from the
late Maastrichtian Jagüel Formation and the
Maastrichtian part of the Roca Formation.
Material. Thirteen variably preserved specimens, generally as calcite
replacements of original shell. GHUNLPam 806-809; 1177-
1178; 10764; 10803; 15192-15193; 16049; 16582; 17170.
Holotype. Specimen GHUNLPam 808; all other specimens are
paratypes.
Type locality. Salitral de La Amarga (36º 33' S, 68º 9'
W), province of La Pampa.
Description. Shell mytiliform, small (largest specimen
42 mm long and 16 mm high); umbones terminal,
sharp; dorsal and posterior margins merged into
a wide uninterrupted curve that ends abruptly at
junction with ventral margin; anteroventral margin
concave; area next to posterior margin flat; ventral
area of shell enclosed by postumbonal ridge conspicuously
concave, deep; greatest shell width at central
area near ventral margin; byssal slit well developed;
shell surface covered by 35 to 45 radial flat, smooth
and quadrate-sectioned radial ribs; ribs generally
simple but occasionally bifurcating near dorsal and
posterior margins; intercalated radial ribs appearing
sometimes after growth stops marked by commarginal
lines; intercostal spaces narrower than ribs,
deeply incised, with scaly ornamentation (visible on
well preserved specimens) resulting from the fine
commarginal growth lines.
Measurements. Holotype GHUNLPam 808, length:
23 mm, height: 13.5 mm, width: 11 mm.
Remarks. Septifer mayai n. sp. is very similar to the material
described by Dartevelle and Freneix (1957, p. 49)
as Brachidontes ? charmesi (Péron, 1890) from the Cenomanian-
Senonian of Bulu-Zambi in Congo (pl. 6,
fig. 8) and the Senonian of Logbatjeck in Cameroon
(pl. 6, fig. 9). Questionably placed in Brachidontes, the
species described by Péron probably fits better in Septifer
(Dartevelle and Freneix, 1957). Examination of the
African specimens (MNHN-R 51397), which were collected
from Upper Cretaceous rocks in Cameroon, allowed
recognition of the following differences: smaller
size, less concave ventral margin and radial ribs bifurcating
at about half their length from the umbones.
Age. Late Maastrichtian. Zone CC26 (late Chron 30N
- Chron 29R).
Etymology. After Domingo "Cocho" Maya, in recognition
of his help during the field work in the Barda
Baya area, province of La Pampa.
Genus Mytilaster Monterosato, 1883
Type species. Mytilus lineatus Gmelin, 1791; original designation.
Remarks. The type species of Mytilaster is very similar
in outline and ornamentation to the specimens
from the Roca Formation, although the presence of
dysodont teeth behind the ligament could not be observed
in our material because no interiors are available.
Therefore, generic placement is subject to confirmation.
Fossil records of the genus are not common
and probably need further taxonomic work in
order to confirm them. It is known from the Miocene
of Poland (Brzozowska, 2004), and Recent species of
the genus, such as Mytilaster lineatus and Mytilaster
minumus (Poli, 1795), inhabit brackish waters of the
eastern Mediterranean, Black, and Caspian seas.
Mytilaster ? edseli n. sp.
Figure 3.4
Diagnosis. Small Mytilaster ? with very strong postumbonal
keel; anterior flank almost vertical and
completely smooth except for commarginal growth
lines; posterior area covered by c. 65 narrow and
rounded ribs separated by shallow interspaces about
one third the width of ribs.
Localities. Arroyo Loncoche (35º 41' S, 69º 40' W),
Arroyo Brea (35º 31' S, 69º 43' W), and Arroyo Las
Ramaditas (34º 54' S, 69º 32' W), all in the province of
Mendoza.
Stratigraphic range. All the material comes from the
Maastrichtian part of the Roca Formation.
Material. Six specimens, variably preserved. GHUNLPam 10035,
10143-10144; 15093-15094; 15097.
Holotype. GHUNLPam 10143; the rest of the specimens are
paratypes.
Type locality. Arroyo Brea (35º 31' S, 69º 43' W),
province of Mendoza.
Description. Shell small, reaching up to 28 mm in the
largest specimen, but generally around 15-18 mm,
mytiliform, with terminal umbones and a slight anteroventral
lobe; strong postumbonal ridge; anterior
part of dorsal margin straight, joining the almost
straight to very slightly convex posterior part of dorsal
margin at shell mid-length forming an angle of c.
110º; ventral margin slightly concave and posteroventral
end rounded; greatest width of shell at
about one third of height; outer shell surface divided
into two distinct areas: anterior area smooth and only
carrying fine commarginal growth lines, posterior
area beginning just in front of postumbonal ridge
and covered by c. 65 fine rounded ribs with shallow
interspaces that are never more than one third the
width of ribs; commarginal ornamentation of very
fine growth lines and occasionally a few growth
ledges.
Measurements. Holotype GHUNPam 10143,
length: 18 mm, height: 10 mm.
Remarks. As stated above, the similarity in outline
and ornamentation of this species with Mytilus lineatus
is striking. However, further material may prove
that the generic placement lies elsewhere. A species
described by Stoliczka (1871, p. 379-380, pl. 23, fig, 4,
6, and 7) as Modiola (Brachydontes) radiatula Stoliczka,
1871, from the Arrialoor Group near Comarapolliam,
southern India, is quite similar to our material. It
shows the same shell shape and ornamentation pattern
as Mytilaster ? edseli n. sp. and probably belongs
in Mytilaster too, as suggested by the anterior lobe
and radial ornamentation, but the radial ribs appear
to be weaker and more numerous, and the commarginal
growth lines are stronger. In addition, in the
Indian species the postumbonal ridge is weaker.
Age. Late Maastrichtian. Zone CC26 (late Chron 30N
- Chron 29R).
Etymology. This species is named after Edsel Brussa,
paleontologist at the Universidad Nacional de La
Pampa, Argentina.
Subfamily CRENELLINAE Adams and Adams, 1857
Genus Gregariella Monterosato, 1883
Type species. Modiolus sulcatus Risso, 1826 (= Modiola opifex Say,
1825); by monotypy.
Remarks. The genus Gregariella includes a number of
species closely resembling Musculus Röding, 1798
(type Mytilus discors Linné, 1758). The two genera differ
mainly in the pattern of ornamentation. While
both share valves with two radially ribbed areas at
each end and a smooth central area, in Gregariella the
first few ribs of the posterior area do not reach the
ventral margin, which they do in Musculus. In addition,
the postumbonal ridge is notably more marked
in Gregariella, which produces strongly keeled shells.
This contrasts shells of Musculus, which are generally
flatter as the postumbonal keel is poorly developed
or else it is entirely missing. Gregariella has been
described from rocks dated as far back as Eocene
(Beu, 2004).
Gregariella amara n. sp.
Figure 3.5-8
Diagnosis. Gregariella with well defined postumbonal
ridge; anterior area with 7-10 ribs and posterior
area with about 65 to 80, less crowded below postumbonal
ridge (18-20) than on dorsal area; rib section
gently convex.
Localities. Cerros Bayos (37º 40' S, 67º 30' W) and
Salitral de La Amarga (36º 33' S, 68º 9' W), province
of La Pampa; General Roca (39º 00' S, 67º 32' W),
province of Río Negro; Arroyo Brea (35º 31' S, 69º 43'
W), province of Mendoza.
Stratigraphic range. This species occurs within the
topmost beds of the late Maastrichtian Jagüel
Formation, and also in the overlying Maastrichtian-
Danian Roca Formation.
Material. Forty moderately well preserved specimens, most of
them as internal moulds but partially covered by calcite replacements
of the original shell. GHUNLPam 586-616, 4894; 4924-4925;
5226-5227; 10141-10142; 10157-10158.
Holotype. Specimen GHUNLPam 592 is designated holotype; the
rest of the specimens are paratypes.
Type locality. Salitral de La Amarga (36º 33' S, 68º 9'
W), province of La Pampa.
Description. Shell small (ranging between 7.5 and 25
mm long, but most of them about 18 mm), subovate,
umbones sub-terminal; anteroventral margin
straight to gently convex at mid-length; anterior dorsal
margin straight and ascendant; dorsal posterior
margin convex; posterior margin rounded; greatest
shell width at mid-length of shell; well defined postumbonal
ridge; shell ornamentation of fine radial
ribs covering anterior and posterior areas of shell; anterior
area with 7-10 ribs and posterior area with
about 65 to 80, less crowded below postumbonal
ridge (18-20) than on dorsal area; ribs on most dorsal
part of the shell deflected upwards to meet dorsal
margin at a nearly straight angle; few intercalated
ribs present; central area smooth; rib section gently
convex; first few ribs of posterior area not reaching
ventral margin; intercostal spaces shallow, as wide as
ribs.
Measurements. Holotype GHUNLPam 592, length:
16.6 mm, height: 8.6 mm, width: 7.0 mm.
Remarks. Gregariella amara n. sp. appears to be one of
the earliest members of the genus in the Western
Hemisphere. In southern South America, the earliest
representative known up to now is Modiola andina Ortmann, 1902 (p. 122, pl. 24, fig. 4; Frassinetti and
Covacevich, 1999, p. 21, pl. 3, fig. 2-3) from the late
Oligocene-early Miocene Centinela Formation exposed
just South of Lake Pueyrredón, province of
Santa Cruz, Argentina. Ortmann's species is very
similar and probably belongs in Gregariella too, but it
appears to be slightly larger, with a sharper portumbonal
keel, and the radial ribs are less numerous and
restricted to the posterior end only. Another probable
member of this genus is Modiola aprilis Feruglio,
1935 (p. 67, pl. 1, fig. 5.; Feruglio, 1936, p. 211, pl. 21,
fig. 7; Camacho, 1992, p. 37, tab. 1), from the Danian
Salamanca Formation, a unit exposed along the
Atlantic coast of central Patagonia just north of
Comodoro Rivadavia. However, identification of
Feruglio's material is at least problematic. The specimens
were not available for study and his original
description is too imprecise to warrant any certainty
in the taxonomic placement of his material, however
much the general shape resembles other taxa from
approximately coeval deposits in this part of South
America as is the case of Gregariella amara. New collections
from the Salamanca Formation may eventually
help to clarify the status of Feruglio's species.
According to the original description, Brachidontes
bifurcatus Popenoe, 1937 (p. 383, pl. 46, fig. 2) from
the Upper Cretaceous Williams Formation in
Southern California can be placed in Gregariella, as it
shows the same pattern of ornamentation, although
the hinge details remain unknown. While similar to
the Patagonian species, the ribs are wider, fewer and
more strongly marked. The shell is also significantly
larger, reaching up to twice the length.
Modiolus kirkerensis Clark, 1918 (p. 133, pl. 9, fig. 8;
Moore, 1983, p. A68, pl. 17, fig. 9) from the Miocene
San Ramon Sandstone in central California resembles
the Patagonian species in sculpture, although the anterior
part of the shell is rather poorly preserved and
thus no traces of the sculpture are preserved there.
However, the shell is much larger in the Californian
species. Moore (1983, p. A68) placed Clark's species
doubtfully in Brachidontes. Such a placement seems to
be incorrect but only further study will allow a correct
generic assignment.
Modiolus pontotocensis del Valle and Medina, 1980
(p. 53-54, pl. 2, fig. 3-5) from the Upper Cretaceous
López de Bertodano Formation exposed at Cape
Lamb on Seymour Island (Antarctica), is similar to Gregariella amara in its shell outline, relatively low
umbonal ridge, and radial ornamentation. However,
the ribs are weaker and less distinct than in the
northern Patagonian species and the shells are considerably
larger, reaching a length of 42 mm.
Modiolus cf. M. pontotocensis del Valle and Medina
sensu Zinsmeister and Macellari, 1988 (p. 263-264,
Fig. 8.14 -15), from unit 9 (late Maastrichtian) of the
López de Bertodano Formation, has more evenly
convex shells. The ornamentation is not apparent in
the available figures, but the authors mention the
presence of radial ribs, albeit not whether they are all
over the surface or restricted to the anterior and posterior
areas only.
Another Antarctic species that resembles Gregariella
amara -at least superficially- is Arcuatula sootryeni
Stilwell and Zinsmeister, 1992 (p. 58, pl. 3, figs
c, e, and g), from the Eocene La Meseta Formation exposed
in Seymour Island. The radial ribs and the general
shell outline -similar to Gregariella amara - suggest
it may fit in Gregariella.
An earliest Eocene specimen described by Tessier
(1952, p. 317, pl. 19, fig. 5-6) as Musculus elegans
(Sowerby, 1812) and collected in Kade-Baldji,
Senegal, appear to be quite similar to Gregariella amara.
They differ in the shell proportions, as the African
specimen seems to be higher and with a wider anterior
end. The ornamentation is also different, as the
specimen described by Tessier has 12 ribs on the anterior
area and those on the posterior are fewer,
stronger and the intercostals spaces are wider.
Age. Late Maastrichtian-Danian. Zone CC26 to Zone
NP4 (late Chron 30N - Chron 26R).
Etymology. Latin for "amarga" (bitter), after Salitral
de La Amarga, the type locality.
Superfamily PINNOIDEA Leach, 1819
Family PINNIDAE Leach, 1819
Genus Pinna Linné, 1758
Type species. Pinna rudis Linné, 1758; subsequent designation by
Children, 1823.
Remarks. According to Turner and Rosewater
(1958), Rosewater (1961), Packard and Jones (1965),
and Cox and Hertlein (1969), Pinna is characterized
by equivalve wedge-shaped shells, anteriorly
placed umbones, medial ridge, radial ribs, interior
nacreous layer divided into two distinct lobes by a
longitudinal groove, and the posterior adductor
muscle scar placed within the posterior nacreous
lobe. Species of Pinna are generally distinguished
from each other based on the general shell outline,
apical angle, strength of the medial ridge, and characteristic
ornamentation of the outer shell surface.
The genus is known from rocks as early as
Carboniferous and fossil species show a cosmopolitan
distribution. Recent representatives of this
group of suspension feeding bivalves are restricted
to tropical and subtropical shallow seas. Extant
species live with the anterior end buried in a soft
substrate and attached to particles of sediment by
means of well developed byssal threads (see Yonge,
1953; Stanley, 1970; Moore, 1983; Butler et al., 1993;
Richardson et al., 1999, for further details on the life
mode of pinnids).
Pinna sp. aff. P. cretacea (Schlotheim, 1813)
Figure 4. 1

Figure 4. 1, Pinna sp. aff. P. cretacea Schlotheim, right valve of slightly compressed specimen / valva derecha de un especimen algo comprimido,
GHUNLPam 15314. 2, Pinna sp., right valve / valva derecha, GHUNLPam 2913. 3-6, Plesiopinna pampeana n. sp. 3, posterior
view of shell broken perpendicular to hinge, showing flattened dorsal area, holotype / vista posterior de la conchilla mostrando el area dorsal aplanada, holotipo, GHUNLPam 810. 4, posterior view of obliquely broken shell, showing flattened dorsal area / vista posterior de la conchilla mostrando el area dorsal aplanada, GHUNLPam 811. 5, left valve / valva izquierda, GHUNLPam 811. 6, right valve, holotype / valva derecha, holotipo, GHUNLPam 810. 7, Atrina sp., right valve / valva derecha, GHUNLPam 17308. Scale bar 1 cm / escala gráfica 1 cm.
Localities. Cerro Butaló (35º 50' S, 69º 40' W), Arroyo
Brea (35º 31' S, 69º 43' W), Arroyo Las Ramaditas (34º
54' S, 69º 32' W), and Laguna Amarga (34º 46' S, 69º
33' W), all in the province of Mendoza.
Stratigraphic range. All specimens come from the
Roca Formation, which is late Maastrichtian at all
four localities.
Material. Five specimens, internal moulds of anterior ends showing
fragments of replaced shell. GHUNLPam 10138, 10459, 15314,
and 15091-15092.
Description. Preserved shell anterior wedge-shaped;
margins straight; apical angle of c. 40º; medial ridge
well defined; dorsal flank ornamented with c. 10
strong, slightly wavy radial ribs; ventral flank with
curved ribs meeting medial ridge at an angle of 25º;
shell thin, reaching up to 1.1 mm.
Remarks. The available specimens are very fragmentary
for any accurate comparison. However, shell
outline, radial ribs, median ridge, and curved ventral
ribs suggest that they are close to Pinna cretacea
(Schlotheim, 1813). This species of Pinna is very common
in Upper Cretaceous rocks from localities
spreading over most of the globe and seems to have
been cosmopolitan at that time (Seeling and
Bengston, 2003).
Age. Late Maastrichtian. Zone CC26 (late Chron
30N-Chron 29R).
Pinna sp. Figure 4. 2
Locality. Cerros Bayos (37º 40' S, 67º 30' W), province
of La Pampa.
Stratigraphic range. All the specimens come from
the Roca Formation, that at this locality is Danian.
Material. Six internal moulds, fragmented and very weathered,
but exhibiting vestiges of the original shell. GHUNLPam 1950-
1951, 2913-2914 and 4917-4918.
Description. Shell triangular, elongate, with terminal
umbones; anterior and posterior margins of anterior
part of the shell meeting at umbones at an angle of c.
35º; straight indistinct ridge running from anterior
end towards posterior margin, rendering a flat rhomboidal
shell section; outer surface with at least seven
straight, low, and thin radial ribs separated by wide
and shallow intercostal spaces on the dorsal flank,
and numerous curved ribs on the ventral flank that
intercept the medial ridge.
Measurements. GHUNLPam 2913, height: 51 mm,
width: 28 mm.
Remarks. Because of the poor preservation of the
available specimens, the interior of the shells is not
visible. Thus, it remains unclear whether the nacreous
layer was or not divided by a groove. However,
the narrow shell outline, the laterally rather flat section,
low apical angle, and very shallowly concave
ventral margin suggest that - while different from P.
aff. P. cretacea mentioned previously - these specimens
fit in Pinna. Also because of the fragmentary
nature of the available material, naming a new
species appears premature. Better specimens,
though, may confirm that it is in fact a distinct
Danian species from northern Patagonia.
The specimens are similar to Pinna anderssoni
Wilckens, 1910 (SMNH 1610-1625). However, the following
differences were noticed: the ventral flank of
the shells of P. anderssoni has five straight radial ribs
parallel to the medial ridge that are not seen in Pinna
sp.; the curved ribs near the ventral margin in specimens
of P. anderssoni do not intercept the medial
ridge as they do in Pinna sp.; and the anterior angle
is slightly smaller in P. anderssoni.
Pinna sp. is also similar to P. morenoi Wilckens,
1907 (p. 117-118, pl. 5, fig. 2a-b). These two species
can be separated by the smaller size of Pinna sp.,
its relatively more inflated shell, its finer ribs and
greater development of the medial keel.
Contrarily, P. sobrali Zinsmeister, 1984 (p. 1510, fig.
5G-5H; Stilwell and Zinsmeister, 1992, p. 59-60. pl.
3, fig. j-k), has a less clearly rhomboidal section
and fewer radial ribs, besides being an apparently
larger shell.
Differences are greater with P. freneixae Zinsmeister
and Macellari, 1988, from the Upper Cretaceous
beds of the López de Bertodano Formation
(units 5 through 9), in Seymour Island, Antarctica.
The Antarctic species has shells with a narrower apical
angle, stronger radial ribs, and a straight ventral
margin.
Comparison with other species referred to Pinna
is rather fruitless until the taxonomy of the Upper
Cretaceous and Paleogene members of this group
from the Southern Hemisphere is more clearly understood.
A revision of all the nominal fossil species
of Pinna, together with enhanced stratigraphic data
about them, is sure to reveal many cases of synonymy
and should certainly aid in the understanding
of the paleobiogeographic relationships within
this group of bivalves.
Age. Danian. Zone NP1 (Chron 29R-Chron 29N).
Genus Plesiopinna Amano, 1956
Type species. Plesiopinna atriniformis Amano, 1956; original designation.
Remarks. The genus Plesiopinna was introduced by
Amano (1956, p. 71, pl. 1, fig.1-5) for material collected
in the Albian-Cenomanian Gosyonoura Group exposed
at Shishijima, Japan. This peculiar shell is subtrigonal
in outline and two are the characters that
separate it from other members of this family. One of
these is the strongly curved medial ridge, that runs
from the umbones to the posteroventral position, describing
a well defined curve that renders the anteroventral
part of the concave shell. The other typical
feature of this genus is the very wide and flat saillike
posterior area. No traces of the interior are
known. Therefore, its affinities with other pinnids remain
uncertain. It has been classed as a subgenus of
Pinna, but the general shape reminds more of some
species of Atrina Gray, 1847. Until the interior is studied,
this can not be resolved. Besides the type species,
other material included in this genus is Plesiopinna sp. described by Cobban (1977, p. 14, pl. 1, fig. 1, 3)from the Oak Canyon Member of the Cenomanian
Dakota Sandstone of New Mexico. This specimen also
clearly shows the two shell features described for
the type species.
Plesiopinna pampeana n. sp.
Figure 4.3-6
Diagnosis. Shell trigonal to ham-shaped; tearshaped
section; anterior ventral margin concave; dorsal
margin straight; postumbonal keel very well defined
and clearly differentiated from the rest of the
shell surface, which is rather flat; ridge curved, describing
an arch following ventral margin from umbonal
area to posterior ventral sector of shell margin;
shell ornamentation of 12 strong and flat radial ribs
on dorsal flank, separated by widening intercostal
spaces, which at posterior margin reach up to three
times the width of ribs.
Locality. Barda Baya (36º 54' S, 67º 55'W), province of
La Pampa.
Stratigraphic range. The material comes from the
late Maastrichtian Jagüel Formation.
Material. Three moderately well preserved specimens, with replaced
shells. GHUNLPam 810 - 811 and 4825.
Holotype. Specimen GHUNLPam 810; the other specimens are
paratypes.
Type locality. Barda Baya (36º 54' S, 67º 55'W),
province of La Pampa.
Description. Trigonal ham-shaped shell; available
specimens broken, but seemingly large and very inflated
(preserved part of largest specimen, reaching
140 mm long and 82 mm wide and a width/height ratio
of 1.3 measured at 55 mm from the presumed anterior
end); umbones terminal, but broken in the three
known specimens; dorsal margin straight; ventral
margin sigmoid, concave near the anterior end; posterior
margin not preserved, but growth lines showing
it was gently rounded; maximum shell convexity
marked by a rounded and curved keel that runs from
the umbones to the posterior ventral margin, dividing
the shell surface into two distinct flanks; dorsal flank
flat, with twelve relatively wide, radial ribs separated
by wide intercostal spaces; both widen towards the
margin, intercostals more than ribs; rib section very
slightly convex; straight commarginal growth lines
on dorsal flank visible in intercostal spaces; commarginal
ribs intercepting median ridge at an angle of c.
30º to 35º. Shell thick (2.7 mm at most inflated part of
postumbonal ridge), decreasing towards dorsal margin,
where it is very thin.
Measurements. Holotype GHUNLPam 810, length:
113 mm; width: 82 mm.
Remarks. These remarkable shells clearly belong in
the Pinnidae, and fit best in Plesiopinna. Generic assignment
is supported by the most striking feature of
these shells, the curved median ridge that coupled
with the convex ventral flank and much flatter dorsal
flank produce the distinct tear shaped section. Although
the placement in this genus is relatively easy
on account of these characters, further studies including
a phylogenetic analysis among the Pinnidae
(extant and fossil) may prove otherwise. None of the
specimens shows the interior nacreous shell layer,
rendering it difficult to establish the presence or absence
of any bilobation or the shape and position of
the adductor muscle scar.
The species from La Pampa can be distinguished
from the type species by its larger shell, its flatter
posterior area and its ornamentation of radial ribs
covering the entire posterior flank. However, the
holotype is an internal mould with all but a few fragments
of the shell missing. The radial sculpture,
while visible, is not well preserved enough to compare
with our material. The postumbonal ridge is not
quite as strong as in Plesiopinna pampeana n. sp. nor is
the posterior area as flat as in our material.
Plesiopinna sp. from the Oak Canyon Member of
the Dakota Sandstone in New Mexico (Cobban, 1977,
p. 14, pl. 1, fig. 1, 3) resembles our species more closely
in shell outline and in the large and flat posterior
area and strongly curved postumbonal ridge.
However, the ornamentation is different in the two
species and allows easy separation. While the dorsal
part of the posterior flank of Plesiopinna sp. only
shows fine commarginal growth lines, in P. pampeana
this area is covered by radial ribs. In the North
American species, the radial ribs are restricted to the
ventral part of the dorsal flank and they are more numerous
and densely packed. The commarginal anteroventral
ribs are weaker and also far more restricted
than in P. pampeana.
South American species of Pinnids superficially
similar to Plesiopinna pampeana can be readily differentiated
as none of them show the straight dorsal margin,
flat posterior area and strongly curved and very inflated
umbonal ridge. Such is the case of Pinna tumida Philippi,
1899 (p. 50-51, pl. 25, fig. 1) from the Cretaceous of
Chile, Atrina reginamaris (Maury, 1930, p. 190, pl.7, fig.
3) from the Creataceous of northeastern Brazil, and A.
rioturbiensis Griffin, 1991 (p. 128-129, fig. 5.1-5.3), from
the Eocene of southwestern Patagonia.
Another species that resembles Plesiopinna pampeana
is Atrina triangularis Zinsmeister (1983, p. 1286,
fig. 1H) from the Paleocene of the Simi Hills in California.
It also has a broad triangular shape, but the
ridge is not as strongly marked and only very slightly
curved, while the ribs are narrower. The interior is
unknown too.
The striking shell-form of this pinnid may be related
to a life habit slightly different than the one
generally observed in modern Pinnids, i.e., vertically
buried in soft bottoms and byssally attached to
particles of sediment (Yonge, 1953; Stanley, 1970,
1972). The large relatively flat wing-like posterior
area and the strongly inflated and curved postumbonal
ridge suggest that - while still byssaly attached
as supported by the byssal gape - it could
have lived in a not quite as vertical position (i.e.,
with the ligament almost perpendicular to the substrate
surface), with the straight hinge line inclined
at an angle to the bottom and thus with the anteroventral
area within the sediment and the posterodorsal
expanded area above it. This mode of
life is suggested for other groups of bivalves with
analogous shell outlines such as Bakevellids and
Pteriids (Aberhan and Muster, 1997). It is also the
case of Bakevellia pintadae Damborenea, 1987 (p.
126-129, p. l 1, fig. 1-5; text fig. 4), from the Lower
Jurassic of the Neuquén Basin, and Isognomon lusitanicum
(Sharpe, 1849), I. praemytiloides Arkell, 1933
and I. volaticum Zakharov, 1966, from Jurassic localities
in the Northern Hemisphere (Fürsich, 1980).
Likewise, Bakevellids from the Lower Cretaceous
of the Neuquén Basin probably had a similar habit
(Lazo, 2003).
Age. Late Maastrichtian. Zone CC26 (late Chron 30
N-Chron 29R).
Etymology. Pampeana for La Pampa from were the
type specimen was collected.
Genus Atrina Gray, 1842
Type species. Pinna nigra Dillwyn, 1817; subsequent monotypy,
Gray, 1847.
Remarks. The distinguishing character that separates
Pinna from Atrina, i.e., the bilobation of the inner
prismatic layer, is not always preserved in fossil pinnids.
However, as discussed by other authors (Rosewater,
1961; Zinsmeister, 1984; Griffin, 1991), the
ham-shape of the shell and the rhomboidal shell section
are generally better developed in members of
Atrina, therefore allowing generic placement of fossil
material lacking other key characters.
Atrina ? sp. Figure 4.7
Locality. Liu Malal (36º 45' S, 68º 43'W), province of
Mendoza.
Stratigraphic range. Roca Formation, that at this locality
is Danian.
Material. One specimen, an internal mould. GHUNLPam 17308.
Remarks. The only available specimen probably
referable to Atrina is a poorly preserved internal
mould that nevertheless shows the strong, straight
postumbonal ridge typical of most species of this
genus. It somehow resembles A. rioturbiensis Griffin,
1991 (p. 128-129, fig. 5.1-5.3) from the Eocene
Río Turbio Formation, in southwestern Patagonia,
but further comparisons will be possible only with
better specimens in which an adequate suite of characters
can be observed. Pinna tumida Philippi, 1899
(p. 50-51, pl. 25, fig. 1) from the Cretaceous of Chile,
and Atrina reginamaris (Maury, 1930, p. 190, pl. 7,
fig. 3) from the Creataceous of northeastern Brazil
also show an overall similarity in shell shape and
section.
Age. Danian. Zone NP1-Zone NP2 (late Chron 29RChron
29N).
Conclusions
Although known from the earliest years of the
nineteenth century, the fossil invertebrate faunas
from the Maastrichtian and Paleocene of the Roca
and Jagüel formations have remained poorly understood.
This is in part due to the fact that the preservation
is generally poor, and intensive collection is
necessary to assemble enough specimens (other than
the extremely common oysters with calcitic shells) in
which key characters are present that may allow
proper taxonomic placement. Some of the material
described herein - while imperfectly preserved - in
general shows enough characters to allow positive
identifications. The new species described are all related
to taxa already known from other parts of the
world in rocks of the same or similar age. Such is the
case of the mytilids, in which very similar forms occur
in Africa, Europe, India, and North America.
However, the exact relationship among these taxa
must remain as yet uncertain, at least until better and
more abundant material is collected from a wider
range of localities.
The presence of Septifer in Maastrichtian rocks of
western La Pampa and southern Mendoza suggests
that during this time there were hard substrates
available nearby, as the specimens studied are generally
bivalved and relatively unworn. The location of
this kind of substrate has yet to be identified.
Contrarily, the rest of the fauna described consists of
soft bottom dwellers, a life mode consistent with the
lithology of the bearing rocks. Pinna cretacea is a cosmopolitan
species and the taxon found in the Roca
Formation is very close to it. On the other hand,
Plesiopinna seems to be a more restricted genus and
the specimens described are the only ones known so
far from South America.
The relationship between the distribution of this
group of bivalves and water temperature is still unclear,
although it appears possible that the presence
of Pinna sp. aff. P. cretacea in the area is reflecting the
cosmopolitan distribution of this taxon in accordance
with a global increase (at 65.5 Ma) in surface and intermediate
water temperatures of approximately 3-4
ºC (Barrera and Savin, 1999).
Of interest is the probable presence of Mytilaster,
a genus that nowadays inhabits brackish environments
in the Northern Hemisphere. Its occurrence in
beds of the Roca Formation needs to be properly assessed
in view of the normal marine character of the
rest of the bivalve taxa accompanying it.
A survey of the stratigraphic distribution of the
material described shows that the only Maastrichtian
species that survived into the Danian was Gregariella
amara suggesting that the environmental changes
taking place at this crucial time were important
enough to ensure a significant impact within the
Mytiloida.
References
1. Aberhan, M. and Muster, H. 1997. Paleobiology of Early Jurassic
Bakevellid bivalves from western Canada. Palaeontology 40:
799-815.
2. Amano, M. 1956. Some Cretaceous fossils from south-western
Japan (Part 1). Kumamoto Journal of Science, Series B, Section 1,
Geology 2: 63-93.
3. Arkell, W.J. 1933. The Jurassic System in Great Britain. Clarendon
Press, Oxford, 681 pp.
4. Barrera, E. and Savin, S.M. 1999. Evolution of late Campanian-
Maastrichtian marine climates and oceans. In: E. Barrera and
C.C. Johnson (eds.), Evolution of the Cretaceous Ocean-
Climate System, Special Paper, Geological Society of America 332:
245-282.
5. Bertels, A. 1968. Micropaleontología y estratigrafía del límite
Cretácico-Terciario en Huantrai-Có (provincia de Neuquén).
Parte 1: Ostrácoda. Ameghiniana 5: 279-298.
6. Bertels, A. 1969. Estratigrafía del límite Cretácico-Terciario en
Patagonia Septentrional. Revista de la Asociación Geológica
Argentina 24: 41-54.
7. Bertels, A. 1970. Los foraminíferos planctónicos de la cuenca
Cretácico-Terciaria en Patagonia septentrional (Argentina),
con consideraciones sobre la estratigrafía de Fortín General
Roca (Provincia de Río Negro). Ameghiniana 7: 1-56.
8. Beu, A.G. 2004. Marine Mollusca of oxygen isotope stages of the
last 2 million years in New Zealand. Part 1: Revised generic
positions and recognition of warm-water and cool-water migrants.
Journal of the Royal Society of New Zealand 34: 111-265.
9. Beu, A.G. and Maxwell, P.A. 1990. Cenozoic Mollusca of New
Zealand. Paleontological Bulletin New Zealand Geological Survey
58: 1-518.
10. Bodenbender, G. 1892. Sobre el terreno Jurásico y Cretácico de los
Andes Argentinos entre el Río Diamante y el Río Limay.
Boletín de la Academia Nacional de Ciencias 13: 5-44.
11. Böhm, J. 1903. Uber Ostreen von General Roca am Río Negro.
Zeitschrift der Deutschen Geologisches Gesellschaft 55: 71-72.
12. Brzozowska, J. 2004. Przebieg sedymentacji osadów "saramatu
detrytycznego" na obszarze pomiedzy Stawianami a
Sedziejowicami. Przeglad Geologiczny 52: 55-60.
13. Burckhardt, C. 1900. Profils géologiques transversaux de la Cordillère
Argentino-Chilienne. Stratigraphie et Tectonique. Anales del
Museo de La Plata, Sección Geológica y Mineralógica 2: 1-136.
14. Burckhardt, C. 1901. Le gisement supracrétacique de Roca (Río
Negro). Revista del Museo de La Plata 10: 1-16.
15. Butler, A.J., Vicente, N., and Gaulejac, B. de 1993. Ecology of the
pterioid bivalves Pinna bicolor Gmelin and Pinna nobilis L.
Marine Life 3: 37-47.
16. Camacho, H.H. 1970. Nuevos Fósiles del Cretácico Superior
(Maestrichtiano) del Departamento Malargüe, Provincia de
Mendoza, Argentina. Ameghiniana 7: 329-334.
17. Camacho, H.H. 1992. Algunas consideraciones acerca de la transgresión
marina paleocena en la Argentina. Academia Nacional
de Ciencias, Miscelanea 85: 1-41.
18. Casadío, S. 1998. Las ostras del límite Cretácico-Paleógeno de la
Cuenca Neuquina (Argentina). Su importancia bioestratigráfica
y paleobiogeográfica. Ameghiniana 35: 449-472.
19. Casadío, S., Griffin, M. and Parras, A. 2005. Camptonectes and
Plicatula (Bivalvia: Pteriomorphia) from the Upper
Maastrichtian of Northern Patagonia: Paleobiogeographic implications.
Cretaceous Research 26: 507-524.
20. Casadío, S., Parras, A., Concheyro, A. and Pires, M. 1998. El diacronismo
de la sedimentación carbonática en la Cuenca
Neuquina durante la transición Cretácico-Paleógena. 7º
Congreso Argentino de Paleontología y Bioestratigrafía (Bahía
Blanca), Resúmenes: 126.
21. Children, J.G. 1822-1824. Lamarck's genera of shells translated
from the French, with plates from original drawings by Miss
Anna Children. Quarterly Journal of Science 14 (1822): 64-86; 14
(1823): 298-322; 15 (1823): 23-52; 15 (1823): 216-258; 16 (1823):
49-79; 16 (1824): 241-264.
22. Clark, B.L. 1918. The San Lorenzo Series of middle California.
University of California Publication, Bulletin Department of
Geology 11: 45-234.
23. Cobban, W.A. 1977. Characteristic Marine Molluscan Fossils
From the Dakota Sandstone and Intertongued Mancos Shale,
West-Central New Mexico. Professional Paper, U. S. Geological
Survey 1009: 1-30.
24. Concheyro, A. and Villa, G. 1996. Maestrichtian-Danian (K/P) calcareous
nanofossils in the Liu Malal section. Northern
Patagonia, Argentina. Palaeopelagos 6: 281-297.
25. Cox, L.R. and Hertlein, L.G. 1969. Superfamily Pinnacea. In: R.C.
Moore (ed.), Treatise on Invertebrate Paleontology, Part N,
Mollusca, The Geological Society of America and the
University of Kansas, 6: N281-N285.
26. Damborenea, S.E. 1987. Early Jurassic Bivalvia of Argentina. Part
2: Superfamilies Pteriacea, Buchiacea and part of Pectinacea.
Palaeontographica Abteilung A, Band 199: 113-216.
27. Dartevelle, E. and Freneix, S. 1957. Mollusques fossiles du Crétacé
de la côte occidentale d'Afrique, du Cameroun à l'Angola.
Parte 2: Lamellibranches. Annales du Musée Royal du Congo
Belge, Série 8, 20: 1-271.
28. Del Valle, R. and Medina, F.A. 1980. Nuevos invertebrados fósiles
de Cabo Lamb (Isla Vega) y Cabo Morro (Isla James Ross).
Contribuciones del Instituto Antártico Argentino 228: 51-57.
29. Dillwyn, L.W. 1817. A descriptive Catalogue of Recent Shells. J. and
A. Arch, London, 2 v., 1092 pp.
30. Feldmann, R.M., Casadío, S., Chirino-Gálvez, L. and Aguirre-
Urreta, M.B. 1995. Fossil Decapod Crustaceans from the Jagüel
and Roca Formations (Maastrichtian-Danian) of the Neuquén
Basin, Argentina. Journal of Paleontology, Memoir 43: 1-22.
31. Feruglio, E. 1935. Relaciones estratigráficas y faunísticas entre los
estratos cretácicos y terciarios en la región austral del Lago
Argentino y en la del Golfo San Jorge (Patagonia). Boletín de
Informaciones Petroleras 128: 69-93; 130: 65-100.
32. Feruglio, E. 1936. Palaeontographia Patagonica. Memorie Instituto
Geologico della Universita di Padova 11: 1-384.
33. Frassinetti, D. and Covacevich, V. 1999. Invertebrados fósiles
marinos de la Formación Guadal (Oligoceno superior-
Mioceno inferior) en Pampa Castillo, Región de Aisén, Chile. Boletín del Servicio Nacional de Geología y Minería de Chile 51: 1-
96.
34. Fritzsche, H. 1919. Eine Fauna aus schichten der Kreide-
Tertiärgrenze in der Argentinischen Cordillere des südlichen
Mendoza. Centralblatt für Mineralogie, Geologie, und
Paläontologie 1919: 359-369.
35. Fürsich, F. 1980. Preserved life position of some Jurassic bivalves.
Paläontologisches Zeitschrift 54: 289-300.
36. Gmelin, J.F. 1791. Caroli a Linné Systema Naturae per Regna Tria
Naturae, Secundum Classes, Ordines, Genera, Species, cum characteribus,
differentiis, synonymis, locis. Editio decima tertia, aucta,
reformata, cura J. F. Gmelin. Georg. Emanuel., Leipzig, pp.
3021-3910.
37. Gray, J.E. 1842. Synopsis of Contents of the British Museum. London,
308 pp.
38. Gray, J.E. 1847. A list of the genera of Recent Mollusca, their synonyms
and types. Proceedings Zoological Society of London 15:
129-219.
39. Griffin, M. 1991. Eocene Bivalves from the Rio Turbio Formation,
Southwestern Patagonia (Argentina). Journal of Paleontology
65: 119-146.
40. Griffin, M., Casadío, S. and Parras, A. 2005. Maastrichtian and
Danian Species of Turkostreini (Ostreidae, Crassostreinae)
from the Neuquén Basin, Argentina. Ameghiniana 42: 257-276.
41. Ihering, H. von. 1903. Les mollusques des terrains crétaciques superieurs
de l'Argentine orientale. Anales del Museo Nacional de
Buenos Aires, serie 3, 2: 193-229.
42. Ihering, H. von. 1907. Les mollusques fossiles du Tertiaire et du
Crétacé superieur de l'Argentine. Anales del Museo Nacional de
Buenos Aires, serie 3, 7: 1-611.
43. Ihering, H. von. 1914. Catalogo de Molluscos cretáceos e terciarios
da collecçao do auctor. Notas do Museu Paulista 1: 1-113.
44. Lamarck, J.B. de. 1819. Histoire naturelle des animaux sans vertèbres.
Lamellibranches. (chez l'auteur, au Jardin du Roi), Paris 6: 1-
233.
45. Lazo, D.G. 2003. Taxonomy, facies relationships and palaeobiology
of bakevelliid bivalves from the Lower Cretaceous of westcentral
Argentina. Cretaceous Research 24: 765-788.
46. Linné, C. 1758. Systema Naturae per regna tria naturae. Vol. 1,
Regnum animale, Editio decima, Edit. Laurentii Salvii,
Estocolmo, 823 pp.
47. Malumián, N. and Caramés, A. 1995. El Daniano marino de
Patagonia (Argentina): paleobiogeografía de los foraminíferos
bentónicos. In: C. Náñez (ed.), Paleogeno de América del Sur,
Publicación Especial de la Asociación Paleontológica Argentina 3:
83-105.
48. Matheron, P. 1843. Catalogue méthodique et descriptif des corps
organisés fossiles du département des Bouches du Rhône, et
lieux circonvoisins: précédé d'un mémoire sur les terrains
supérieurs au grès bigarré du S. E. de la France. Répertoire des
Travaux de la Société Statistique de Marseille 6: 1-269.
49. Maury, H. 1930. O Cretáceo de Parahyba do Norte. Monografía
Serviço Geológico e Mineralógico do Brasil 8: 1-305.
50. Monterosato, T.A. 1883. Conchiglie littorali mediterranee. Naturalista Siciliano 3: 87-91.
51. Moore, E.J. 1983. Tertiary Marine Pelecypods of California and
Baja California: Nuculidae through Malleidae. Professional
Paper, United States Geological Survey 1228-A: 1-99.
52. Mörch, O.A.L. 1853. Catalogus conchyliorum quae reliquit D.
Alphonso d'Aguirra et Gadea, Comes de Yoldi, regis Daniae cubiculariorum
princeps, ordinis Danebrogici in prima classe et ordinis
tertii eques. Fasc. Secundus, Acephala. Ludovici Kleini,
Copenhagen, 74 pp.
53. Ortmann, A.E. 1902. Tertiary Invertebrates. In: W.B. Scott (ed.),
Reports of the Princeton University Expedition to Patagonia
1896-1899, volume 4, Paleontology I, Part 2: 45-332, J. Pierpoint Morgan Publishing Foundation, Princeton.
54. Packard, E.L. and Jones, D.L. 1965. Cretaceous pelecypods of the
genus Pinna from the west coast of North America. Journal of
Paleontology 39: 910-915.
55. Parras, A., Casadío, S. and Pires, M. 1998. Secuencias depositacionales
del Grupo Malargüe (límite Cretácico-Paleógeno), sur de la
provincia de Mendoza, Argentina. In: S. Casadío (ed.), Paleógeno
de América del Sur y de la Península Antártica, Publicación
Especial de la Asociación Paleontológica Argentina 5: 181-19.
56. Péron, A. 1890. Exploration scientifique de la Tunisie. Description de
mollusques fossiles des terrains crétacés de la région sud des Hauts-
Plateaux de la Tunisia, recueillis en 1885 et 1886 par M. Ph.
Thomas. 2º Partie. Pélécypodes. Imprimerie Nationale, París, pp.
105-327.
57. Philippi, R.A. 1899. Los fósiles secundarios de Chile. Brockhaus,
Leipzig - Santiago de Chile, 104 pp.
58. Poli, G. 1795. Testacea utriusque Siciliae eorumque historia et anatome
tabulis aeneis illustrata. Regio Typographeio, Parma, 264 pp.
59. Popenoe, W.P. 1937. Upper Cretaceous Mollusca from southern
California. Journal of Paleontology 11: 379-402.
60. Recluz, C.A. 1848. Description d'un Nouveau Genre de Coquilles
Bivalves nommé Septifère (Septifer). Revue Zoologie 1848: 275-
279.
61. Richardson, C.A., Kennedy, H., Duarte, C.M., Kennedy, D.P., and
Proud, S.V. 1999. Age and growth of the fan mussel Pinna nobilis
from south-east Spanish Mediterranean seagrass
(Posidonia oceanica) meadows. Marine Biology 133: 205-212.
62. Risso, J. A. 1826. Histoire naturelle des principales productions de
l'Europe méridionale et particulièrement de celles des environs de
Nice et des Alpes Maritimes, volume 4, Aperçu sur l'histoire naturelle
des mollusques qui vivent sur les bords de la Mediterranée
boréale et des coquilles terrestres, fluviatiles, et marines, subfossiles,
fossiles et pétrifiées, qui gisent dans les diverses formations des Alpes
Maritimes. F. G. Levrault, Paris, 439 pp.
63. Röding, P.F. 1798. Museum Boltenianum sive Catalogus cimeliorum e
tribus regnis naturae quae olim collegerat Joa. Fried. Bolten,
M.D.p.d. Pars Secunda, Conchylia sive Testacea Univalvia,
Bivalvia et Multivalvia. Johan Christi Trapii, Hamburg, 119 pp.
64. Rosewater, J. 1961. The Family Pinnidae in the Indo-Pacific. Indo-
Pacific Mollusca 1: 175-226.
65. Say, T. 1825. On a New Species of Modiola. Journal of the Academy
of Natural Sciences of Philadelphia 4: 368-370.
66. Schlotheim, E. F. von. 1813. Beiträge zur Naturgeschichte der
Versteinerungen in geognostischer Hinsicht: Taschenbuch für
gesammte Mineralogie, C. C. Leonhard, Frankfurt, 7: 3-135.
67. Seeling, J. and Bengston, P. 2003. The bivalve Pinna cretacea
(Schlotheim, 1813) from the Cretaceous of Brazil. Acta
Paleontologica Polonica 48: 475-480.
68. Selin, N. and Latypov, Y. 2006. Distribution pattern, population
structure, and growth of Septifer bilocularis (Bivalvia:
Mytilidae) on reefs of Southern Vietnam. Russian Journal of
Marine Biology 32: 88-95.
69. Sharpe, D. 1849. On the Secondary District of Portugal which lies
on the North of the Tagus. Quarterly Journal of the Geological
Society of London 6: 135-201.
70. Soot-Ryen, T. 1955. A report on the Family Mytilidae
(Pelecypoda). Publications University Southern California, Alan
Hancock Pacific Expeditions 20: 1-170.
71. Sowerby, J. de C. 1812. Mineral Conchology of Great Britain; or
Coloured Figures and Descriptions of those Remains of Testaceous
Animals of Shells, which have been preserved at Various Times and
Depths in the Earth. Meredith, London, v. 1, 244 pp.
72. Stanley, S.M. 1970. Relation of shell form to life habits of the
Bivalvia (Mollusca). Memoir Geological Society of America 125: 1-
296.
73. Stanley, S.M. 1972. Functional morphology and evolution of
byssally attached bivalve mollusks. Journal of Paleontology 46:
165-212.
74. Stilwell, J.D. and Zinsmeister, W.J. 1992. Molluscan Systematics
and bioestratigraphy. Lower Tertiary La Meseta Formation,
Seymour Island, Antarctic Peninsula. American Geophysical
Union, Anctarctic Research Series 55: 1-192.
75. Stoliczka, F. 1870-1871. Cretaceous fauna of southern India, Vol. 3,
The Pelecypoda, with a review of all known genera of this
class, fossil and Recent. Memoirs of the Geological Survey of
India, Palaeontologia Indica 6: 1-537.
76. Swainson, W. 1840. A treatise on malacology of shell and shell-fish.
The Cabinet Cyclopedia, Natural History. Longman, London, 420
pp.
77. Tessier, F. 1952. Contribution à la Stratigraphie et à la Paléontologie de
la Partie Ouest du Sénégal (Crétacé et Tertiarie). Troisième partie.
Paléontologie (Thèse Aix-Marseille 1950). Faculté des Sciences de
Marseille, Dakar, pp. 281-571.
78. Turner, R. and Rosewater, J. 1958. The Family Pinnidae in the
Western Atlantic. Johnsonia 3: 285-326.
79. Weaver, C.E. 1927. The Roca Formation in Argentina. American
Journal of Science, Serie 5, 13: 417-434.
80. Weaver, C.E. 1931. Paleontology of the Jurassic and Cretaceous of
West Central Argentina. Memoir University of Washington 1: 1-
469.
81. Wilckens, O. 1907. Die Lamellibranchiaten, Gastropoden u.s.w.
der oberen Kreide Südpatagoniens. Berichte der naturforschenden
Gesellschaft zu Freiburg 15: 97-166.
82. Wilckens, O. 1910. Die Anneliden, Bivalven, und Gastropoden
der antarktischen Kreideformation. Wissenschaftlich
Ergebenisse der Schwedischen Südpolar-Expedition, 1901-1903,
unter der Leitung von Dr. Otto Nordenskjöld 3: 1-132.
83. Yonge, C.M. 1953. Form and habit in Pinna carnea Gmelin.
Philosophical Transactions, Royal Society of London, Series B 237:
335-374.
84. Zakharov, V.A. 1966. Late Jurassic and early Cretaceous Bivalvia
(Anisomyaria) in northern Siberia and the conditions of their existence.
Akademiya Nauk SSSR, Moscow, 184 pp.
85. Zinsmeister, W.J. 1983. New late Paleocene mollusks from the
Simi Hills, Ventura County, California. Journal of Paleontology
57: 1282-1303.
86. Zinsmeister, W.J. 1984. Late Eocene bivalves (Mollusca) from the
La Meseta Formation, collected during the 1974-1975 joint
Argentine-American expedition to Seymour Island, Antarctic
Peninsula. Journal of Paleontology 58: 1497-1527.
87. Zinsmeister, W.J. and Macellari, C.E. 1988. Bivalvia (Mollusca)
from Seymour Island, Antarctica Peninsula. In: R.M.
Feldmann and M.O. Woodburne (eds.), Geology and
Paleontology of Seymour Island, Antarctic Peninsula, Memoir
of Geological Survey of America 169: 253-283.
Recibido: 22 de noviembre de 2006.
Aceptado: 27 de noviembre de 2007.