
Fig.1. Mapa de localización del área de estudio "Parque Nacional Predelta", Localidad de Diamante, Entre Ríos, Argentina.
PALEOBOTÁNICA Y PALINOLOGÍA
Análisis fitolíticos en depósitos de ambientes fluviales del río Paraná en su sección media (Entre Ríos, Argentina)
Noelia Isabel Patterer1,2*, Carlos Adrián González1,2, Esteban Passeggi1,2 y Alejandro Fabián Zucol1,2
1 Laboratorio de Paleobotánica, Centro de Investigaciones
Científicas (CICYTTP-CONICET), Dr. Matteri y España
(E3105BWA) Diamante, Entre Ríos, Argentina.
2 Facultad de Ciencia y Tecnología, Universidad Autónoma
de Entre Ríos, Argentina.
* noepatt1@hotmail.com
Resumen: El presente trabajo define la composición fitolítica de distintos ambientes insulares de la región más austral del Paraná medio, estableciendo características florísticas y fluviosedimentarias del sistema planta-sustrato. El Paraná medio, se desplaza por una llanura aluvial (faja inundable compleja y de gran tamaño). Los análisis de fitolitos han mostrado ser una valiosa contribución para comprender el tipo de ambiente en que se desarrollaron los depósitos fluviales recientes, tanto para el estudio de los depósitos de fondo, como de los ambientes insulares vinculados. Se establecieron 6 estaciones, denominadas: Albardón viejo, Albardón intermedio, Albardón joven, Media loma, Pajonal y Bajo. El procesamiento se realizó mediante la técnica de Laboriau (1983) para las muestras vegetales y se siguió el protocolo de procesamiento de Zucol et al. (2010), para las muestras clásticas. El análisis fitolítico resultó ser una herramienta útil, ya que nos permitió marcar diferencias significativas en las diferentes unidades de estudio, caracterizando de forma clara los componentes actuales y su contribución al sustrato, como así también, aquellos elementos que han distinguido estos ambientes en tiempos pasados recientes.
Palabras clave: Análisis fitolíticos; Río Paraná medio; Parque Nacional Predelta; Provincia de Entre Ríos.
Summary: Phytolith analysis in fluvial deposits of ambiences of the middle reach Parana river (Entre Ríos, Argentina). This paper defines the phytoliths composition from different environments of the insular region southernmost Middle Paraná River, establishing fluvio-sedimentary and floristic characteristics, of the plant-substrate associations. The Middle Paraná, moves through a floodplain. The phytolith analysis have proved a valuable contribution to understanding the type of environment, both for the study of bottom deposits as well as such as island environments. Six stations were established: "albardón joven", "albardón intermedio", "albardón viejo", "media loma", "pajonal" and "bajo". The processing was performed using the technique of Laboriau (1980) for plant samples and followed the protocol processing Zucol et al. (2010) for clastic samples. Phytolith analysis proved a useful tool, because we could make significant differences in the various units of study, clearly differing from existing components and their contribution to the substrate, as well as those elements that have characterized these environments in the past recent.
Key words: Phytolith analysis; Middle Paraná River; Predelta National Park; Entre Ríos province.
Introducción
El río Paraná es el componente más importante
del sistema fluvial del Río de La Plata. El tramo
correspondiente al Paraná medio, se desplaza por
una llanura aluvial (faja inundable compleja y de
gran tamaño, Iriondo, 1988), desde la confluencia
del Paraná-Paraguay hasta el ápice del delta,
localizado algo al sur de Rosario (Paoli & Schreider,
2000). Ésta planicie aluvial, cuyo sistema de canales
de río, lagunas poco profundas, islas y humedales,
sigue siendo básicamente inalterado y desocupado
por el hombre (Iriondo et al., 2007).
Los análisis de fitolitos han mostrado ser una
valiosa contribución para comprender el tipo de
ambiente en que se desarrollaron los depósitos
fluviales recientes, tanto para el estudio de los
depósitos de fondo, como de los ambientes insulares
vinculados (Bertoldi de Pomar, H. 1970, 1971, 1972, 1974, 1975, 1976, 1980 y 1983; Bertoldi
de Pomar, H. &Tur, N., 1970). Los silicofitolitos
son partículas de sílice amorfo hidratado (Si2.n
H2O) que se forman en los espacios intracelulares
o extracelulares de las plantas por el proceso de
biomineralización; luego de la absorción de sílice
en estado soluble de aguas subterráneas (Piperno,
1988), ésta se moviliza a distintos sectores de la
planta en donde se produce su precipitación y
se aloja generalmente por el resto de la vida del
organismo, para luego ser liberado al medio tras
la muerte y descomposición del tejido en donde
se encontraba. Estos cuerpos se incorporan en los
depósitos y de acuerdo al tipo de ambiente pueden
poseer un carácter local o extra local (Zucol et al., 2005).
De esta forma, se hace necesario establecer
la producción fitolítica de las comunidades
actuales, con la finalidad de compararlas con las
asociaciones fitolíticas de depósitos antiguos.
Esta contribución busca definir la composición
fitolítica de las comunidades vegetales de distintos
ambientes insulares de la región más austral del
Paraná medio, mediante el estudio del aporte
vegetal al sustrato y su comparación con los
depósitos recientes, estableciendo características
florísticas y fluviosedimentarias del sistema plantasustrato
de estos ambientes.
Materiales y Métodos
Área de estudio
El área se corresponde con un ecosistema de
llanura de inundación. Este sistema geomorfológico
e hidrológicamente dinámico, presenta ambientes
lóticos y lénticos permanentes, asociados a áreas
de transición (Junk et al., 1989). Los suelos son
de origen principalmente fluvial, de texturas limoarcillosas
o arenosas (de grano medio a fino),
presentando escasa estructuración y desarrollo
en sus perfiles. Si bien existe gran variación, esta
puede sintetizarse en dos gradientes principales: a)
uno del tipo geomorfológico, donde la diferencia
de altura entre albardón y bajo establece la
diferencia entre tipos de suelos (sobre todo la
aparición de suelos hidromorfos), y b) un segundo
tipo vinculado a la energía relativa del agua, que
se interpreta como "distancia al cauce principal"
(sensu Passeggi, 2000).
El patrón hidrológico que presenta esta llanura
de inundación en el Paraná Medio corresponde a un
cauce típicamente anastomosado y se caracteriza por
la presencia de sucesivas divisiones y confluencia de
la corriente (Passeggi, 2000). La llanura aluvial del
mismo, tiene 600 km de longitud desde la confluencia
Paraná-Paraguay hasta el Delta del Paraná (Paoli et
al., 2000). El área de estudio pertenece a la unidad
geomorfológica "llanura de bancos" (Iriondo, 1972),
la cual debe su formación al sucesivo adosamiento de
bancos de cauce, formados por procesos de acreción
lateral del cauce del río Paraná. Este evento determina
la construcción de una sucesión de toposecuencias
naturales, conformadas por albardón-pajonal-bajo,
cuya antigüedad, se incrementa con su distancia al
cauce principal. Posteriormente a su formación, estas
geoformas quedan sujetas a los aportes sedimentarios
producidos por los periódicos eventos de inundación
(depósitos de acreción vertical), en función de la
distancia y la altura con respecto al cauce principal que
determinan, en definitiva, el grado de aislamiento con
el mismo (Paoli et al., 2000).
El área elegida para el este trabajo es el Parque
Nacional Predelta (32º03´43´´S; 60º 38´39´´O) (Fig.
1), el cual comprende una zona de islas en el extremo
norte del Delta del río Paraná, situado a 4 km al
Sur de la ciudad de Diamante (Provincia de Entre
Ríos, Argentina), que comprende un área insular
de 2.458 ha, más un pequeño sector de ambientes
de barranca del río, con variedad de ambientes y
comunidades (Malvarez et al., 1992; Aceñolaza et
al., 1999), los cuales para este caso pueden vincularse
en forma general y de acuerdo a sus características
geomorfológicas como:
Fig.1. Mapa de localización del área de estudio "Parque Nacional Predelta", Localidad de Diamante, Entre
Ríos, Argentina.
Albardón: unidad constituida por un albardón alto
de considerable desarrollo formado por los elementos
fluviales de mayor envergadura. Resultan las áreas más
elevadas del Parque, aunque las evidencias señalan
que las aguas las cubren hasta una altura de más de un
metro durante las máximas crecientes (Malvarez et al., 1992). Dependiendo de la antigüedad de las geoformas
y de la vegetación dominante y característica podemos
encontrar en el área, albardones jóvenes, intermedios
y viejos.
Media Loma - Pajonal: áreas extensas, en donde
los gradientes de reemplazo de los distintos grupos de
especies son amplios y más evidentes que en las otras
unidades (Malvarez et al., 1992). Ocupan los sectores
inmediatos a lo albardones en oposición al cauce
principal y con una zona ecotonal hacia el centro
deprimido del cuerpo insular en donde comparte
especies con el bajo.
Bajo: sector más bajo del cuerpo insular, con la
presencia de agua en superficie en gran parte del año y
vegetación que de acuerdo al desarrollo de este cuerpo
de agua, se puede presentar formando zonificaciones
que responden a la presencia de aguas permanentes,
ocasionales y esporádicas; con un gradiente altitudinal
de escasa altura. En algunos casos, dependiendo del
sector insular, el desarrollo de un bosque pobre en
altura y cobertura, sobre los sectores de mayor altura,
se hacen más dispersos y abundan los ejemplares
muertos cubiertos por enredaderas (Malvarez et al., 1992).
Desde el punto de vista biogeográfico el área
del Parque Nacional Pre-Delta, se encuentra en la
Provincia Paranense (Dominio Amazónico) (sensu Cabrera & Willink, 1973; Cabrera, 1994) con un gran
desarrollo de las comunidades de selvas marginales.
Mientras que para Morrone (2001) el área se ubica
dentro de la Provincia del Bosque Paranaense, que se
distribuye al sudeste del Brasil, noreste de la Argentina
y este de Paraguay.
Las características climáticas del área la ubican
dentro de un clima templado/cálido-húmedo, las
temperaturas medias anuales se encuentran cercanas
a los 19º C. El régimen pluviométrico es de 900 mm,
con precipitaciones que se registran principalmente
en los periodos de octubre a abril (73%). El patrón
hidrológico se corresponde con un régimen pulsátil
anual de inundación, con una época de estiaje (bajante)
que se produce en agosto-septiembre, mientras que el
pulso de inundación suele darse sobre fines del verano
principios del otoño (Aceñolaza et al., 2004).
Del total del área, se establecieron 6 estaciones
denominadas: Albardón viejo, Albardón intermedio,
Albardón joven, Media loma, Pajonal y Bajo. Donde
se realizaron 6 perfiles en total, uno por cada estación,
que fueron muestreados de acuerdo a la siguiente
metodología, se tomó una muestra superficial, es
decir el mantillo u hojarasca, la cual se diferenció
en hojarasca superficial y la muestra de sedimento
de reciente incorporación, esta última obtenida por
medio de la técnica de flotado, seguida de 3 muestras
inferiores cada 5 cm de espesor, hasta llegar a los 15
cm de profundidad, obteniendo de esta manera cuatro
muestras por sitio. Para la extracción de material
silíceo del mantillo, se utilizó la técnica de calcinación
de Labouriau (1983), basada en la incineración de
tejidos vegetales a altas temperaturas dentro de un rango adecuado para reducir el tejido orgánico a
cenizas, dejando así su contenido biomineral.
El sedimento se procesó de acuerdo al protocolo de
procesamiento descripto por Zucol et al (2010), para
ello se tomó entre 15 - 20 gr de muestra y se realizó
su limpieza eliminando los distintos compuestos
(sales solubles, materia orgánica y carbonatos) que
puedan provocar la aglomeración de material silíceo
e imposibilite su correcta observación, la separación
granométrica se realizó por tamizado para el material
grueso y por pipeteado para el material fino, de
esta manera se obtuvieron tres fracciones: gruesa
(> 250 μm); media (53-250 μm) y fina (8-53 μm).
Con las fracciones de 8 a 250 μm, se realizó una
separación densimétrica con politungstato de sodio
como líquido pesado (dens. 2,3). Con el material
flotado se obtuvieron preparados microscópicos que
se realizaron con aceite de inmersión y Bálsamo de
Canadá, como medio de montaje.
Fueron contados de 400 a 500 fitolitos por
muestra usando microscopio Nikon Eclipse E200,
las fotografías fueron tomadas con una cámara digital
Nikon Coolpix S4.
Los fitolitos no articulados fueron definidos usando
una clasificación basada en la propuesta de Patterer et al. (2011), basada en las propuestas de Twiss et
al. (1969), Bertoldi de Pomar (1971), Mulholland,
(1989), Twiss (1992), Kondo et al. (1994), Fredlund
y Tieszen (1994) y Zucol (1996) y siguiendo las
normativas y descriptores propuestos por el ICPNWG
(2005) (Fig.2). Para el resto de material silíceo
por ejemplo, fitolitos articulados, se detallaron los
elementos celulares y/o tisulares que los componían
de acuerdo a Zucol (1995, 1996).

Fig. 2. Clasificación de los morfotipos fitolíticos y acrónimos utilizados para su identificación e ilustración en
las distintas figuras (modificado de Patterer et al., 2011).
Para el caso de biolitos como espículas de
espongiarios, estomatocístes de crisostomatáceas,
diatomeas y otros restos como granos de polen, fueron
observados pero no cuantificados.
Con la información de los recuentos se creó una
matriz básica de datos, obteniendo el porcentaje
de frecuencia relativa de cada morfotipo fitolítico,
realizando con estos valores, gráficos de barras.
Mientras que para los análisis multivariados se
utilizó el programa PAST, PAleontological STatistics (Hammer et al., 2001).
Resultados Albardones
Vegetación característica
Los albardones están dominados por bosques
de curupí (Sapium haematospermum Müll. Arg.),
asociados frecuentemente con seibo (Erythrina cristagalli L.) y esporádicamente timbó (Enterolobium
contortisiliquum (Vell.) Morong), como así también
cardas (Eryngium L.).
Puede observarse algunos sectores de los
albardones caracterizados por la presencia de
timbó blanco o timbosillo (Albizia inundata (Mart.)
Barneby & J.W.Grimes) y ambientes de media
loma con bosques bajos de espinillo (Acacia caven (Molina) Molina). Desde el punto de vista de la
posición topográfica que ocupan y su fisonomía,
estos albardones y medias lomas son ambientes bien
diferenciables entre sí.
Albardón viejo
Asociación fitolítica
Las asociaciones fitolíticas del mantillo u hojarasca
superficial, presenta escaso material silíceo, los fitolitos
son principalmente articulados (Fig. 3 A, B, C), entre
ellos se pudo observar elementos buliformes (flabelos
y poliédricos), pelos silicificados, células epidérmicas
(Fig. 3 A) y subepidérmicas, elementos tipo piezas
de rompecabezas (Fig. 3 S) y aquellos originados en
tejidos de conducción, junto a la abundante presencia
de diatomeas. Dentro de los fitolitos aislados, se
observaron flabelos tipo Fl04, de manera muy
frecuente, así como elongados del tipo Mp09. Dentro
de los fitolitos de menor tamaño se presentaron de
manera muy frecuente elementos bilobados (Fig. 3
D-I), tales como Ha01, Ha02 y Ha07, proteriformes
(Pr01) (Fig. 3 W, X), conos truncados (Ct03) (Fig. 3
J-N) y en forma de bote (Sc01).
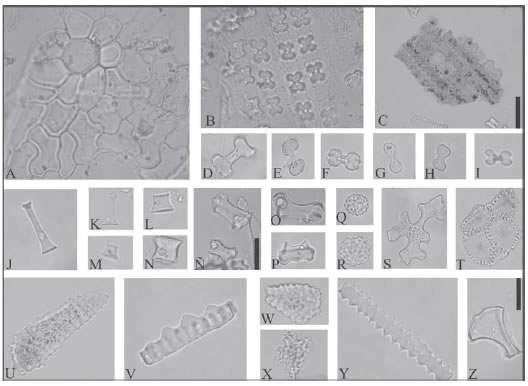
Fig. 3. Fitolitos observados en las muestras. A-C: Fitolitos articulados pertenecientes a muestras de
mantillo. D-Z. Fitolitos pertenecientes a muestras clásticas: D-I: bilobados (Ha). J-N: en forma de cono
truncado (Ct). Ñ-P: fitolitos en forma de silla de montar colapsadas (Sm). Q y R: globulares (Gl). S: fitolitos
de bordes irregulares (Nx). T: fitolito articulado compuesto por elementos cónicos (Pi). U, V y W: fitolitos
fusiformes (Ln). X: fitolito proteriforme (Pr). Y: elemento elongado de borde ondulado (Mp08). Z: fitolito en
forma de flabelo (Fl). Escala gráfica: 20 μm.
En el sedimento, la asociación fitolítica de la
muestra clástica, presentó abundantes fitolitos en
forma de flabelo (Fl03 y Fl04), elongados (Mp09) y
poliédricos (Sx03), entre los fitolitos más pequeños
se encontraron definiendo la asociación, fitolitos
fusiformes (Ln03), en forma de cono truncado (Ct03)
y proteriformes (Pr01).
En lo que respecta a los primeros 10 cm de suelo,
la asociación fitolítica se ve conformada por elementos
fusiformes como Ln03 y proteriformes del tipo Pr01,
muy frecuentes, mientras que los elementos elongados
(Mp08 y Mp09), poliédricos (Sx01, Sx02 y Sx03)
y en forma de flabelos (Fl03 y Fl04), se presentaron
de manera frecuente. En los 5 cm inferiores, se
observaron con mayor frecuencia, elementos en forma
de silla colapsada (Sm06), conos truncados (Ct03)
y globulares (Gl04) (Fig. 3 Q, R) junto a fitolitos
bilobados tipo Ha01, Los fitolitos proteriformes
(Pr01), se observaron con mayor frecuencia.
En lo que respecta a la presencia de biolitos, las
espículas de espongiarios y diatomeas se observaron
de forma abundante, en la totalidad de las muestras.
Albardón intermedio
Asociación fitolítica
En el mantillo, la asociación fitolítica se
caracteriza por la presencia de morfotipos elongados
cortos (Mi01), largos (Mp01 y Mp09), elementos de
conducción biomineralizados (Du02) y en forma de
abanico (Fl04); dentro de los morfotipos de mayor
tamaño. Del grupo de los fitolitos de menor tamaño,
la asociación se caracteriza por la presencia de conos
truncados (Ct03 y Ct05), bilobados tipo Ha07 y
frecuente es la presencia de fitolitos de forma cónica
(Pi01). Se observó la presencia de células epidérmicas
de dicotiledóneas (tipo puzzle), granos de polen y
espículas de espongiarios. En la fracción clástica de
la misma muestra, los fitolitos más frecuentes fueron
Ch01 (en forma de cruz) y Sm01 (en forma de silla
de montar), Ha07 (bilobado), Ct03 (en forma de cono
truncado) y Pr01 (proteriforme).
En los primeros 5 cm, las asociaciones fitolíticas se
caracterizan por la presencia de elementos elongados
(Mp01, Mp08 y Mp09), en forma de flabelo (Fl04),
poliédricos (Mx02 y Mx03) y bilobados (Ha07)
como las formas más abundantes y constantes
dentro de la asociación. Menos frecuentes se
presentaron fitolitos en forma de cruz (Ch01),
conos truncados (Ct03) y globulares (Gl04). En los 10 cm inferiores, la asociación presentó de manera
constante elementos bilobados (Ha01, Ha02 y
Ha07), en forma de cono truncado (Ct03), de
cruz (Ch01), se observaron elementos fusiformes
(Ln03) (Fig. 3 U) abundantes, como así también la
presencia de fitolitos en forma de bote largo (Sc02)
y sillas de montar colapsadas (Sm06) (Fig. 3 Ñ-P).
En todas las muestras se observaron espículas
de espongiarios, fitolitos articulados y pelos
silicificados, así como restos de tejido celular
epidérmico.
Albardón joven
Asociación fitolítica
El mantillo se caracterizó por la presencia casi
exclusiva de elementos articulados tanto de células
cortas como de células largas, principalmente
elementos bilobados, a los cuales acompañan
fitolitos en forma de cruz. En la muestra clástica,
la asociación se caracteriza por la presencia
de fitolitos bilobados (Ha01 y Ha07) y conos
truncados del tipo Ct03. En los primeros 5 cm de
suelo, la asociación fitolitica se mostró sin cambios
significativos con respecto a la muestra superior.
En los restantes 10 cm inferiores del perfil, la
asociación se caracterizó por la presencia de conos
truncados (Ct03), bilobados (Ha01, Ha02 y Ha07) y
fusiformes (Ln03), acompañados de flabelos (Fl04)
elongados (Mp08 y Mp09) y poliédricos (Sx01,
Sx02 y Sx03), los elementos globulares (Gl04) y
en forma de silla colapsada (Sm06) (Fig. 3 Ñ-P) se
presentaron de manera escasa.
Las espículas de esponjas estuvieron presentes
en el total de las muestras analizadas.
Pajonal
Vegetación característica
Las situaciones de interface entre los bosques y
los bajos, se caracterizan por la presencia de la paja
de techar (Panicum prionitis Nees) de gran porte
mientras que en el estrato inferior a este pajonal
se extiende un tapiz vegetal de especies donde se
destacan por su abundancia Eleocharis viridans Kük. ex Osten y Cynodon dactilon (L.) Pers., como
así también la presencia de arbustos como, Sesbania
virgata (Cav.) Pers. y Solanum glaucophyllum Desf.,
los cuales también se extienden hacia bajos anegados.
La presencia llamativa de árboles como el aliso
(Tessaria integrifolia Ruiz & Pav.) y el sauce (Salix
humboldtiana Willd.), está asociada a antiguas
espiras de meandro, que actualmente perduran en el
paisaje pero han sido muy erosionadas.
Asociación fitolítica
El mantillo se caracteriza por la abundancia,
principalmente y de manera constante de elementos
articulados, especialmente células cortas de tipo
bilobadas (Ha01, Ha03 y Ha07) y en forma de cruz
(Ch01), de las formas aisladas si bien de manera
menos frecuente se pudo observar elementos
elongados (Mp01, Mp08 y Mp09), polilobados
(Ph02 y Ph03) y en forma de cono truncado (Ct03).
En la fracción clástica se observaron de manera
muy frecuente fitolitos forma de cruz (Ch01), silla
de montar (Sm01), bilobado (Ha07), cono truncado
(Ct03) y proteriforme (Pr01).
En los primeros 5 cm de suelo, las muestras
presentaron un considerable incremento de fitolitos
aislados y disminución de fitolitos articulados,
principalmente elongados (Mp08 y Mp09), flabelos
(Fl04) y poliédricos (Mx03), de los elementos
pequeños, se observaron frecuentemente fitolitos
bilobados (Ha07), menos abundantes se presentaron
elementos proteriformes (Pr01), en forma de silla
de montar colapsada (Sm06), polilobados (Ph02) y
fusiformes (Ln03).
En los restantes 10 cm de profundidad se observó
un incremento en los morfotipos de mayor tamaño,
como la presencia de elongados (Mp09), flabelos
(Fl04) y poliédricos Sx01, Sx02 y Sx03) asociados a
una mayor presencia de elementos en forma de cono
truncado (Ct03) y bilobados (Ha01 y Ha07).
Se observaron espículas de espongiarios,
diatomeas y granos de polen de manera constante.
Media Loma
Asociación fitolítica
En este perfil, el mantillo se caracterizó por la
abundancia de fitolitos articulados, principalmente
de células cortas, entre ellos elementos articulados
de forma tipo-puzzle, junto a fitolitos en forma
de sombrero articulados (Fig. 3 T), de manera
menos frecuente se observaron elementos bilobados
aislados. La muestra clástica presentó de manera
frecuente fitolitos en forma de flabelo (Fl03 y
Fl04), elongados (Mp08 y Mp09), poliédricos
(Sx01 y Sx02) entre los de mayor tamaño. Entre
los de menor tamaño se observaron de forma
frecuente, globulares (Gl04), fusiformes (Ln03),
cónicos (Pi02), proteriformes (Pr01) y tipo silla de montar colapsada (Sm06). Los primeros 5 cm de
suelo, presentaron una asociación que contó con la
presencia de manera frecuente de bilobados (Ha01y
Ha14) y conos truncados (Ct03).
En los 10 cm inferiores del perfil, se encuentran
definiendo la asociación, los fitolitos bilobados
(Ha14), así como fitolitos en forma de cono
truncado (Ct03), cónicos o en forma de sombrero
(Pi02) (Fig. 3 T), fusiformes (Ln03), proteriformes
(Pr01) y fitolitos en forma de silla de montar
colapsada (Sm06).
Elementos silicios como estomatocistes de
crisotomatáceas y espículas de esponjas, se observaron
de manera abundante en todas las muestras.
Bajo
Vegetación característica
En el área más baja de la toposecuencia
analizada, se desarrolla un terreno saturado con
agua casi en forma permanente y se caracteriza
por la presencia de un tapiz vegetal continuo, con
parches de especies que alternan su dominancia
como Cynodon dactilon, Eleocharis viridans, Enydra anagallis Gardner, Alternanthera
philoxeroides (Mart.) Griseb., Paspalum distichum L. e Hydrocotyle modesta Cham. & Schltdl. entre
otras (Malvarez et al., 1992).
Asociación fitolítica
En la muestra correspondiente al mantillo,
los fitolitos más abundantes son principalmente
articulados originados a partir de células cortas
como de células largas, entre ellos los más
abundantes fueron aquellos en forma de cono
truncado (Ct03 y Ct05) o bilobados de base cónica
(Ha14) y polilobados (Ph02 y Ph03). En la muestra clástica se observó de manera frecuente elementos
globulares (Gl04), en forma de cono truncado
(Ct03), en forma de silla de montar colapsada
(Sm06). Dentro de los fitolitos de mayor tamaño, se
presentaron fitolitos fusiformes (Ln01), elongados
(Mp08 (Fig. 3 I) y Mp09) y poliédricos (Sx01).
En las muestras de suelo, los primeros 10 cm,
se caracterizaron por la presencia de elementos
globulares (Gl04), fusiformes (Ln03), en forma de
cono truncado (Ct03) y en forma de silla colapsada
(Sm06), también presentes en la asociación, de
forma frecuente, elongados (Mp09), en forma de
flabelos (Fl04) (Fig. 3 G) y poliédricos (Sx01 y
Sx02). Los últimos 5 cm del perfil se caracterizaron
por la presencia de elementos bilobados (Ha01
y Ha07), globulares (Gl04), fusiformes (Ln03
y Ln04) y en forma de cono truncado (Ct03),
acompañan la asociación elementos elongados
(Mp08) y poliédricos (Sx01 y Sx03).
Elementos como diatomeas de diversas formas y
tamaños y espículas de espongiarios, se presentaron
frecuentemente.
Consideraciones finales
En términos generales, del análisis de las asociaciones fitolíticas puede considerarse que, el material silíceo se presentó de forma muy abundante y de manera diversa. La frecuencia con la que se presentaron los diferentes morfotipos (Fig. 4) varía en el mantillo con respecto a las muestras de suelo, por un lado la composición de la hojarasca presenta un mayor porcentaje de elementos articulados, los cuales disminuyen a medida que se profundiza en los perfiles. En cuanto a su afinidad botánica, los elementos panicoides (Fig. 3 D-I) fueron en todos los casos los más importantes, seguido por elementos de afinidad danthonioides (Fig. 3 J-N), menos frecuentes fueron los elementos chloridoides, sumados a elementos en forma de silla colapsada (Fig. 2 Ñ-P) (Concave dum-bells) de afinidad bambusoide (Montti et al., 2009), presentes principalmente en la zona de media loma, menos frecuente en pajonal, albardón joven y bajo. Aquellos de afinidad ciperoides (Fig. 3 T) estuvieron presentes en la zona de albardones, así como los fitolitos de dicotiledóneas (Fig. 3 S).

Fig. 4. Esquema de las unidades fisionómicas estudiadas y la frecuencia relativa de los diferentes fitolitos,
agrupados de acuerdo a su afinidad botánica. Barra negra (frecuencia relativa de las muestras del mantillo)
barra gris (frecuencia relativa de las muestras de suelo).
Del análisis multivariado realizado a la totalidad de las muestras, se pudo observar (Fig. 5) una clara diferencia entre las asociaciones fitolíticas de mantillo versus las asociaciones fitolíticas de suelo. De esta forma el análisis presenta dos agrupamiento bien definidos, por un lado las muestras de mantillo de los albardones intermedio y viejo, con las muestras de suelos de todos los perfiles, lo cual se podría entender como resultado de la madurez de estas unidades fisionómicas, cuya composición fitolítica, no se vería alterada de manera frecuente por la acción de las aguas de crecidas, siendo de esta forma más estables frente al resto de las unidades.

Fig. 5. Representación del análisis de correspondencia con respecto a todas las muestras estudiadas. Los
círculos representan las muestras de suelo y los cuadrados las muestras de mantillo.
En los mantillos del albardón joven, media loma, pajonal y bajo, los elementos constituyentes, tendrían más que ver con una composición vegetal actual que caracteriza estas unidades, siendo los morfotipos articulados responsables de la separación de estas muestras frente al resto.
Discusión
En lo que respecta a la transecta de estudio,
la génesis y la evolución geomorfológica que
ha definido las características sedimentológicas
de los materiales, desde su formación; y que
conforman los depósitos sedimentarios de los
ambientes estudiados, tenemos, por un lado, la
influencia de la dinámica hidrosedimentológica del
río Paraná a través del progresivo aumento, en la
proporción de materiales finos, desde los perfiles
cercanos a la fuente de aporte (los cursos de agua
vinculados al cauce principal), hacia el interior de
la llanura de bancos y la pérdida de selección de
los depósitos en la misma dirección. Por otro lado,
estos materiales han sido sometidos a un proceso de
selección en un ambiente de alta energía para luego
ser transportados, sin que su tamaño característico
sea modificado esencialmente, a otro ambiente de
menor energía efectiva de selección, donde sufrieron
un proceso de mezcla conformando una matriz
pobremente seleccionada (Passeggi, 2000). Desde el
punto de vista pedológico, la naturaleza de la fuente
de procedencia de los materiales sedimentarios,
manifiesta su influencia sobre las características
edáficas de los suelos jóvenes asociados a estos
depósitos. Los mismos revelan la ausencia de
horizontes genéticos naturales o la débil expresión
de horizontes incipientes. Dicha composición, no
cumple los requisitos de ninguno de los horizontes
de carácter diagnóstico.
La presente contribución, es el primer análisis de
fitolitos para el área de estudio en lo referente a los
distintos ambientes característicos de una llanura de
inundación. Se cuenta solo con un antecedente en este
tipo de trabajos, obtenidos en sitios arqueológicos
para la presente área de estudio (Bonomo et al., 2011). Los resultados pueden compararse a lo largo
de la sección vertical de cada uno de los perfiles o
bien en sus muestras correspondientes al mantillo, de
este modo, abarcar desde los biomorfos degradados
e incorporados al sustrato, como así también los existentes en restos orgánicos no incorporados al
momento de la toma de muestra.
En el mantillo y de manera casi constante,
las asociaciones fitolíticas se caracterizaron por
presentar elementos articulados principalmente. En
la zona del albardón joven y pajonal, se observaron en
general: fitolitos articulados de células cortas de tipo
bilobados, en forma de cruz o tipo conos truncados.
En el mantillo correspondiente a la zona de la media
loma, se presentaron de manera frecuente elementos
de afinidad chloridoide y en el albardón viejo, las
muestras se caracterizaron por presentar elementos
articulados, pero en este caso, células epidérmicas
o subepidérmicas, pelos silicificados, elementos de
conducción y fitolitos cónicos afines a ciperáceas.
En lo que respecta a las muestras correspondientes
a los niveles de suelo, las mismas se caracterizaron
por diferenciarse claramente de las muestras
correspondientes al mantillo. A su vez en la mayoría
de los perfiles, los primeros centímetros mostraron
diferencias en cuanto a la composición fitolítica con
respecto a la última sección del mismo perfil. Sin
embargo, todas las muestras analizadas presentaron
abundantes microrestos de naturaleza silícea como
así también granos de polen y esporas. Se observaron
abundantes espículas de espongiarios y diatomeas
en todas las muestras, siendo bastantes raros los
estomatocistes de crisostomatáceas.
En cuanto al estado de conservación de los
microrestos, éstos, presentaban un marcado grado
de desgaste, principalmente los fitolitos de mayor
tamaño, mientras que aquellos de menor tamaño
presentaron un bajo grado de desgaste. El análisis
comparativo de la variabilidad y afinidad botánica
de los distintos morfotipos fitolíticos hallados,
mostró una marcada diferencia tanto en abundancia
relativa como en morfotipos característicos de
distintos grupos vegetales, siendo siempre la
familia de las gramíneas, la mejor representada,
con una clara presencia de componentes panicoides,
principalmente en el mantillo, sobretodo en el
correspondiente a la zona del pajonal, donde se
encontraría de manera abundante, especies como Panicum prionitis. Los morfotipos de afinidad
danthonioides, tuvieron una presencia constante
en todo el perfil y los componentes de ciperáceas,
fueron abundantes en las zonas correspondientes a
los albardones. En lo que respecta a la presencia de
elementos característicos de dicotiledóneas como
elementos tipo puzzle, elementos de conducción
y pelos silicificados con su superficie espinosa, se
observaron de manera abundante especialmente en la
zona de albardones (principalmente albardón viejo) y
la zona de media loma, lo que estaría dado por una
vegetación más bien del tipo herbácea y arbórea. Los
morfotipos afines a la familia de las podostemáceas,
se encontraron de manera abundante en todos los
perfiles analizados, principalmente en los niveles más
profundos. Los mismos se encontraban inalterados
en su forma, por lo que puede conjeturarse que o bien
por el escaso transporte por sus forma redondeadas
y el fuerte grado de silicificación, estos cuerpos no
muestran el mismo deterioro ante los agentes de
transportes fluviales, ya que debe tenerse en cuenta
que estas plantas solo se las puede encontrar en aguas
turbulentas y en la actualidad se encuentran presentes
en el NEA hasta los trópicos.
Conclusiones
El análisis fitolítico resultó ser una herramienta útil,
ya que ha permitido marcar diferencias significativas
en las diferentes unidades de estudio. En la mayoría
de los casos, se observó una zonificación marcada,
donde se separan por un lado, las muestras del
mantillo con una composición de materia orgánica
actual o parcialmente degradada, evidenciada en el
tipo de afinidad que presentaron los morfotipos, la
cual coincide con la cobertura vegetal actual, y por
otro lado las muestras de suelo con una composición
bastante característica. En general, el análisis del
mantillo brinda información acerca de la cobertura
vegetal de las zonas estudiadas y la incorporación
inmediata al sustrato.
La presencia de fitolitos que caracterizan a
familias como las podotemáceas, plantas que no se
encuentran formando parte de la cobertura vegetal
actual, es de importancia en el registro obtenido, ya
que su presencia, estaría indicando la consecuencia
de un reemplazo vegetacional, ocurrido como
resultado de cambios de estos paisajes en tiempos
pasados recientes.
Agradecimientos
A todas las personas que dirigen y coordinan el área del Parque Nacional Predelta, Diamante, Entre Ríos. Este trabajo se realizó en el marco del proyecto PIDP - UADER 2003 Res. 553/05.
Bibliografía
1. ACEÑOLAZA, P., J. DE DIOS. MUÑOZ & R. ZANELLO. 1999. Flora y vegetación del Parque Nacional Pre Delta. Reuniones de Comunicaciones de la Asociación Biológica del Litoral. Santa Fé.
2. ACEÑOLAZA, P., H. POVEDANO, A. MANZANO, J. DE DIOS. MUÑOZ, J. ARETA & A. L. RONCHI VIRGOLINI. 2004. Biodiversidad del Parque Nacional Pre-Delta. En: Temas de la Biodiversidad del Litoral Fluvial Argentino, Aceñolaza, F.G. (ed.). INSUGEO Misceláneas 12: 169-184 p.
3. BERTOLDI DE POMAR, H. 1971. Ensayo de clasificación morfológica de los silicofitolitos. Ameghiniana 8: 317-328.
4. BERTOLDI DE POMAR, H. 1976. Métodos de preparación de sedimentos clásticos para su estudio microscópico. I. Tratamientos previos. Asoc. Cs. Nat. Lit. 7: 1-55.
5. BERTOLDI DE POMAR, H. 1970. Fitolitos y Zoolitos. Su significado geológico en sedimentos continentales. Bol. Asoc. Geol. Cba. 1: 21-31.
6. BERTOLDI DE POMAR, H. 1972. Ópalo Organógeno en Sedimentos Superficiales de la Llanura Santafesina. Ameghiniana 9: 265-279.
7. BERTOLDI DE POMAR, H. 1975. Los silicofitolitos: Sinopsis de su conocimiento: Darwiniana 19: 173- 206.
8. BERTOLDI DE POMAR, H. & N. M. TUR. 1970. Células silicificadas en gramíneas acuáticas. Revista de la Asociación de Ciencias Naturales del Litoral 1:17-18.
9. BONOMO, M., M. M. COLOBIG, E. PASSEGGI, A. F. ZUCOL & M. BREA. 2011. Multidisciplinary studies at Cerro Tapera Vázquez site, Pre-Delta National Park, Argentina: The archaeological, sedimentological and paleobotanical evidence. Quaternary International 245 48-61.
10. CABRERA, A. L. 1994. Regiones Fitogeográficas Argentinas: Encicl. Arg. Agr.y.Jar. Fascículo 1, Editorial ACME S.A.C.I., Buenos Aires, 298 pp.
11. CABRERA, A. L. & A. WILLINK. 1980. Biogeografía de América latina. Monografía de la Secretaria General de la Organización de los Estados Americanos. Serie biología Nº 13. Washington, D.C. 123 p.
12. FREDLUND, G. G. & L. T. TIESZEN. 1994, Modern phytolith assemblages from the North American Great Plains: Journal Biogeography 21, 321-335.
13. HAMMER, Ø, D.A.T. HARPER & P. D. RYAN. 2001. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeont. Elec.4(1), 9 pp.
14. IRIONDO, M. H. 1972. Mapa geomorfológico de la llanura aluvial del Río Paraná desde Helvecia hasta San Nicolás, República Argentina. Rev. Asoc. Geol. Arg. 27: 155-160.
15. IRIONDO, M., M. J. PARMA & J. C. PAGGI. 2007. The Middle Paraná River Limnology of a Subtropical Wetland. Springer-Verlag, Berlin Heidelberg, 382 pp.
16. IRIONDO, M.H. 1988. A comparision between the Amazon and Paraná River Systems, In: Transport of carbon and minerals in major world rivers, lakes and estuaries (Degens, E. et al., Eds.), SCOPE/UNEP, Hamburg, Part 5, 77-92 pp.
17. JUNK, W. J., P. BAYLEY & R. E SPARKS. 1989. The flood pulse concept in river floodplain systems. En: Proc. of the Internat. Large River. Symp. Canad. Spec. Publ. Fish Aquatic. Sci. Dodge, D.P. (Ed.). 101-127 pp.
18. KONDO, R., C. CHILD & I. ATKINSON. 1994. Opal phytoliths of New Zealand: Maanaki Whenua Press, 85 pp.
19. LABOURIAU, L.G. 1983. Phytolith work in Brazil: a minireview: The Phytol. News 2: 6-10.
20. MADELLA, M., A. ALLEXANDRE & T. BALL. 2005. International Code for Phytolith Nomenclature 1.0.: Annals of Botany 96(2), 253-260.
21. MALVAREZ, A., P. KANDUS & J. A. MERLER. 1992. Evaluación y diagnóstico del Parque Nacional "Predelta La Azotea" Inf. Ined. 22 pp. UBA-APN.
22. MONTTI, L., M. FERNÁNDEZ HONAINE, M. OSTERRIETH & D., GRACIANO RIBEIRO. 2009. Phytolith analysis of Chusquea ramosissima Lindm. (Poaceae: Bambusoideae) and associated soils. Quaternary International 193:80-89.
23. MORRONE, J. J. 2001. Biogeografía de América Latina y el Caribe. M and T- Manuales y Tesis SEA, vol. 3. 25-29 pp.
24. MULHOLLAND, S.C. 1989. Phytolith shape frequencies in North Dakota grasses: a comparison to general patterns: Journal Archaeology Science 16, 489-511.
25. PAOLI, C., M. IRIONDO & N. GARCIA. 2000. Características de las cuencas de aporte. En: El río Paraná en su tramo medio. Contribución al conocimiento y prácticas ingenieriles en un gran rio de llanura. Paoli, C. y Schreider, M. (eds.), Centro de publicaciones Universidad Nacional del Litoral Tomo 1: 27-69 p.
26. PAOLI, C. & M., SCHREIDER.2000. El río Paraná en su tramo medio "Contribución al conocimiento y prácticas ingenieriles en un gran río de llanura", Centro de Publicaciones, Secretaria de Extención, UNL (Universidad Nacional del Litoral), Santa Fé, Argentina, Tomo 1, 309 pp.
27. PASSEGGI, E., 2000, Caracterización sedimentológica del material parental de los suelos asociados a los depósitos del cauce del Tramo medio del río Paraná, Tesis Doctoral. Universidad Católica de Santa Fé, 216 pp.
28. PATTERER, N. I., E. PASSEGGI & A. F. ZUCOL. 2011. Análisis fitolíticos de suelos del sudoeste de la Provincia de Entre Ríos (Argentina) como una herramienta para comprender sus procesos pedológicos. Rev. Mex. Cs. Geol., v. 28, núm. 1, 2011, 132-146 p.
29. PIPERNO, D. R. 1988. Phytolith analysis: an archaeological and geological perspective. San Diego. Academic Press 280 pp.
30. TWISS, P. C. 1992. Predicted world distribution of C3 and C4 grass phytoliths, en Rapp, G. Jr , Mulholland, S.C. (eds.), Phytoliths Systematics. Emerging Issues: Advances in Archaeological and Museum Science 1, 113-128.
31. TWISS, P. C., E. SUESS & R. M. SMITH. 1969. Morphological classification of grass phytoliths: Soil Science of America Proceeding 33,109-115.
32. ZUCOL, A. F. 1995. Microfitolitos: II Análisis de las clasificaciones. Ameghiniana 32: 243-248.
33. ZUCOL, A. F. 1996. Microfitolitos de las Poaceae argentinas: I. Microfitolitos foliares de algunas especies del género Stipa (Stipeae: Arundinoideae), de la Provincia de Entre Ríos. Darwiniana 34: 151- 172.
34. ZUCOL, A. F., M. BREA & A. SCOPEL. 2005. First record of fossil wood and phytolith assemblages of the Late Pleistocene in El Palmar National Park (Argentina). Journal of South American Earth Sciences 20: 33-43.
35. ZUCOL, A. F., E., PASSEGGI, M., BREA, N. I., PATTERER, MA. G., FERNÁNDEZ PEPI & M. M., COLOBIG. 2010 b. Phytolith analysis for the patrok aike lake drilling proyect: sample treatment protocols for the pasado microfossil manual". En: Corbella, H. & Maidana, N. I. (Eds.). 1ª Reunión Internodos del Proyecto Interdisciplinario Patagonia Austral y 1er Workshop Argentino del Proyecto Potrok Aike Maar Lake Sediment Archive Drilling Project. Proyecto Editorial PIPA. Buenos Aires, Argentina. Pp 81-84.
Recibido el 3 de febrero de 2012,
aceptado el 31 de
octubre de 2012.