
Figura 1. 1, ubicación general de los afloramientos / location map; 2, perfil estratigráfico de la localidad PBN / stratigraphical section at PBN; 3, perfil estratigráfico de la localidad PBS / stratigraphical section at PBS.
Estudio palinológico de la Formación Punta del Barco, Cretácico Inferior (Aptiano) de la provincia de Santa Cruz, Argentina. Descripciones sistemáticas I: esporas monoletes y triletes lisas y cinguladas
Magdalena Llorens1
1Museo Paleontológico ¨Egidio Feruglio¨. Av. Fontana 140, 9100 Trelew, Chubut, Argentina. magdalena.llorens@gmail.com
Resumen. Se describen sistemáticamente 20 especies de esporas dispersas trilete lisas, cinguladas y monoletes. El material procede de la Formación Punta del Barco, Cretácico Inferior de la provincia de Santa Cruz. Estas formas pertenecen a los morfogéneros Biretisporites, Deltoidospora, Todisporites, Cyathidites, Densoisporites, Foraminisporis, Contignisporites, Laevigatosporites y Reticulosporis. Este último se reporta por primera vez para Argentina, constituyendo además uno de los registros más antiguos del género. Algunos de estos taxones representan a grupos vegetales que no habían sido reportados anteriormente para esta tafoflora, ampliando así su conocimiento previo.
Abstract. Palynological Study Of The Punta Del Barco Formation, Lower Cretaceous Of Santa Cruz Province, Argentina. Systematic Descriptions I: Monolete And Smooth And Cingulate Trilete Spores. Smooth and cingulate trilete and monolete disperse spores from the Punta del Barco Formation, Lower Cretaceous of Santa Cruz Province are being described in this paper. The trilete spores belong to the genera Biretisporites, Deltoidospora, Todisporites, Cyathidites, Densoisporites, Foraminisporis, Contignisporites, Laevigatosporites and Reticulosporis. The last genus is mentioned for the first time in Argentina, and is one of the oldest known records. Some of these taxa represent vegetal groups no reported for this taphoflora, enlarging therefore its previous knowledge.
Palabras clave. Palinología; Sistemática; Cretácico Inferior; Provincia de Santa Cruz; Argentina.
Key words. Palynology; Systematic; Lower Cretaceous; Santa Cruz Province; Argentina.
Introducción
El Grupo Baqueró (ex Formación Baqueró, Archangelsky 1967) está compuesto -de base a techopor las formaciones Anfiteatro de Ticó, Bajo Tigre y Punta del Barco, y se encuentra ampliamente distribuido en el centro de la Provincia de Santa Cruz en el ámbito del Cratón del Deseado (Cladera et al., 2002). Su unidad superior -la Formación Punta del Barco- motivo de este estudio, aflora casi exclusivamente en el área de la Meseta Baqueró (figura 1.1), y está compuesta por depósitos volcaniclásticos, en particular epiclásticos y piroclásticos. Los primeros dominan la mitad inferior de la columna estratigráfica con conglomerados y areniscas en facies de canal, arcilitas y limolitas gris oscuras; mientras los segundos caracterizan la mitad superior, que incluye tobas primarias altamente friables y otras retrabajadas (figura 1.2, 1.3). Esta litología ha sido interpretada como un paleoambiente de llanura surcada por un sistema fluvial entrelazado (Cladera et al., 2002).
Figura 1. 1, ubicación general de los afloramientos / location map; 2, perfil estratigráfico de la localidad PBN / stratigraphical section at PBN; 3, perfil estratigráfico de la localidad PBS / stratigraphical section at PBS.
Por otra parte, el Grupo cuenta con dataciones
isotópicas realizadas con el método 40Ar/39Ar, sobre
dos muestras provenientes de niveles tobáceos. Uno
de ellos corresponde a la sección superior de la
Formación Anfiteatro de Ticó, con un resultado de
118,56 ± 1,40 Ma; en tanto la restante, a la base de la
mitad superior de la sección de Formación Punta del
Barco en la localidad homónima, y su estudio arrojó
una edad de 119,65 ± 0,45 Ma (Corbella, 2001). El
análisis de las muestras se realizó en University of
Oregon (USA), y permitió ubicar al Grupo Baqueró
en el Aptiano temprano.
El estudio paleontológico de este Grupo ha tenido
numerosos aportes, dados la abundancia y el excelente
grado de preservación que presentan los restos
vegetales tanto mega como microscópicos. En su
gran mayoría, esos estudios se refieren a la Formación
basal -Anfiteatro de Ticó- de la que se conocen
ampliamente su megaflora y contenido polínico
(véase compendio en Archangelsky, 2003). Mientras
para la Formación Punta del Barco habían sido reportados sólo algunos restos vegetales megascópicos,
en su gran mayoría improntas (Berry, 1924;
Feruglio, 1951; Herbst, 1960, 1962 y 1966 -ver lista
completa en Archangelsky, 2001-); entre ellos se destacan
varios helechos (Hausmannia, Gleichenites, Cladophlebis) y algunas gimnospermas (Araucaria, Pseudoctenis). Estas últimas han brindado también
restos de tejido cuticular en estado de charcolificación
o momificación (Del Fueyo y A. Archangesky,
2002; A. Archangelsky et al., 1995; Llorens, et al.,
2003).
Aunque la particular historia tafonómica de la vegetación aptiana (Cladera y Cúneo, 2002) ha permitido registrar muy bien la paleoflora del momento, esta formación presenta una litología poco promisoria para contener palinomorfos. El primer indicio que sugirió la posibilidad de la presencia de palinomorfos en la Formación Punta del Barco fue el hallazgo de excelentes cutículas de la cicadal Pseudoctenis ornata (A. Archangelsky et al., 1995). Es así que la unidad superior no brindó información de su microflora sino hasta hace pocos años, cuando se dio a conocer el primer registro palinológico (Llorens, 2000). Se realizó también un trabajo tafonómico basado en una singular asociación de esporas perisporadas (Llorens y Cladera, 2002) y recientemente, se reportó una asociación polínica compuesta mayormente por polen de angiospermas y organismos planctónicos (Llorens, 2003a). En ese mismo año, la autora presentó un estudio preliminar de los granos de polen afines a las plantas con flor hallados en esta asociación (Llorens, 2003b).
En esta publicación se aborda al estudio taxonómico de los palinomorfos registrados en la Formación Punta del Barco, comenzando con la descripción sistemática de las esporas dispersas pertenecientes a las turmas Monoletes y Triletes (infraturmas Laevigati y Cingulati).
Materiales y métodos
Se estudiaron las asociaciones polínicas provenientes de 6 niveles fértiles que forman una secuencia, siendo el nivel 1 el más antiguo y el nivel 6 el más moderno. Esta secuencia resultó de la integración de los datos obtenidos en los distintos niveles fértiles de las dos localidades estudiadas: Punta del Barco Norte (PBN) y Punta del Barco Sur (PBS), obtenida mediante los criterios de arreglos de facies y los contactos entre las diferentes formaciones, tanto en su relación de techo como de base. Otra herramienta de correlación utilizada fueron los diferentes niveles fosilíferos comunes a ambos perfiles. El punto de partida para esta integración fue el evidente contacto erosivo de la Formación Punta del Barco con las infrayacentes Bajo Tigre y Anfiteatro de Ticó, respectivamente. Una vez integradas sus bases, el siguiente punto de correlación es una toba de casi 1 metro de espesor, portadora de plantas fósiles de taxones presentes en las dos columnas en cuestión (Llorens, 2005). Se halló también una muy buena correlación de los 6 niveles pelíticos de este muestreo palinológico en ambos perfiles. Finalmente, se utilizó la transgresión marina de pelitas verdes portadoras de invertebrados fósiles de la Formación Monte León (Mioceno). Los niveles N1, N3 y N6 provienen de PBN, en cambio, los niveles N2 y N5, de PBS. El nivel N4 es el único representado en ambos perfiles, aunque la mayor parte de la información procede de la muestra tomada en PBS (figuras 1.2, 1.3).
El procesamiento de las muestras se llevó a cabo
en gran parte en el laboratorio de la División Paleobotánica
del Museo Argentino de Ciencias Naturales
"Bernardino Rivadavia" (MACN) y el resto, en el laboratorio
de ácidos del Museo Paleontológico Egidio
Feruglio (Mef), siguiendo el método clásico de HFHCl,
con especial cuidado en reducir los riesgos de
destrucción del material por medios físicos. En las
muestras que presentaron abundante materia orgánica
amorfa, se aplicó una breve oxidación adicional
con ácido nítrico. El residuo obtenido se filtró por
malla de 100 y 25 µm, y en las dos muestras en que se
detectó la presencia de granos de polen afines a las
angiospermas se realizó también un filtrado por malla
de 10 µm para facilitar la visión de los mismos. El
montaje de las diferentes fracciones del residuo se
llevó a cabo con glicerina-gelatina para ser observado
con microscopía óptica. Los microscopios ópticos
utilizados fueron el Leitz Dialux Nº 881392 y Leitz
Diaplan 512834/135200, ubicados en el Mef y en la
Div. Paleobotánica del MACN respectivamente. Las
fotomicrografías se tomaron con el microscopio Zeiss
Axioskop 2, propiedad de la Dra. A. Archangelsky, y
las coordenadas de los ejemplares aquí ilustrados corresponden
al mismo. Las ilustraciones de microscopio
electrónico de barrido se obtuvieron en el Instituto
Botánico Darwinion de Buenos Aires, con un
microscopio ZEISS 940A.
El repositorio se encuentra en la División
Paleobotánica del MACN "Bernardino Rivadavia"
bajo la sigla BA Pal Nº 5500 al 5504 y 5550 al 5577 y
en la colección del Museo Paleontológico Egidio
Feruglio, MPEF Palin 56 al 62.
Paleontología sistemática
Lista de especies de esporas descriptas para la Formación Punta del Barco
Biretisporites cf. B. potoniaei Delcourt y Sprumont 1955
Biretisporites sp. A
Biretisporites sp. B
Biretisporites sp. C
Contignisporites glebulentus (Dettmann) emend. Filatoff y Price 1988
Contignisporites burgerii Filatoff, Mc Kellar y Price en: Filatoff y
Price 1988
Contignisporites cooksoniae (Balme) Dettmann 1963
Cyathidites sp. cf. C. minor Couper 1953 Deltoidospora sp. A
Deltoidospora sp. B
Densoisporites velatus (Weyland y Krieger) emend. Krasnova ex. Dettmann 1963
Densoisporites corrugatus Archangelsky y Gamerro 1965
Foraminisporis dailyi (Cookson y Dettmann) Dettmann 1963
Foraminisporis sp.
Laevigatosporites ovatus Wilson y Webster 1946
Laevigatosporites sp.
Reticulosporis sp. 1
Reticulosporis sp. 2
Reticulosporis sp. 3
Todisporites sp.
Turma TRILETES Reinsch emend. Dettmann 1963
Subturma AZONOTRILETES Luber emend. Dettmann 1963
Infraturma LAEVIGATI (Bennie y Kidston) Potonié 1956Género Biretisporites (Delcourt y Sprumont) Delcourt, Dettman y Hughes 1963
Especie tipo. B. potoniaei, Delcourt y Sprumont 1955.
Biretisporites cf. B. potoniaei Delcourt y Sprumont 1955 Figura 2.1
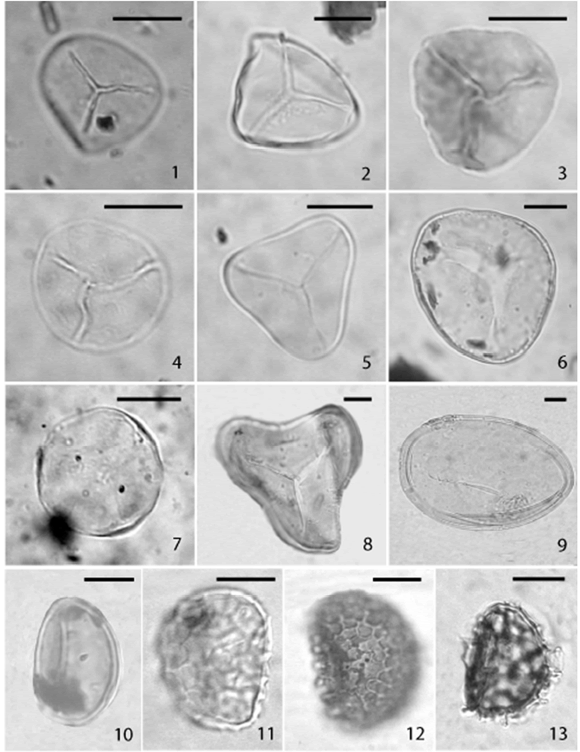
Figura 2. Esporas lisas y monoletes / smooth and monolete spores; 1, Biretisporites cf. B. potoniaei Delcourt y Sprumont (Pm BA Pal Nº
5570: 6.2/90.3); 2, Biretisporites sp. A (Pm BA Pal Nº 5552: 11.6/83.5); 3, Biretisporites sp. B (Pm BA Pal Nº 5552: 1.1/92.1); 4, Biretisporites sp. C (Pm BA Pal Nº 5552: 18.9/88.6); 5, Deltoidospora sp. A (Pm BA Pal Nº 5552: 3.9/112.4); 6, Deltoidospora sp. B (MPEF
Palin 61: 92.3/10.8); 7, Todisporites sp. (Pm BA Pal Nº 5566: 6.6/96); 8, Cyathidites sp. cf. C. minor Couper (Pm BA Pal Nº 5571: 4.6/95.9); 9, Laevigatosporites sp. (Pm BA Pal Nº 5559: 17.6/86); 10, Laevigatosporites ovatus Wilson y Webster (Pm BA Pal Nº 5570: 10.9/92.8); 11, Reticulosporis sp. 1 (Pm BA Pal Nº 5552: 10.9/92.8); 12, Reticulosporis sp. 2 (Pm BA Pal Nº 5567: 9.5/93,7); 13, Reticulosporis sp. 3 (Pm BA Pal Nº 5569: 21.9/92,3). Escala / scale bar: 10 µm.
Descripción. Esporas triletes de contorno ecuatorial
subtriangular con ángulos ampliamente redondeados
o bien subcircular, y cara distal globosa. La lesura
es recta acompañada por labios de un espesor variable,
y un largo de 4/5 del radio. Las ramas de la lesura
pueden verse con alguna sinuosidad, pero nunca
en las tres de un mismo ejemplar. Exina psilada, o
bien ligeramente granular en la cara distal, debido a
una corrosión incipiente.
Dimensiones. (13 ejemplares) Diámetro ecuatorial:
15,5(18,5)21,7 µm; espesor de la exina: 0,4-0,9 µm; ancho
del labio: 0,7-1,2 µm; alto del labio: 0,9-2 µm.
Discusión y comparaciones. Esta forma es comparable
a la especie tipo del género, pero no puede ser
asignada a ella definitivamente por poseer un tamaño
sensiblemente menor (50-60 µm en B. potoniaei); la
altura de los labios de la lesura también es más pequeña,
respondiendo al tamaño y la relación diámetro
ecuatorial/altura del labio. Morfológicamente es
muy similar a Biretisporites sp. de la Formación
Springhill de Patagonia Austral (Baldoni y
Archangelsky, 1983), pero en ese caso los ejemplares
tienen un tamaño sensiblemente mayor (70-79 µm).
Material estudiado. PBN N1: BA Pal 5552, 5569, 5570, 5574 y 5575.
Biretisporites sp. A Figura 2.2
Descripción. Esporas triletes lisas, de contorno ecuatorial
subtriangular con los ángulos ampliamente redondeados.
La cara distal es claramente convexa,
mientras que la proximal se presenta en forma piramidal,
con los rayos de la marca trilete alcanzando el
ecuador, acompañados por labios tenues, angostos y
rectos.
Dimensiones. (2 ejemplares) diámetro ecuatorial
27,5-28,6 µm; espesor de la exina 0,7-1 µm; labio: ancho
0,7 µm, y alto 0,8 µm.
Discusión y comparaciones. Se incluye esta forma en
el género Biretisporites por sus características generales,
pero no se realizó su asignación específica debido
a que sólo se encontraron dos ejemplares, cuyas
dimensiones difieren de las especies conocidas.
Material estudiado. PBN N1: BA Pal 5552.
Biretisporites sp. B Figura 2.3
Descripción. Esporas pequeñas biconvexas, de contorno
subtriangular a subcircular y de aspecto globoso. La
lesura es levemente sinuosa, se extiende hasta el ecuador
flanqueada por labios más o menos angostos y altos,
razón por la cual se observan ondulaciones debidas
a compresión. Exina totalmente lisa en la mayoría
de los casos, o bien con irregularidades por diferencia
de espesor, probablemente producto de corrosión.
Dimensiones. Diámetro ecuatorial (10 ejemplares) 11,8
(18) 19,8 µm; espesor de la exina (6 ejemplares) 0,4-0,9
µm; ancho del labio 0,8-0,9 µm; alto del labio 1,1-1,5 µm.
Discusión y comparaciones. Se incluye esta forma en
el género por coincidir en los caracteres dados en la
diagnosis enmendada por Delcourt et al. (1963).
Existen similitudes con Biretisporites cf. B. potoniaei en
Prámparo (1988), pero en ese caso las dimensiones
son sensiblemente mayores y la lesura recta, sin alcanzar
los ángulos. Por otro lado, B. potoniaei del
Albiano de Oklahoma (Wingate, 1980) tiene un gran
parecido y un tamaño también menor al de la especie
tipo (34-50 µm), pero no alcanza a ser tan reducido
como en este material.
Material estudiado. PBN N1: BA Pal 5566, 5552, 5577 y 5568.
Biretisporites sp. C Figura 2.4
Descripción. Esporas trilete biconvexas y globosas,
de contorno ecuatorial circular. Lesura recta a levemente
sinuosa, encerrada entre labios delgados y bajos,
llegando hasta el ecuador. La exina es levigada en
la mayoría de los casos, pero también puede aparecer
escabrada; es muy delicada, por lo que se encuentra
frecuentemente plegada.
Dimensiones. (20 ejemplares) diámetro ecuatorial
15,4 (20,5) 27,5 µm; espesor de la exina 0,4-1 µm; ancho
del labio 0,7-1 µm; alto del labio 0,9-1,5 µm.
Comentarios. La morfología general de estas esporas
es comparable con el género Todisporites, pero la presencia
de un labio como carácter continuo para el
conjunto, permite ubicarla dentro de Biretisporites. Si
bien el número de ejemplares de este tipo es suficiente
para proponer un nuevo taxón, se ha preferido
no realizar una asignación específica por encontrar
una marcada semejanza con el cuerpo central de una
espora perisporada muy abundante en este nivel
(PBN N1), en el que además a menudo se encuentra
la perina aislada, por lo que es muy probable que este
tipo morfológico corresponda a ejemplares incompletos.
Sin embargo, se considera conveniente describirlos
y ubicarlos en un morfogénero que los caracterice,
hasta tanto esa espora perisporada sea descripta
en una futura contribución.
Material estudiado. PBN N1: BA Pal 5551, 5552, 5566 y 5577.
Género Deltoidospora Miner 1935
Especie tipo. Deltoidospora hallii Miner 1935.
Afinidad botánica. El hallazgo de estas formas estaría asociado con el de algunos helechos mesozoicos afines a las familias Gleicheniaceae y Cyatheaceae (Balme, 1995; Jansonius y Hills, 1976: 748).
Deltoidospora sp. A Figura 2.5
Descripción. Esporas trilete subtriangular de lados
rectos y ángulos redondeados. Exina lisa; lesura simple
de ramas rectas a levemente sinuosas que se extienden
hasta el ecuador.
Dimensiones. (2 ejemplares) Diámetro ecuatorial
17,5-22 µm; espesor de la exina 0,5 µm.
Discusión y comparaciones. Esta forma tiene una gran
similitud con el género Leiotriletes, según la enmienda
realizada por Potonié y Kremp (1954) donde se dio como
carácter diagnóstico para ese género una configuración
piramidal de las facetas (referencia de Jansonius
y Hills, 1976: 1472). Sin embargo en los ejemplares aquí
estudiados, esa configuración es deltoide, razón por la
cual se asigna a Deltoidospora, sin poder darles un nombre
específico por la escasez de material.
Material estudiado. PBN N1: BA Pal 5552.
Deltoidospora sp. B Figura 2.6
Descripción. Esporas trilete de contorno subtriangular
a subcircular con lados convexos y ángulos ampliamente
redondeados. Exina gruesa, rígida y lisa
en su superficie externa, mientras que la superficie
interna se presenta rugosa. Lesura recta que se extiende
casi hasta el ecuador.
Dimensiones. (8 ejemplares) diámetro ecuatorial 29-
38 µm; espesor de la exina 1,5 µm.
Comparaciones. La principal diferencia de estas esporas
con la especie tipo, con Deltoidospora sp. A y con
D. minor citadas para el Grupo Baqueró (Archangelsky
y Villar de Seoane, 1994), radica en que todas
poseen lados rectos o levemente cóncavos que les
confieren una forma triangular, mientras que D. sp. B.
tiene un contorno subtriangular a subcircular.
Material estudiado. PBN N3: MPEF Palin 61.
Género Todisporites Couper 1958
Especie tipo. Todisporites major Couper (1958).
Todisporites sp. Figura 2.7
Descripción. Esporas trilete marcadamente globosas,
biconvexas y con contorno ecuatorial siempre circular.
Exina psilada, hialina y rígida. La lesura es poco
conspicua, de ramas rectas que llegan hasta el ecuador
y puede pasar al hemisferio distal, aparentando
estar bifurcada en el extremo.
Dimensiones. (4 ejemplares) diámetro ecuatorial 16-
36 µm; espesor de la exina 0,7-1 µm.
Discusión y comparaciones. Esta forma es diferente
a la especie tipo y a otras formas previamente descriptas
para este morfogénero en la bibliografía por
no presentar pliegues en la exina. Sin embargo, el escaso
material rescatado hace imposible proponer una
asignación específica.
Afinidad botánica. Familia Osmundaceae (Balme,
1995).
Material estudiado. PBN N1: BA Pal 5566 y 5552; PBS N4: MPEF Palin 56.
Género Cyathidites Couper 1953
Especie tipo. Cyathidites australis Couper 1953.
Cyathidites sp. cf. C. minor Couper 1953 Figura 2.8
Descripción. Espora trilete de contorno ecuatorial
triangular, de lados cóncavos y ángulos ampliamente
redondeados. La lesura es simple y bien delimitada,
con ramas rectas que llegan hasta 2/3 del radio.
Exina gruesa, lisa a escabrada.
Dimensiones. (1 ejemplar) diámetro ecuatorial 48 µm;
espesor de la exina 1,2 µm; ramas de la lesura 16 µm.
Discusión y comparaciones. Esta espora es muy semejante
a las descriptas por Archangelsky (1972) para
el Eoceno de Río Turbio en Santa Cruz como C. patagonicus (70-90µm), diferenciándose por el diámetro
ecuatorial que en el material aquí estudiado es considerablemente
menor. Por otro lado, es muy semejante
también a la especie tipo, con la que coincide en dimensiones,
pero no así en el largo de la lesura, que en
este caso es de 2/3 del radio. La mayor afinidad se ha
encontrado con C. minor Couper (1953), pero su diámetro
es menor al mínimo citado para esa especie, y
además tiene una exina más delgada (1,2 µm) que la
allí citada (1,5 µm).
Afinidad botánica. Se vincula este tipo de esporas
con las familias Gleicheniaceae, Dipteridaceae y
Cyatheaceae (Balme, 1995).
Material estudiado. PBN N1: BA Pal 5571 y 5574.
Subturma ZONOTRILETES Waltz 1935
Infraturma CINGULATI Potonié y Klaus 1954Género Densoisporites (Weyland y Krieger) emend. Dettmann 1963
Especie tipo. Densoisporites velatus Weyland y Krieger 1953.
Afinidad botánica. Clase Lycopsida (Balme, 1995).
Densoisporites velatus (Weyland y Krieger) emend.
Krasnova (ex. Dettmann 1963) Figura 3.4

Figura 3. Esporas cinguladas / cingulate spores; 1-3, Densoisporites corrugatus Archangelsky y Gamerro; 1-2, MO / LM, 1, cara proximal
/ proximal view y / and 2, cara distal / distal view (Pm BA Pal Nº 5500: 12.5/99.8); 3, fotografía tomada con MEB / SEM; 4, Densoisporites velatus (Weyland y Krieger) emend. Krasnova (ex. Dettmann, 1963) (Pm BA Pal Nº 5500: 8.1/99.1); 5, Foraminisporis dailyi (Cookson y
Dettmann) Dettmann (Pm BA Pal Nº 5566: 4.7/106); 6, Foraminisporis sp. (Pm BA Pal Nº 5566: 6.8/106,4); 7, Contignisporites glebulentus (Dettmann) emend. Filatoff y Price (Pm BA Pal Nº 5577: 19.6/93.1); 8, Contignisporites burgeri Filatoff, Mc Kellar y Price: cara proximal
mostrando las facetas ornamentadas / proximal view showing ornamentation (Pm BA Pal Nº 5577: 19.8/89.8); 9, Contignisporites cooksoniae (Balme) Dettmann (Pm BA Pal Nº 5577: 20.5/90.9). Escala / scale bar: 10 µm.
1958. D. perinatus Couper, p. 145, lám. XXIII, figs.6-9.
1963. D. microrugulatus Brenner, p. 61, lám. XV, fig. 6 y lám. XVI,
fig. 1.
1965. D. velatus Weyland y Krieger emend. Krasnova sensu Dettmann; Archangelsky y Gamerro, p. 162, lám. 1-4.
2003. D. velatus Weyland y Krieger; Archangelsky y Llorens, p. 76,
fig. 2J.
Descripción. Esporas trilete de contorno subtriangular
a circular y lados convexos; cara distal convexa y
proximal subcónica. La lesura varía en extensión y
nitidez, pudiendo ser poco visible o, en los casos
donde es más evidente, presentar un engrosamiento.
Las ramas son levemente sinuosas y se extienden
hasta el borde interno del cíngulo. Exina compuesta
por dos capas con una separación variable entre ellas,
excepto en la cara proximal. La interna es de un espesor
homogéneo en toda su superficie, o bien puede
presentar tres engrosamientos circulares o elípticos
cerca del polo proximal; la capa externa tiene una
textura microalveolar, y se encuentra engrosada a nivel
del ecuador. Los alvéolos son más amplios en la
cara distal que en la proximal, y son elongados y de
mayor tamaño aún en el cíngulo, dándole a éste un
aspecto estriado-esponjoso. Frecuentemente el cíngulo
se encuentra engrosado en los ángulos.
Dimensiones. Diámetro ecuatorial (77 ejemplares)
26,7 (30) 75 µm; diámetro del cuerpo central (26 ejemplares)
22 (26) 30 µm; espesor del cíngulo (37 ejemplares)
1,5 (2,5) 4 µm; lados (8 ejemplares) 2 (2,6) 4
µm, ángulos 3 (4,5) 5,3 µm.
Observaciones. Cabe destacar el amplio rango de tamaños
observados dentro de esta forma polínica. No
obstante, sólo se registran unos pocos ejemplares con
más de 48 µm de diámetro, concentrándose la moda
dentro de un rango reducido (30-34 µm).
Discusión y comparaciones. El género Densoisporites fue enmendado por Dettmann (1963), quien agregó
como carácter diagnóstico la presencia de tres engrosamientos
(papilas) en la capa interna de la exina, cerca
del polo proximal. A tal respecto, Archangelsky y
Gamerro (1965) aclaran para su material de la
Formación Baqueró, que estas papilas no siempre son
visibles, situación que se repite en este material. En
comparación con los ejemplares estudiados por estos
autores para la unidad basal de este Grupo, las medidas
del presente material son considerablemente menores.
Es así que el diámetro ecuatorial varía de 40-
80,5µm en los especímenes de la unidad basal, a 26,7-
75µm en los de la unidad cuspidal del Grupo Baqueró;
siendo el diámetro del cuerpo central de 28,6-68,2 µm
(unidad basal) y 22,3-30 µm (unidad cuspidal) y el espesor
del cíngulo de 2,8-10,8 µm a 1,6-4,2 µm. La variación
entre las medidas puede deberse a las técnicas
de preparación utilizadas en cada caso, a la naturaleza
esponjosa de la exina, o bien a factores ecológicos.
Material estudiado. PBN N1: BA Pal 5500, 5501, 5503, 5550, 5551, 5552, 5557, 5555, 5558, 5567, 5568, 5569, 5572 y 5574; N3: MPEF Palin 60 y 61.
Densoisporites corrugatus Archangelsky y Gamerro, 1965 Figura 3.1-3.3
1965. D. corrugatus Archangelsky y Gamerro, p. 163, lám. II, figs.
1-11.
2003. D. corrugatus Archangelsky y Gamerro; A. Archangelsky y
Llorens, p. 76, fig. 2K.
Dimensiones. Diámetro total (59 ejemplares) 34
(45,5) 59 µm; diámetro del cuerpo interno (41 ejemplares)
23,5 (35,5) 44 µm; espesor del cíngulo: lados
(36 ejemplares) 3 (6,5) 7,5 µm, ángulos (20 ejemplares)
3 (5,5) 7 µm; diámetro polar (1 ejemplar) 34 µm.
Discusión. Si bien esta forma coincide con la descripta
por Archangelsky y Gamerro (1965) para la
Formación Baqueró, cabe mencionar algunos aspectos
diferentes que presenta este material. En primer
lugar, un tamaño menor (34-59 µm) que el descripto
en aquella ocasión (52,7-73 µm). Como en el caso de D. velatus, esa variación en el diámetro puede deberse
en parte a las técnicas originales de preparación,
más oxidantes que las actuales. Por otra parte, se
menciona un ensanchamiento del cíngulo en los ángulos;
en este aspecto, el material es muy variable,
pudiendo presentarse sin ensanchamientos o bien
con los lados más anchos.
Observaciones. En la diagnosis original se da como
carácter diagnóstico la presencia de tres engrosamientos
circulares o elípticos (papilas) de la capa interna
de la exina en el polo proximal; este carácter está
presente en nuestro material sólo en un 50% de los
casos.
Distribución estratiráfica. Esta especie ha sido citada
en el Aptiano-Albiano de la cuenca Austral
(Archangelsky y Gamerro, 1965; Archangelsky y
Llorens, 2003) y en el Barremiano-Aptiano de
Patagonia austral (Archangelsky et al., 1984).
Material estudiado. PBN N1: BA Pal 5500, 5501, 5503, 5550, 5557, 5564, 5567, 5568, 5569, 5572 y 5574; N3: MPEF Palin 60 y 61; PBS N4: MPEF Palin 62.
Género Foraminisporis Krutzsch 1959 (sensu Dettmann 1963)
Especie tipo. Foraminisporis foraminis Krutzsch 1959.
Afinidad botánica. Dettmann (1963) plantea la similitud de estas esporas con las de algunas especies de hepáticas actuales.
Foraminisporis dailyi (Cookson y Dettmann) Dettmann 1963 Figura 3.5
Dimensiones. (9 ejemplares) diámetro ecuatorial 16- 38 µm; espesor de la exina: lados 1,5-3 µm; espesor capa interna de la exina 0,5-1 µm.
Material estudiado. PBN N1: BA Pal 5566 y 5577; N3: MPEF Palin 60; PBS N4: MPEF Palin 57 y 58.
Foraminisporis sp. Figura 3.6
Descripción. Esporas trilete cinguladas de contorno
ecuatorial subcircular. La lesura es simple, porta algunos
gránulos pequeños sólo observables cuando
está cerrada; se extiende hasta el ecuador, abarcando
también el cíngulo. Este último es angosto y presenta
unas pocas fósulas dispuestas ampliamente y de
manera irregular. Exina con dos capas, la interna de
un espesor uniforme; la externa bastante lisa, sólo
con algunos gránulos dispersos y desordenados de
no más de 0,6 µm de base.
Dimensiones. (2 ejemplares) diámetro ecuatorial
22,5-27,5 µm; espesor de la capa interna de la exina
0,8-1 µm; espesor del cíngulo 1,5-2 µm.
Discusión y comparaciones. Dettmann (1963) considera
que el engrosamiento de la exina en el ecuador
es, a diferencia de la diagnosis original, un carácter
distintivo; aunque no es fácilmente distinguible de
no ser en ejemplares que se encuentren en una vista
netamente polar. También destaca que los ejemplares
australianos sólo adquieren esa posición en preparados
microscópicos montados en un 50% de glicerina.
Por tanto considero que esta forma, al presentar esa
característica debe ser incluida en este género, sin poder
llegar a una asignación específica.
Material estudiado. PBN N1: BA Pal 5552 y 5566.
Género Contignisporites Dettman 1963 emend. Filatoff y Price 1988
Especie tipo. Contignisporites glebulentus Dettmann 1963.
Observaciones. Las tres especies del género
Contignisporites aquí descriptas exhiben una diferencia
en las dimensiones con respecto a las diagnosis
originales: siempre son de menor tamaño. Sin embargo,
no se considera al tamaño como un carácter distintivo
por ser una situación que se repite en varios taxones
de esta microflora. Cabe destacar también que
existen diferencias en la medida ∆-ratio, a pesar de
que ha sido concebida como un índice que tiende a
compensar la variabilidad intrapoblacional y los cambios
influenciados por las condiciones postdepositacionales
(Filatoff y Price, 1988). Este valor representa
el cociente entre diámetro ecuatorial y la suma de 4
muros más sus respectivas zonas de separación.
Afinidad botánica. Dentro de las formas actuales se
encuentra una afinidad con las esporas del género Pityrogramma Link (Pteridaceae) (Tryon y Lugardon,
1990: 129 fig. 34).
Contignisporites glebulentus (Dettmann) emend. Filatoff y Price 1988 Figura 3.7
1963. Contignisporites glebulentus Dettmann, p. 74, pl. XV, fig. 1-10
y texto-fig. 5.
1988. C. glebulentus Dettmann; Filatoff y Price, fig. 7 E-P. (Véase en
este trabajo la sinonimia completa).
Dimensiones. (2 ejemplares) diámetro ecuatorial 36-
38 µm; muros distales: alto 1,5-2,4 µm y ancho 1,8
µm; separación entre muros 1 µm; espesor del cíngulo
2,9-3,3 µm; ∆-ratio 2-2,2.
Distribución estratigráfica. Ampliamente distribuida
entre el Jurásico Medio y el Cretácico Superior de
Gondwana (Sajjadi y Playford, 2002). En el país ha sido
reportada únicamente por A. Archangelsky y
Llorens (2003) para el Albiano de la Cuenca Austral
(Formación Kachaike).
Material estudiado. PBN N1: BA Pal 5577.
Contignisporites burgueri Filatoff, Mc Kellar y Price (en: Filatoff y Price, 1988) Figura 3.8
1983. C. cooksonii (Balme) Dettmann; Baldoni y Archangelsky, p.
65, lám. 4, figs. 5 y 6.
1988. Contignisporites burgueri Filatoff y Price, Fig. 8 A-H.
2003. C. burgueri Filatoff y Price; A. Archangelsky y Llorens, p. 76,
fig. 3 B.
Dimensiones. (19 ejemplares) diámetro ecuatorial 24
(32) 37 µm; diámetro polar (2 ejemplares) 21,7-28 µm;
muros distales: alto 1,3-2,3 µm y ancho 1,5-2,8 µm; separación
entre muros 0,8-3,2 µm; espesor del cíngulo
2 (2,7) 4 µm; ∆-ratio 1,8-3,6.
Discusión y comparaciones. Los ejemplares aquí
descriptos presentan menores dimensiones que las
descriptas anteriormente por Filatoff y Price (1988).
Esa situación es particularmente notoria en esta especie,
dado que se han encontrado más individuos
que en las otras y todo el conjunto es muy homogéneo
en sus dimensiones.
Distribución estratigráfica. Baldoni y Archangelsky
(1983) registraron esta especie para la Fm. Springhill,
aunque en esa ocasión la asignaron a C. cooksonii, en
la ilustración allí presentada se observan claramente
los elementos esculturales de las facetas proximales
que son propios de C. burgueri. El primer reporte formal
de esta especie para Argentina fue para el
Albiano de la Cuenca Austral (A. Archangelsky y
Llorens, 2003). Sajjadi y Playford (2002) describen para
esta especie un rango que va del Jurásico Medio al
Cretácico Superior de Australia.
Material estudiado. PBN N1: BA Pal 5569, 5572, 5574, 5575 y 5577.
Contignisporites cooksoniae (Balme) Dettmann 1963 Figura 3.9
1963. Contignisporites cooksonii Balme; Dettmann, p. 75, lám. XV,
fig. 11-16.
1988. C. cooksoniae (Balme) Dettmann; Filatoff y Price, fig. 7 E-P.
(Véase en este trabajo la sinonimia completa).
Dimensiones. (6 ejemplares) diámetro ecuatorial 24
(32) 46,8 µm; espesor del cíngulo 1,7-4 µm; muros distales:
alto 1-1,8 µm y ancho 1,5-2,9 µm; separación entre
muros 0,8-2,4 µm; ∆-ratio 2-2,9.
Distribución estratigráfica. En Argentina esta especie
fue descripta previamente por Volkheimer y
Quattrocchio (1975) para el Jurásico de la Cuenca
Neuquina, por Archangelsky y Gamerro (1966) para
el Grupo Baqueró y por A. Archangelsky y Llorens
(2003) para el Albiano de Santa Cruz (Fm. Kachaike).
Fue descripta también para el Cretácico Medio de la
Cuenca del Golfo San Jorge (Archangelsky et al.,
1994). Presente en el Cretácico de Australia (Filatoff y
Price, 1988).
Material estudiado. PBN N1: BA Pal 5568, 5572, 5574 y 5577.
Turma MONOLETES Ibrahim 1933
Subturma AZONOMONOLETES Luber 1935
Infraturma LAEVIGATOMONOLETI Dybová y Jachowicz1957Género Laevigatosporites Ibrahim 1933
Laevigatosporites ovatus Wilson y Webster 1946 Figura 2.9
Dimensiones. Diámetro ecuatorial (2 ejemplares) 23-
30 µm; diámetro polar 16-20 µm; espesor de la exina
1-1,5 µm.
Distribución estratigráfica. Esta especie ha sido ampliamente
citada para el Cretácico y Terciario en todo
el mundo.
Material estudiado. PBN N1: BA Pal 5570; PBN N3: MPEF Palin 61.
Laevigatosporites sp. Figura 2.10
Descripción. Esporas monolete de grandes dimensiones,
con contorno ovalado en vista polar y planoconvexo
a biconvexo en vista ecuatorial. La lesura es
simple y recta, con una apariencia de corte, que no
llega a abarcar la mitad de la longitud máxima de la
espora. La exina es lisa y rígida, de un espesor de 1,5-
3,2 µm, con pliegues secundarios. A menudo se observa
un pliegue de la exina paralelo al ecuador que
puede confundirse con una capa interna.
Dimensiones. Diámetro ecuatorial (15 ejemplares)
largo: 67 (98,6) 106 µm, ancho: 36 (51,2) 58,2 µm; lesura
(11 ejemplares) 24 (34,2) 40,7 µm; espesor de la
exina (15 ejemplares) 1,5 (2,2) 3,2 µm.
Discusión y comparaciones. Los ejemplares estudiados
son muy semejantes a L. belfordii descripto por
Burger (1984) para el Neocomiano-Aptiano de
Australia y citado por Archangelsky et al. (1983) para
el Cretácico de Argentina austral, aunque en el material
aquí estudiado la lesura es simple, por lo que no
puede incluirse en ese taxón. El resto de las especies afines, tienen un rango de tamaños mucho menor y
además siempre muestran una forma arriñonada en
vista lateral. Los ejemplares que Pflug (en Thompson
y Pflug, 1953) describe como L. discordatus no muestran
ese carácter como tampoco un labio, pero su rango
de diámetro ecuatorial es menor (50-90 µm) que el
aquí descripto.
Material estudiado. PBN N1: BA Pal 5500, 5503, 5550, 5569 y 5572.
Infraturma SCULPTATOMONOLETI Dybová y Jachowicz 1957
Género Reticulosporis Krutszch 1959
1961. Retimonoletes Pierce, p. 33.
1988. Reticulatamonoletes Lu Li Chang, p. 221.
Especie tipo. Reticulosporis miocenicus (Selling) Krtz, Ibid. Schizaea miocenica Selling 1944, p. 68, pl. 4, fig. 46 Sporites secundus Pot. In Thiergart 1937, p. 296, pl.22, fig. 16=holotype; non Potonié 1934, p. 39, pl. 6, fig. 7.
Discusión y comparaciones. Una forma muy relacionada
a esta, es el morfogénero Microfoveolatosporis
Krutzsch 1959. Srivastava (1971) ha considerado a
Reticulosporis como un sinónimo junior de Microfoveolatosporis, en base a la descripción de su especie
tipo. Aquí no se sigue tal criterio y en cambio se
refiere a la diagnosis genérica original de ambos
morfogéneros (Krutzsch, 1959), sin tener en cuenta la
descripción de las especies tipo que pueden solaparse,
ya que introducen nomenclaturas poco claras. En
tanto, las diagnosis genéricas claramente describen
distintos tipos de esculturación de la pared: un retículo
positivo regular para Reticulosporis, y una esculturación
foveolada en Microfoveolatosporis. Las tres
formas aquí descriptas presentan una pared ornamentada
por muros que forman un retículo regular
positivo, y esto las enmarca dentro del primer morfogénero
mencionado, aunque es importante señalar
la incompatibilidad existente con la descripción de la
especie tipo, cuya ornamentación es "alveolar".
Pierce (1961), realizó una somera diagnosis del género Retimonoletes, al que describió como "reticulate,
monolete spores". Así, debe considerársele como un
sinónimo junior de Reticulosporis Krutzsch. Otra forma
relacionada es el género Reticulatamonoletes, al
que Lu Li Chang (1988) ha definido como formas monoletes
con muros conspicuos que forman retículos
amplios (referencia tomada de Jansonius y Hill, 1992:
4951). Por lo anteriormente expresado, esta forma
puede incluirse en Reticulosporis.
Distribución estratigráfica. Es esta la primer mención
del género para la Argentina. Cabe señalar
además, que se trata de una de las menciones más
antiguas del género, ya que ha sido registrado mayormente
en sedimentos terciarios o bien del Cretácico
Superior. La mención más antigua corresponde
al Cretácico bajo de Checoslovaquia (Vavrdova,
1964).
Reticulosporis sp. 1 Figura 2.11
Descripción. Espora monolete concavo-convexa y
forma oval en vista ecuatorial. Exina delgada, ornamentada
con muros bajos -0,7 µm de alto- y delgados
-0,5 µm de ancho- que forman un retículo regular de
lúminas grandes penta o hexagonales con un diámetro
de 3,5 - 5 µm.
Dimensiones. (1 ejemplar) diámetro ecuatorial 22
µm; diámetro polar 16 µm; espesor de la exina (incluida
la escultura) 1,3 µm.; diámetro de los alvéolos
3,5-5µm.
Material estudiado. PBN N1: BA Pal 5552.
Comparaciones. Este tipo de espora es muy diferente a la especie tipo del género puesto que ella está caracterizada por una pared gruesa, con una estructura radial o granular, que le confiere un aspecto foveolado, a diferencia de los muros que definen claramente el retículo en R. sp. 1. Por otra parte, dentro de las formas aquí descriptas, ésta es la más disímil por poseer muros bajos y el retículo más amplio.
Reticulosporis sp. 2 Figura 2.12
Descripción. Espora monolete cóncavo-conexa de
forma oval en vista ecuatorial. Exina gruesa, esculturada
con muros elevados y robustos (2 µm) de un ancho
irregular (0,7-1,5 µm) que forman un retículo
apretado con pequeñas lúminas penta-hexagonales.
Dimensiones. (1 ejemplar) diámetro ecuatorial 33
µm; diámetro polar 23 µm; espesor de la exina (incluida
la ornamentación) 3,5 µm; alto de los muros 2
µm; diámetro de los alvéolos 2,5-3 µm.
Material estudiado. PBN N1: BA Pal 5567.
Comparaciones. Esta espora se diferencia de Reticulosporis sp. 1 y R. sp. 3 por su retículo apretado y el ancho irregular de sus muros de aspecto robusto.
Reticulosporis sp. 3 Figura 2.13
Descripción. Espora monolete cóncavo-conexa de
forma oval en vista ecuatorial. Exina gruesa ornamentada
por muros delicados muy delgados (0,5 µm)
y elevados (3 µm) que forman un retículo amplio con
lúminas de forma penta o hexagonales.
Dimensiones. (1 ejemplar) diámetro ecuatorial 22
µm; diámetro polar 16 µm; espesor de la exina 1,5
µm; diámetro de los alvéolos 4-4,5 µm.
Material estudiado. PBN N1: BA Pal 5569.
Comparaciones. La particularidad de esta espora con respecto a las antes descriptas, reside en que sus muros son elevados y finos, con un aspecto translúcido.
Discusión
En esta publicación se presenta la primera parte
del estudio sistemático de las microsporas dispersas
de la Formación Punta del Barco, con la ubicación del
área de estudio y de los niveles polínicos portadores
de esta microflora. Los grupos morfológicos abordados
corresponden a la turma Monoletes (5 especies) y
las infraturmas Laevigati (8 especies) y Cingulati (7
especies) dentro de la turma Triletes, sumando 20 las
especies descriptas.
La forma más abundante de esta asociación de
esporas es la correspondiente al género
Densoisporites (30,9%), con dos especies representadas:
D. velatus y D. corrugatus. Su afinidad botánica
la vincula con las Lycophyta, grupo botánico previamente
reportado para la Formación Punta del Barco
mediante el registro de megasporas (Llorens y
Morbelli, 2006). Su abundancia relativa dentro de la
microflora estaría relacionada a los ambientes acuáticos
que este tipo de plantas ocupó, concediendo
una sobre-representación de las plantas productoras
en la palinoflora.
Esta es la primera referencia de la División
Bryophyta para la unidad, ilustrada por las esporas
del morfogénero Foraminisporis. Asimismo, dentro de
las Filicales no se había citado previamente la participación
de la Familia Osmundaceae en esta tafoflora,
que es sugerida por la presencia de esporas de esa
afinidad botánica, de los géneros Todisporites y
Biretisporites, este último constituye el 10,23% del total
de esporas recuperadas. De igual manera, en esta
publicación se da conocer por primera vez la presencia
de la Familia Pteridaceae, que está muy bien documentada
por tres especies del género Contignisporites
que constituyen el 6,14% de las esporas de
esta microflora.
Es de destacar la diversidad de esporas monoletes
en esta asociación polínica. Las esporas monoletes
con esculturación en su exina son poco frecuentes
en el registro palinológico del Mesozoico, y se
trata generalmente de muy escasos ejemplares. Este
hecho impide en muchos casos, como en este, hacer
asignaciones específicas; sin embargo es interesante
darlos a conocer, ya que se trata de formas peculiares
que podrían tener valor bioestratigráfico. En tal
sentido, este es el primer reporte para el país del género
Reticulosporis Krutszch, representado por tres
formas diferentes. Ese morfogénero a su vez, había
sido previamente citado para sedimentos cretácicos
tardíos y terciarios de todo el mundo. De esta manera,
ésta se constituye en una de las menciones
más antigua para ese taxón, junto con la que realizara
Vavrdova (1964) para el Cretácico bajo de Checoslovaquia.
Agradecimientos
Este trabajo se realizó en el marco del proyecto de investigación "Cretácico Inferior de Patagonia Austral: Fitoestratigrafía de áreas seleccionadas" (PICT 99-6044). La autora desea agradecer a S. Archangelsky por la lectura crítica del manuscrito, y a A. Archangelsky por la cesión de su microscopio.
Bibliografía
1. Archangelsky, A. y Llorens, M. 2003. Palinología de la Formación Kachaike, Cretácico Inferior de la Cuenca Austral, provincia de Santa Cruz. 1- Esporas lisas y cinguladas. Ameghiniana 40: 71-80.
2. Archangelsky, A., Andreis, R.R., Archangelsky, S. y Artabe, A. 1995. Cuticular characters adapted to volcanic stress in a new Cretaceous cycad leaf from Patagonia, Argentina. Considerations on the stratigraphy and depositional history of the Baqueró Formation. Review of Palaeobotany and Palynology 89: 213-233.
3. Archangelsky, S. 1967. Estudio de la Formación Baqueró, Cretácico inferior de Santa Cruz, Argentina. Revista del Museo de La Plata. Paleontología 32: 63-170.
4. Archangelsky, S. 1972. Esporas de la Formación Río Turbio (Eoceno), provincia de Santa Cruz. Revista del Museo de La Plata (Nueva Serie) Paleontología 6: 65-100.
5. Archangelsky, S. 2001. Evidences of an Early Cretaceous Floristic change in Patagonia, Argentina. 7th International Symposium on Mesozoic Terrestrial Ecosystems, Asociación Paleontológica Argentina, Publicación Especial 7: 15-19.
6. Archangelsky, S. 2003. Flora Cretácica del Grupo Baqueró. Ed.: Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" (en CD).
7. Archangelsky, S. y Gamerro, J.C. 1965. Estudio palinológico de la Formación Baqueró (Cretácico), Provincia de Santa Cruz I. Ameghiniana 4: 159-170.
8. Archangelsky, S. y Gamerro, J.C. 1966. Estudio palinológico de la Formación Baqueró (Cretácico), Provincia de Santa Cruz II. Ameghiniana 4: 201-209.
9. Archangelsky, S. y Villar de Seoane, L. 1994. Estudio palinológico de la Formación Baqueró (Cretácico), Provincia de Santa Cruz, Argentina. VI. Ameghiniana 31: 41-53.
10. Archangelsky, S., Baldoni A.M., Gamerro J. C. y Seiler J. 1983. Palinología estratigráfica del Cretácico de Argentina Austral. 2. Descripciones sistemáticas. Ameghiniana 20: 199-226.
11. Archangelsky, S., Baldoni, A.M., Gamerro J.C. y Seiler, J. 1984. Palinología estratigráfica del Cretácico de Argentina austral. 3 Distribución de las especies y conclusiones. Ameghiniana 21: 15-33.
12. Archangelsky, S., Bellosi E.S., Jalfin G.A. y Perrot C. 1994. Palynology and alluvial facies from the mid-Cretaceous of Patagonia, subsurface of San Jorge Basin, Argentina. Cretaceous Research 15: 127-142.
13. Baldoni, A.M. y Archangelsky, S. 1983. Palinología de la Formación Springhill (Cretácico Inferior), Subsuelo de Argentina y Chile Austral. Revista Española de Micropaleontología 15: 47- 101.
14. Balme, B.E. 1995. Fossil in situ Spores and Pollen grains: An annotated Catalogue. Review of Palaeobotany and Palynology 87: 81-322.
15. Berry, E.W. 1924. Mesozoic Plants from Patagonia. American Journal of Science 7: 473-482.
16. Brenner, G.J. 1963. The spores and pollen of the Potomac Group of Maryland. Maryland Board Natural Resources, Department of Geology, Mines and Water Resources, Bulletin 27: 1- 215.
17. Burger, D. 1984. Palynology of subsurface Lower Cretaceous strata in the Surat Basin, Queensland. Bureau of Mineral Resources, Bulletin 150: 1-16.
18. Cladera, G., Andreis, R. y Archangelsky, S. y Cúneo, R. 2002. Estratigrafía del Grupo Baqueró, Patagonia (Provincia de Santa Cruz, Argentina). Ameghiniana 39: 3-20.
19. Cladera G.A. y Cúneo R. 2002. Fossil plants buried by volcanic ash in the Lower Cretaceous of Patagonia. Current Topics on Taphonomy and Fossilization, 399-403.
20. Corbella, H. 2001. Tuffs of the Baqueró Group and the Mid- Cretaceous frame Extraandean Patagonia, Argentina. 11º Congreso Latinoamericano de Geología y 3º Congreso Uruguayo de Geología. Trabajo 190, 6 p. (en CD).
21. Couper, R.A. 1953. Upper Mesozoic and Cainozoic Spores and Pollen Grain from New Zealand. Paleontological Bulletin of the New Zealand Geological Survey 22: 1-77.
22. Couper, R.A. 1958. British Mesozoic Microspores and Pollen Grain (A Systematic and Stratigraphic study). Palaeontographica B 103: 79-117.
23. Delcourt, A.R. y Sprumont, G. 1955. Les spores et grains du pollen du Wealdien du Hainaut. Mémoires de la Société belge de Geologie 5: 1-73.
24. Delcourt, A.F., Dettmann, M.E. y Hughes, N.F. 1963. Revision of some Lower Cretaceous Microspores from Belgium. Palaeontology 6: 282-292.
25. Del Fueyo, G.M. y Archangelsky, A. 2002. Araucaria grandifolia Feruglio from the Lower Cretaceous of Patagonia. Cretaceous Research 23: 255-267.
26. Dettmann, M.E. 1963. Upper Mesozoic microfloras from Southeastern Australia. Royal Society of Victoria 77: 1-148.
27. Feruglio, E. 1951. Piante del Mesozoico de la Patagonia. Publicación Instituto Geológico Universidad de Torino 1: 35-80.
28. Filatoff, J. y Price P.L. 1988. A Pteridacean spore lineage in the Australian Mesozoic. Memorial Association of Australasian Palaeontologists 5: 89-124.
29. Herbst, R. 1960. Descripción de algunos ejemplares fértiles de Hausmannia (Protorhipis) papilio Fer., procedentes de los estratos de Baqueró, Santa Cruz. Acta Geológica Lilloana 3: 227-232.
30. Herbst, R. 1962. Sobre especies de Gleichenites de los sedimentos baqueroenses de Santa Cruz, Patagonia. Ameghiniana 2: 141-151.
31. Herbst, R. 1966. Nuevos elementos de la flora fósil de la Formación Baqueró, Santa Cruz, Patagonia. Acta Geológica Lilloana 8 : 75-89.
32. Ibrahim, A.C. 1933. Sporenformen des Aegir-horizonts des Ruhr- Reviers. Dissertation, University of Berlin, Konrad Triltsch (ed.), Wurzburg, 47 pp.
33. Jansonius J. y Hills L.V. 1976. Genera File of Fossil Spores. Special Publication of the Department of Geology, University of Calgary, 748, 1472.
34. Jansonius J. y Hills L.V. 1992. Genera File of Fossil Spores - Supplement 11. Special Publication of the Department of Geology, University of Calgary, 4951.
35. Krutzsch, W. 1959. Mikropaläeontogische (sporenpaläeontogische) Untersuchungen in der Braunkohle des Geiseltales. Geologie 8: 1-425.
36. Llorens, M. 2000. Estudio Palinológico de la Formación Punta del Barco (Grupo Baqueró) Cretácico Inferior, provincia de Santa Cruz. 11º Simposio Argentino de Paleobotánica y Palinología, resúmenes: 56.
37. Llorens, M. 2003a. Polen de angiospermas de la Formación Punta del Barco (Grupo Baqueró), Aptiano de Santa Cruz, Argentina. Ameghiniana Suplemento Resúmenes 40: 27R.
38. Llorens, M. 2003b. Granos de polen de angiospermas de la Formación Punta del Barco (Aptiano), provincia de Santa Cruz, Argentina. Revista del Museo Argentino de Ciencias Naturales, nueva serie 5: 235-240.
39. Llorens, M. 2005. ( Palinología de la Formación Punta del Barco, Cretácico Inferior de Santa Cruz. Tesis doctoral, Universidad Nacional de La Plata, 210 pp. Inédita.).
40. Llorens, M. y Cladera G. 2002. Esporas perisporadas como evidencia tafonómica. 8º Congreso Argentino de Paleontología y Bioestratigrafía. Resúmenes: pp. 73.
41. Llorens, M. y Morbelli, M. A., 2006. Megasporas de la Formación Punta del Barco, Aptiano de Santa Cruz. 13º Simposio de Paleobotánica y Palinología. Actas: 49.
42. Llorens, M., Passalia M.G. y Cladera, G. 2003. Nuevos aportes a la flora de la Formación Punta del Barco (Grupo Baqueró) Aptiano de Santa Cruz, Argentina. Ameghiniana Suplemento Resúmenes 40: 28R.
43. Lu, L.C. 1988. Middle Devonian Microflora from Haickou Formation at Shijiapo in Zhanyi of Yunnan, China. Memoirs of the Nanjing Institute of Geology and Palaeontology, Academia Sinica 24: 109-222.
44. Miner, E.L. 1935: Paleobotaical examination of Cretaceous and Tertiary coals. American Midland Naturalist 16: 585-625.
45. Pierce, R.L. 1961. Lower Upper Cretaceous plant microfossils from Minesota. Bulletin of the Geological Survey of University of Minnesota 42: 1-86.
46. Potonié, R. 1934. Zur Mikrobotanik des eocanen Humodils des Geiseltals. Arbeiten aus dem Institut fur Palaobotanik und Petrographie der Brennsteine 4: 25-125.
47. Potonié, R. 1956. Synopsis der Gattungen der Sporae dispersae. I. Teil: Sporites. Beihefte zum Geologischen Jahrbuch 23: 103 p.
48. Potonié, R. y Kremp, G. O. W. 1954. Die Gattungen der palaozoischen Sporae dispersae und ihre Stratigraphie. Geologisches Jahrbuch 69: 111-194.
49. Prámparo, M.B. 1988. Esporas triletes levigadas y apiculadas de la Formación La Cantera (Cretácico de la Cuenca de San Luis) en su localidad tipo. 4º Congreso Argentino de Paleontología y Bioestratigrafía (1986), Actas 3: 51-62.
50. Sajjadi, F. y Playford, G. 2002. Systematic and stratigraphic palynology of Late Jurassic-earliest Cretaceous strata of the Eromanga Basin, Queensland, Australia: Part One. Palaeontographica B 261: 1-98.
51. Srivastava, S.K. 1971. Monolete spores from the Edmonton Formation (Maestrichtian), Alberta, Canada. Review of Palaeobotany and Palynology 11: 251-265.
52. Thiergart, F. 1937. Die Pollenflora der Niederlausitzer Braunkohle, besonders im Profil der Grube Marga bei Senftenberg. Jahrbuch der Preussische geologischen Landesanstalt und Bergakademie zu Berlin 58: 282-356.
53. Thomson, P.W. y Pflug, H. 1953. Pollen und Sporen des mitteleuropäischen Tertiärs: Gesamtübersicht über die stratigraphisch uns paläontologisch wichtigen Formen. Palaeontographica B, 94: 1-138.
54. Tryon, A.F. y Lugardon, B. 1990. Spores of the Pteridophyta. Springer- Berlag, New York, 648 pp.
55. Vavrdova, M. 1964. Fossil Microplankton From The Tesin- Hradiste Series (Lower Cretaceous).Part 1-Dinoflagellates. Sbornik Geologickych Ved,Paleontologie Volume 4: 91-104.
56. Volkheimer, W. y Quattrocchio, M. 1975. Palinología Estratigráfica del Titoniano (Formación Vaca Muerta) en el área de Caichigüe (Cuenca Neuquina). Ameghiniana 12: 193-241.
57. Weyland, H. y Krieger, W. 1953. Die Sporen und Pollen Aachener Kreide und ihre Bedeutung für die Charakterisierung des Mittleren Senons. Palaeontographica B 95: 6-29.
58. Wilson, L.R. y Webster R.M. 1946. Plant microfossils from a Fort Union Coal of Montana. American Journal of Botany 33: 271-278.
59. Wingate, F.H. 1980. Plant Microfossils from the Denton Shale Member of the Bokchito Formation (Lower Cretaceous, Albian) in Southern Oklahoma. Oklahoma Geological Survey Bulletin 130: 1-93.
Recibido: 13 de marzo de 2006.
Aceptado: 4 de diciembre de 2007.