ARTÍCULOS
Procesos de intensificación y análisis tafonómicos: el
caso del sitio Laguna El Sosneado 3 (LS-3) (Mendoza,
Argentina)
Clara Otaola, Miguel A. Giardina y Fernando J. Fernández
Clara Otaola. Consejo Nacional de Investigaciones Científicas y Técnicas. Instituto Argentino de Nivología, Glaciología y
Ciencias Ambientales (CONICET-IANIGLA). Museo de Historia Natural de San Rafael. Av. Ballofet s/n, Parque Mariano
Moreno (5600), San Rafael, Mendoza, Argentina.
E-mail: claraotaola@yahoo.com.ar
Miguel A. Giardina. CONICET- IANIGLA. Museo de Historia Natural de San Rafael. Av. Ballofet s/n, Parque Mariano
Moreno (5600), San Rafael, Mendoza, Argentina.
E-mail: mgiardina@mendoza-conicet.gob.ar
Fernando J. Fernández. CONICET. Universidad Nacional de la Plata. Laboratorio de Anatomía Comparada y Cátedra de
Zooarqueología. Calle 64 s/n e/113 y 120 (1900), La Plata, Buenos Aires, Argentina.
E-mail: fernandofernandez@yahoo.com
Recibido 13 de mayo 2013.
Aceptado 27 de agosto 2013
RESUMEN
En este artículo presentamos los resultados del análisis del sitio arqueológico LS-3, localizado en la porción
cordillerana del sur de Mendoza. Este posee una antigüedad máxima de 2000 años AP, por lo tanto es contemporáneo
a momentos en los que se ha registrado un proceso de intensificación. Para entonces se observa
la incorporación de nuevos espacios a los circuitos de movilidad y un mayor espectro de recursos en la dieta,
que incluye fauna de pequeño tamaño y bajo rendimiento económico. Sin embargo, los análisis tafonómicos
realizados en los conjuntos recuperados en este sitio señalan que los humanos no fueron los principales agentes
responsables del ingreso de la fauna a LS-3. La fauna pequeña y mediana no presenta indicios de explotación
humana, y la fauna de tamaño grande no muestra evidencias de procesamiento intensivo. Consideramos que el
análisis tafonómico exhaustivo en sitios como este es fundamental para evaluar si son factibles de ser utilizados
para analizar el proceso de intensificación planteado para la región.
Palabras clave: Proceso de intensificación; Evidencias arqueofaunísticas; Análisis tafonómicos; Sur de Mendoza.
ABSTRACT
Intensification processes and taphonomic analyses: the case of laguna el Sosneado
3 site (ls-3). In this article the results of the analyses carried out at archaeological site LS-3 are presented.
This site is located in the Andean portion of southern Mendoza and has an earliest date of 2000 years BP. It is
contemporary with an intensification process proposed for the region, which implied the incorporation of new
areas within mobility systems and the broadening of the diet to include low ranked fauna. Taphonomic studies,
however, note that humans were not the main agents responsible for bringing the specimens to the site. Small
and medium fauna do not show evidence of human butchering, and large fauna does not show evidence of
intensive butchering. It is argued that comprehensive taphonomic analysis on sites like this is essential to assess
whether it is feasible to use such materials to study the process of intensification proposed for the region.
Keywords: Intensification Process; Archaeofaunal record; Taphonomic analyses; Southern Mendoza.
INTRODUCCIÓN
El sitio arqueológico Laguna El Sosneado 3 (LS-3)
se encuentra en la región cordillerana del sur de la
provincia de Mendoza. Está ubicado en las proximidades
de la laguna homónima, en la cuenca superior
del río Atuel, a 2100 msnm (34°51' S, 69°53'
O; Figura 1 a y b). En la región del sur mendocino,
el proceso de intensificación de los recursos ha sido
evaluado a partir de la teoría de forrajeamiento óptimo
(Pianka 1983; Stephens y Krebs 1986), utilizando
modelos como el de amplitud de la dieta (Bettinger
1991; Nagaoka 2002). En este contexto, la ampliación
de los rangos de acción, así como de la diversidad
de los recursos consumidos, habría sido una de las
consecuencias observables de dicho proceso (Neme
2007; Neme y Gil 2008; Giardina 2010; Llano 2013).
Siguiendo este modelo, se espera en nuestra región de
estudio evidencias de consumo de fauna de bajo rendimiento
económico, como los vertebrados de tamaño
mediano y pequeño a partir de la segunda mitad del
Holoceno tardío.
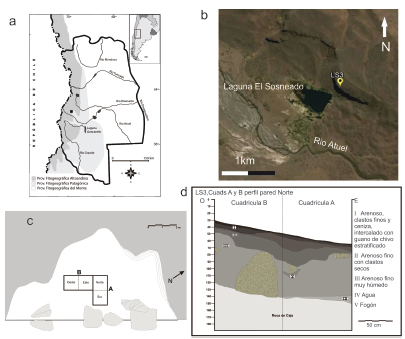
Figura 1. a: Mapa fitogeográfico (sensu Cabrera 1976) de la provincia de Mendoza con la
ubicación del sitio arqueológico LS-3; b: imagen satelital del Google Earth que muestra la
localización del sitio y los principales accidentes topográficos del lugar; c: planta del sitio
con la disposición de las cuadrículas A y B; d: esquema estratigráfico del perfil norte de
las cuadriculas A y B.
Los objetivos del presente trabajo radican en analizar
las características del conjunto arqueofaunístico del
sitio LS-3, atendiendo especialmente a los agentes que
intervinieron en su conformación. Por otro lado, en
función de los antecedentes mencionados, se espera
que el conjunto arqueofaunístico de este sitio muestre
evidencias concordantes con un consumo antrópico
de animales de tamaño
mediano y pequeño. Para
discutir esta expectativa,
analizamos aquí los agentes
responsables del origen
de vertebrados pequeños,
medianos y grandes y su
potencial uso por las poblaciones
humanas.
El sitio arqueológico
Laguna El Sosneado 3
El sitio LS-3 se encuentra
en un área ecotonal
entre las provincias
fitogeográficas Patagónica
y Altoandina (Cabrera
1976). El área se caracteriza
por tener un clima
frío y seco, con precipitaciones
invernales de tipo
nival (600 a 1000 mm
anuales) y la vegetación
dominante es la estepa
arbustiva (Cabrera 1976)
(Figura 1a). La cueva LS-3 se encuentra a unos 40 m sobre el
nivel de la laguna, en un farallón basáltico en el que
hay otras cuatro cuevas y aleros separados entre 100
y 60 m entre sí. LS-3 posee una superficie aproximada
de 45 m2. La apertura de la cueva es de 12 m y la
distancia máxima entre la línea de goteo y la pared
interior es de 6 m (Figura 1 c). Los primeros trabajos
en el sitio fueron realizados en el año 1992, cuando
se realizaron dos sondeos de 50 x 50 cm cada uno. En
uno de ellos se alcanzaron los 60 cm de profundidad
sin evidencias de restos arqueológicos. En el segundo
sondeo se hallaron restos de microvertebrados, fragmentos
óseos indeterminados y un artefacto lítico en
la Unidad 2 (Neme 2007) que, junto con un fragmento
de hueso largo con fractura longitudinal hallado en la
misma unidad, constituyen las únicas evidencias claras
de actividad antrópica en este sitio. En el año 2000 se
amplió la excavación a una superficie de 3 m2. Esta
fue realizada siguiendo niveles artificiales de 5 cm
de espesor cada uno, y se alcanzó una profundidad
máxima de 1,05 m.
Se describió la estratigrafía sedimentaria observando
el perfil de las cuadrículas excavadas (Figura
1 d). La unidad estratigráfica I está constituida por
sedimentos arenosos con clastos finos, intercalados
con ceniza y guano de Capra hircus. En la cuadrícula
B esta unidad posee un espesor de 10 cm, mientras
que en la cuadrícula A es de 5 cm. La unidad estratigráfica
II posee un espesor irregular pero de unos 10 cm en casi toda la cuadrícula B y llega a un espesor
de 20 cm en partes de la cuadrícula A. Está formado
por sedimentos de arena fina intercalada con clastos
de arcilla seca. La unidad III tiene 70 cm de espesor y
está formada por sedimentos arenosos, finos, y posee
mucha humedad. En particular, este sitio se caracteriza
por tener un alto contenido de humedad en su
matriz sedimentaria. A partir del nivel de excavación
4, el sedimento se presenta muy húmedo y arcilloso,
y en el nivel 5 aparecieron rocas en un estado avanzado
de descomposición. A partir del nivel 12 de la
cuadrícula A, los sedimentos son tan húmedos que
fue necesario cernirlos con agua. Para el análisis y
comparación diacrónica de los materiales, la secuencia
fue dividida en cuatro unidades de análisis de espesor
equivalente (Tabla 1).
Tabla 1. Unidades de análisis y sus fechados radiocarbónicos.

MATERIALES Y MÉTODOS
El análisis de los materiales fue desarrollado de
acuerdo con las metodologías particulares de cada grupo
taxonómico (aves, reptiles y mamíferos). En cuanto
a los mamíferos y las aves, se realizaron subcategorías
de acuerdo con el tamaño. Se incluyen dentro de la
categoría micromamíferos a aquellas especies con un
peso menor a 1 kg, tales como marsupiales, roedores
cávidos y cricétidos. Los mamíferos medianos considerados
aquí comprenden a aquellos taxones con un
peso de entre 1 y 15 kg, tales como dasipódidos, los
roedores mayores a 1 kg, los cánidos y los mustélidos.
Por último, la referencia a mamíferos grandes remite
a aquellos que poseen un peso mayor a 15 kg. Con
respecto a las aves; consideramos aves pequeñas a
columbiformes y paseriformes, y aves medianas a los
anátidos, falconiformes y estrigiformes.
Para el análisis de reptiles y micromamíferos se
siguió la metodología y clasificación propuesta por
Andrews (1990), Fernández-Jalvo y Andrews (1992)
y Castillo et al. (2001). En este marco, se observaron
las huellas de corrosión producidas por la acción de
ácidos digestivos sobre las superficies de los restos
dentarios y poscraneales (i.e., epífisis proximal de fémur
y epífisis distal de húmero), los cuales se caracterizan
por generar áreas de reducción del esmalte
dentario y porosidades en los extremos de los huesos.
Se analizó el grado de fractura de los restos craneales y
poscraneales, y se contabilizaron los restos completos
y las diferentes porciones (i.e., diáfisis, epífisis proximal
y epífisis distal) (Andrews 1990; Fernández-Jalvo y Andrews 1992; Castillo et al. 2001). Se evaluó la
abundancia relativa de los elementos esqueléticos y se
consideró la representatividad de cada uno de ellos en
el marco del número mínimo de individuos: NMEi / (Ei
x NMI) x 100, donde NMEi es el número mínimo de
elementos esqueléticos particulares en la muestra y Ei es
el número esperado de ese elemento esquelético en un
individuo (Andrews 1990; Fernández-Jalvo y Andrews
1992). Por otro parte, se observaron los efectos de la
exposición de los restos óseos y dentarios a los agentes
meteóricos (i.e., la radiación ultravioleta [UV] del
sol, el viento, la lluvia y los cambios de temperatura),
siguiendo la propuesta de estadios de meteorización
de Andrews (1990) para los restos de micromamíferos.
También se evaluaron otras variables posdepositacionales
tales como pisoteo, marcas de raíces, impresiones de
óxido de manganeso, abrasión por transporte hídrico y
corrosión sedimentaria (cf. Korth 1979; Andrews 1990;
Fernández-Jalvo y Andrews 2003).
Para el estudio de los conjuntos óseos de aves
pequeñas se siguió la metodología desarrollada por
Ericson (1987), Bocheński et al. (1998), Livingston
(1989) y Bocheński y Tornberg (2003). En primer lugar,
se buscó segregar aquellos conjuntos que provienen de
actividad humana de aquellos que han sido depositados
por agentes naturales mediante el estudio de egagrópilas,
del mismo modo que con los otros microvertebrados.
En tal sentido, se evaluó la corrosión producida
por los ácidos digestivos siguiendo la metodología
propuesta por Bocheński y Tomek (1997). Además, se
siguió la propuesta de Bocheński y Tornberg (2003), en
relación con restos no ingeridos por aves diurnas, y se
observaron el grado de fragmentación de los huesos,
la relación de elementos del miembro anterior con el
posterior, la relación de elementos proximales y distales,
y la relación de elementos del esqueleto axial y
apendicular. Los datos obtenidos para aves pequeñas
de LS-3 son comparados con el modelo de Bocheński y
Tornberg (2003), para testear si el registro avifaunístico
es producto de la acción depredadora de aves rapaces.
Asimismo, se compara la curva de la representación de
elementos anatómicos y el patrón de alteración de los
ácidos gástricos en huesos, generados por dos especies
de aves Strigiformes (Tyto alba [lechuza de campanario]
y Bubo virginianus magellanicus [nuco]), que habitan en
la región de estudio (cf. Fernández et al. 2009).
La identificación taxonómica, anatómica y la cuantificación
de los restos de mamíferos de tamaño
mediano y grande se realizaron siguiendo los criterios
desarrollados por Lyman (1994,
2008) y Mengoni Goñalons (1999).
Las variables analizadas para evaluar
el perfil tafonómico para este grupo
de vertebrados fueron la meteorización
(sensu Behrensmeyer, 1978); la proporción
de especímenes no identificados, modificaciones mecánicas (i.e., pisoteo, abrasión, marcas
de roedores, daños producidos por carnívoros),
modificaciones químicas (i.e., presencia de manganeso,
termoalteración, teñido por causas indeterminadas,
daños generados por raíces), tipo de fracturas de los
huesos; estado del hueso al momento de fracturarse
(i.e., en estado seco o fresco) y longitud máxima de
los huesos (Outram 2002).
Tanto las modificaciones
mecánicas como las químicas
fueron clasificadas según
la morfología del daño por
analogía con imágenes publicadas
en la bibliografía
consultada (Haynes 1980;
Binford 1981; Martin 1998;
Fernández et al. 2010) y por
observaciones actualísticas
propias. Además se consultaron
muestras actuales del
Instituto Multidisciplinario
de Historia y Ciencias
Humanas, CONICET. Esta
colección contiene material
óseo con punctures, pits y scoring (cf. Binford 1981)
generadas por Lycalopex
culpaeus, Puma concolor y Conepatus chinga (ver
Martin 1998; Mondini 2003).
Finalmente, para evaluar
el origen antrópico de los
distintos taxa se consideró
la presencia de huellas de
corte en los especímenes y
la presencia de termoalteración
en zonas diagnósticas
identificadas como resultado
de la cocción de las presas
(Pardiñas 1999; Medina et
al. 2012). En cuanto a los
conjuntos óseos de camélidos,
para definir su carácter
antrópico, se siguieron
los criterios definidos
por L´Heureaux y Borrero
(2002), tales como la presencia
de modificaciones
culturales en los huesos,
asociación de los elementos
óseos con artefactos, la
proporción diferencial de
huesos del esqueleto axial/
apendicular y proporción
diferencial entre elementos
enteros/fragmentos.
RESULTADOS
Como se ilustra en la Tabla 2, los conjuntos arqueofaunísticos
están principalmente conformados
por micromamíferos (88%), seguidos en orden de
importancia por los mamíferos de tamaño mediano
y grande (6,3%), los reptiles (4,1%) y por último las
aves (1,4%).
Tabla 2. Representación taxonómica de los especímenes hallados en el sitio LS-3.

Referencias: *Sólo elementos poscraneales.
Reptiles
En el sitio se registra un NISP de 261 reptiles, representados
por saurios leiolaémidos (Liolaemus sp.) en
toda la secuencia, con el agregado de un leiosáurido
(Pristidactylus cf. P. scapulatu en las unidades 1 y 2 y un
gekkónido ([Homonota cf. H. darwini] en la Unidad 1).
Los porcentajes de los restos afectados por la corrosión
digestiva en las cuatro unidades variaron entre
5,7% y 22% (Tabla 3), y en todos los casos esta incidió
en forma ligera, ya que presenta un leve poceado. Los
valores de abundancia relativa de los elementos fueron
similares en casi todas las unidades. No obstante, la
Unidad 1 fue la que más se diferenció, ya que exhibe
una mejor representación de maxilares y menor
abundancia de tibias. Con respecto a las fracturas, se
destaca que no se halló ningún cráneo completo en
toda la secuencia. Sin embargo, el nivel de fractura
de los elementos poscraneales fue escaso en todas
las unidades y tanto las vértebras como las cinturas
estaban completas. Se destaca que todos los elementos
poscraneales rotos presentaron superficies de fractura
con ángulos agudos y bordes ásperos y sin marcas de
digestión. Es interesante destacar que no se registraron
restos con marcas de corte en ninguna unidad y que
sólo tres huesos de la Unidad 1 presentaron rastros de
termoalteración (Tabla 3).
Tabla 3. Modificaciones tafonómicas sobre las superficies de los
especímenes.

Nota: Los valores están expresados en porcentajes.
En lo que concierne a los procesos posdepositacionales,
cabe aclarar que en toda la secuencia no se
hallaron restos con evidencias de meteorización. Por
otro lado, los conjuntos presentaron restos óseos con
marcas de raíces del tipo "Sphenoichnia", y exhiben
un incremento gradual desde las unidades más superficiales
hasta las más profundas (Tabla 3). Asimismo,
se registraron especímenes con marcas de corrosión
sedimentaria por humedad del suelo, que muestran
un patrón similar al anterior (Tabla 3). En toda la secuencia
no se hallaron evidencias de redondeamientos
en los extremos ni de protuberancias de los huesos.
Aves
En relación con el conjunto de aves se registró
un NISP de 89, y se identificó un total de 12 taxones.
La mayoría de las aves presentes corresponden a
Passeriformes (56,6%), aunque existe una abundancia
significativamente menor de Columbidae (11,1%), aves
rapaces diurnas/nocturnas, entre ellas Cathartidae y
Strigiformes (14,5%), aves de ambientes acuáticos tales
como Podicipedidae, Anatidae, Charadriidae (7,7%), y
el restante 10,1% corresponde a aves que no pudieron
ser determinadas en ningún nivel taxonómico.
La muestra avifaunística presenta registro anatómico
de todo el esqueleto, aunque en frecuencias
diferenciales. En ese sentido, el esqueleto axial representa
el 15,7%; el miembro anterior, el 47,2%; y
el miembro posterior, el 37,1% (considerando todos
los taxones de aves presentes en el sitio). El porcentaje
más alto se encuentra representado por elementos
de densidad ósea alta (cf. Savanti 1994), tales como
el tibiotarso (14,8%), tarsometatarso (10,2%), ulna
(13,6%) y húmero (12,5%). Estos porcentajes, en la
mayoría de los casos, corresponden a Columbidae y
Passeriformes. El 56% de los elementos de este grupo
taxonómico se encuentra fragmentado, hecho que
dificultó la identificación anatómica y taxonómica de
algunos de ellos. Solamente un 10% de los elementos
óseos fracturados presentan la superficie
de fractura redondeada debido a la acción que
ejercen sobre los bordes los ácidos digestivos.
En el resto de la muestra las fracturas predominantes
son las de tipo transversal, generadas
cuando el hueso se encuentra en estado seco.
Asimismo, se han registrado marcas de corrosión
digestiva, que varían entre 28% y 21%.
Estas se caracterizan por la presencia de hoyos
y depresiones con bordes redondeados localizados
en los extremos de los huesos.
Tanto
las alteraciones digestivas como diagenéticas
se registraron en taxones de tamaño pequeño
como Charadriiformes, Columbiformes y
Passeriformes y no se identificaron en taxones
medianos como Podicipedidae, Anatidae,
Cathartidae. Asimismo, las marcas de carnívoros
(6,7%) fueron observadas en restos
asignados a T. alba, Columbidae, Anatidae y
Passeriformes. En ningún espécimen se identificaron
marcas de corte por acción humana.
Los agentes y procesos posdepositacionales
identificados que tuvieron incidencia en la formación del registro avifaunístico fueron la acción
corrosiva de raíces y los efectos de la humedad del
suelo, que habrían contribuido a la rápida alteración
de elementos óseos (Tabla 3). Además, en todas las
unidades se observaron elementos con marcas de
roído, registradas en los especímenes asignados a T. alba y Passeriformes y en las unidades 2 a 4 algunos
especímenes exhibieron alteraciones térmicas en restos
de Passeriformes, Strigiformes y Anatidae (Tabla
3). Finalmente, escasos restos presentaron evidencias
de meteorización, de modo que el 99% de los especímenes
correspondieron al estadio 0, según la escala
de Behrensmeyer et al. (2003).
Micromamíferos
En cuanto a los mamíferos de tamaño pequeño,
con un NISP de 5527, los roedores cricétidos
sigmodontinos dominan toda la secuencia,
siendo Phyllotis xanthopygus y Euneomys
chinchilloides los taxones más representados,
seguidos por Eligmodontia sp., Abrothrix hirta, Chelemys macronyx y Loxodontomys micropus.
En proporciones menores se encuentra el roedor
histricognato Ctenomys sp.
Los porcentajes de los restos afectados por
la corrosión digestiva se encuentran en toda
la secuencia, y corresponden principalmente
a la categoría ligera (Tabla 3). En tal sentido,
en el 23,6% de los incisivos la corrosión se
concentró en los extremos y el 25,4% de los
molares exhibieron cúspides con un contorno
más redondeado y el 0,3% mostraron un ligero
poceado en el esmalte. El 29,3% de los restos
poscraneales exhibieron evidencias de poceado
en las superficies articulares (Figura 2a) y
el 6,5% mostraron un aspecto de abrasión o
pulido.

Figura 2. Modificaciones tafonómicas predepositacionales y
posdepositacionales en especímenes hallados en el sitio LS3; a: epífisis
proximal de fémur de Sigmodontinae con marcas de digestión ligera en
trocánter mayor; b: diáfisis y epífisis distal de fémur de Sigmodontinae
con marcas de raíces; c: incisivo de Sigmodontinae con marcas de
corrosión sedimentaria en dentina; d: ilion de Lama guanicoe con
marcas de mamífero carnívoro en cresta ilíaca; e: acercamiento de
la foto anterior que muestra el detalle de las marcas de carnívoro; f:
diáfisis de fémur de L. guanicoe con negativos de impacto.
En las cuatro unidades se observó un patrón
similar de representación de partes esqueletarias;
los elementos más abundantes fueron las
mandíbulas, húmeros, fémures y tibias, y los
menos abundantes fueron las costillas y los
huesos del autopodio. La proporción de restos
fracturados en las distintas unidades fue
similar (U1=42,2%; U2=39,3%; U3=39,9%;
U4=38,5%). Se halló un sólo cráneo completo
en la Unidad 4, y el 93,9% de los mismos
especímenes correspondieron a maxilares
sin arcos zigomáticos. En todas las unidades
se registraron escasas mandíbulas completas
(U1=13,7%; U2=11,4%; U3=10,1%;
U4=14,3%). Los valores de fractura de los
elementos poscraneales fueron similares en
todas las unidades, variando desde 38,4% en
la U1 hasta 50,8% en la U2. Se destaca que
todas las unidades presentaron una mayor proporción
de los elementos poscraneales rotos con superficies
de fracturas con ángulos agudos y bordes ásperos
(U1=96,6%; U2=91,7%; U3=96,6%; U4=97,4%). Por
otra parte, la totalidad de los elementos del autopodio
(calcáneos, astrágalos, metapodios y falanges) estaban
completos y se hallaron pocas vértebras fracturadas.
Sin embargo, los elementos más frágiles, como
las costillas, escápulas, pelvis y radios, presentaron
un grado alto de fractura (U1=77,3%; U2=72,7%;
U3=76,9%; U4=75,2%).
Por otra parte, se destaca que en ninguno de los
tres conjuntos se registraron restos con marcas de cortes
en toda la secuencia, y que escasos restos presentaron
evidencias de termoalteración (Tabla 3). En lo que concierne a los procesos posdepositacionales,
se encontraron muy pocos restos con evidencias
de meteorización a lo largo de toda la secuencia. Por
otra parte, no se observaron especímenes óseos con
marcas de pisoteo. Se registraron restos con marcas
de raíces de tipo "Sphenoichnia" en todas las unidades
(Tabla 3, Figura 2b). Asimismo, se recuperaron
especímenes con evidencias de corrosión sedimentaria
(Figura 2c) a lo largo de toda la secuencia (Tabla
3) y se hallaron restos con impresiones de óxido de
manganeso sólo en la U4 (Tabla 3). En ninguna parte
de la secuencia se observaron restos óseos con redondeamiento
en los extremos ni protuberancias de los
restos óseos y dentarios, lo que indica que no actuaron
los efectos de la abrasión por transporte hídrico,
desplazamientos y rozamientos de los restos contra
el sedimento.
Mamíferos mediano-grandes
Se halló un NISP de 406 vertebrados medianos y
grandes; sin embargo, este número de especímenes
puede ser producto de la elevada fragmentación de estos.
Debido a este estado, sólo 61 especímenes fueron
identificados taxonómicamente, dentro de los cuales,
21 corresponden a la categoría Camelidae (MNI=1) y
uno sólo a Lycalopex griseus. Dentro de este subconjunto se registró una pelvis
de guanaco y siete vértebras articuladas en posición
anatómica, sin evidencias de procesamiento antrópico,
y con marcas de carnívoro de tamaño mediano (Figura
2d). Sin embargo, los mamíferos carnívoros no afectaron
intensivamente a este conjunto, de modo que
sólo en la U1 y la U2 se encontraron este tipo de
marcas y en proporciones muy bajas (2,6% y 0,85%,
respectivamente). El tamaño de los hoyuelos y la
distancia entre ellos coinciden con las cúspides del
segundo y tercer molar de L. griseus (Figura 2e). En
cuanto al registro de especímenes termoalterados, este
es muy escaso, al igual que en los grupos taxonómicos
descriptos anteriormente. Por otra parte, se halló únicamente
un fragmento de diáfisis de fémur con fractura
helicoidal y negativo de impacto en la unidad 1, que
podría ser de origen antrópico (Figura 2f).
En cuanto a los procesos posdepositacionales,
la meteorización afectó de manera más intensiva a
aquellos huesos que se encuentran en la unidad más
profunda de la secuencia, mientras que en la U1 y U2
los elementos exhiben los valores más bajos de esta
categoría (Tabla 3). Además, sólo se registraron dos
especímenes con marcas de acción de roedores en la
Unidad 1, y la acción de las raíces afectó a los restos
óseos en todas las unidades de manera relativamente
homogénea. Finalmente, en toda la secuencia hay
evidencias de corrosión sedimentaria, especialmente
en los niveles más profundos (Tabla 3).
DISCUSIÓN
Los huesos hallados en cuevas o refugios rocosos
están frecuentemente muy alterados, y uno de los
problemas más usuales para la interpretación de estos
conjuntos arqueofaunísticos es la dificultad para diferenciar
las modificaciones de los conjuntos producidos
por la depredación y las modificaciones acaecidas por
los procesos posdepositacionales (e.g., Andrews 1990;
Mondini 1995, 2000). Los estudios tafonómicos realizados
en el sitio LS-3 se focalizaron en discutir en
qué medida los seres humanos fueron responsables de
la acumulación ósea del conjunto hallado y, en este
sentido, evaluar si este es pertinente para discutir el
modelo de intensificación propuesto para la región
del sur de Mendoza.
La evaluación del consumo de vertebrados medianos
y pequeños en poblaciones prehistóricas es compleja,
dada la multiplicidad de procesos que actúan en
la depositación del conjunto faunístico. La dificultad
en la recuperación de estos especímenes durante el
trabajo de campo, las complicaciones en la identificación
anatómica y taxonómica y los diversos agentes
involucrados en la depositación de estos, entre otros
factores, hicieron poco atractivo su estudio hasta no
hace mucho tiempo. Sin embargo, el reconocimiento
desde la teoría arqueológica, de la necesidad de estudiar
la diversidad en las prácticas de subsistencia, y el
desarrollo de un corpus metodológico en la tafonomía,
hacen necesario y factible abordar este tipo de fauna
(Andrews 1990; Lyman 1994; Kelly 1995).
Discutimos aquí en qué medida los depredadores
naturales, la actividad humana y los procesos posdepositacionales
tuvieron incidencia en la formación de
los conjuntos óseos de este sitio.
Depredación natural
Para evaluar la incidencia de depredadores naturales
(no antrópicos) en la formación del sitio LS-3, se
han analizado variables como la abundancia de los
distintos grupos taxonómicos, la abundancia relativa
de elementos anatómicos y las evidencias de alteración
por ácidos digestivos.
Como se ilustra en la Tabla 2, los micromamíferos
tuvieron una representación ampliamente mayoritaria
entre los microvertebrados del sitio LS-3, en tanto que
los reptiles y las aves correspondieron a una pequeña
fracción del total. Esto resulta coincidente con la dieta
de T. alba, la cual es una rapaz crepuscular y nocturna
que se alimenta principalmente de micromamíferos
y ocasionalmente de otros microvertebrados como
anfibios, reptiles y aves, y también puede consumir
artrópodos (véase Bellocq 2000 y literatura allí citada). Algo similar ocurre con otras Strigiformes (Athene cunicularia y B. v. magellanicus), en casos registrados en
Patagonia (Nabte et al. 2006 y referencias allí citadas).
En cuanto a los valores de abundancia relativa de
elementos anatómicos de los reptiles, se pudo observar
que, a lo largo de la secuencia, las mandíbulas,
húmeros y fémures fueron los elementos más abundantes;
mientras que las vértebras, cúbitos, radios y
peronés fueron los menos frecuentes. Estos resultados
se encuentran acordes con los datos presentados por
Castillo et al. (2001), en un análisis de restos fósiles de
reptiles depositados por T. alba, provenientes del sitio
paleontológico Cueva del Llano en las Islas Canarias,
España (Figura 3 a).
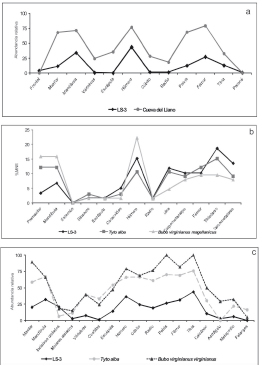
Figura 3. Patrones de elementos esqueletales en promedio
de las unidades del sitio LS-3 en reptiles (a), aves (b) y
micromamíferos (c) y su comparación con una muestra de
reptiles promedio de todas las unidades del sitio Cueva del
Llano (Castillo et al. 2001), restos de aves de egagrópilas de Tyto alba y Bubo virginanus magellanicus (Fernández et al. 2009) y restos de micromamíferos de egagrópilas de T. alba (Andrews 1990) y B. v. virginianus (Gómez 2005).
En relación con la representación anatómica del
registro avifaunístico, estudios de Ericson (1987),
Livingston (1989) y Bovy (2002) sugieren que la mayor
frecuencia de huesos del miembro anterior sobre otras
partes anatómicas puede implicar prácticas culturales
de selección. Estas prácticas actúan seleccionando
otras partes de la presa durante el procesamiento y
consumo, por lo que el miembro anterior tiende a estar
sobrerrepresentado y casi sin modificaciones. Los mencionados
investigadores argumentan que el miembro
anterior puede ser descartado durante el despostamiento
de las presas, mientras que el miembro posterior
se fragmentaría durante la etapa de procesamiento y
cocción (Bovy 2002 y bibliografía allí citada). Por otro
lado, Ericson (1987) propone que los conjuntos que
mantienen una relación proporcional entre miembros
anteriores y posteriores podrían tener un origen natural
de depositación. Esto es coincidente con el patrón
que muestra el subconjunto de paseriformes de
LS-3 (miembro anterior (45,8%) y miembro posterior
(42,4%). Además, en relación con el MNE% utilizado
para identificar el agente responsable, la tendencia que
presenta este sitio se asemeja más a aquellas muestras
actuales de T. alba en la representación de todos los
elementos (Figura 3 b). En este sentido, la representación
de los distintos elementos anatómicos recuperados
en el sitio muestra que el conjunto de LS-3 tiene
una fuerte similitud con la curva de los conjuntos de
aves generados por Strigiformes, principalmente con T. alba (Figura 4).

Figura 4. Frecuencia (%MNE) en las distintas unidades
anatómicas del conjunto avifaunístico. Comparado con
los modelos de Bocheński et al. (1998) para egagrópilas y
Bocheński y Tornberg (2003) para restos no ingeridos.
En relación con la abundancia relativa de elementos
de micromamíferos, las mandíbulas, fémures, tibias
y húmeros fueron los elementos mejor representados;
y las vértebras, astrágalos, metapodios y falanges, los
menos abundantes (Figura 3c). Estos patrones de abundancia
relativa son similares a las muestras de T. alba (Figura 3c). No obstante, a pesar de esta semejanza,
la curva de la muestra de LS-3 es más baja que la de T. alba, lo que sugiere que la acción de los agentes
posdepositacionales alteró la composición original de
los conjuntos, reduciendo la preservación integral del
éstos, aunque manteniendo las proporciones de los
elementos.
Los especímenes de reptiles mostraron porcentajes
bajos de digestión en restos craneales y moderados
en restos poscraneales, y se destaca que, en todos
los casos fueron encuadrarse bajo la categoría ligera
(Figura 5a). No obstante, a pesar de que los restos
fueron afectados de esta manera, los porcentajes de
incidencias son contrastantes con los datos presentados
por Castillo et al. (2001) sobre acumulaciones de reptiles fósiles generadas por T. alba, donde los autores
no registraron evidencias de digestión. Esto desprende
la posibilidad de que los reptiles de LS-3 hayan sido
acumulados por T. alba y otros depredadores, aunque
no se cuenta con datos actualísticos de reptiles acumulados
por otras especies de aves rapaces que habiten
el área, para poder precisar el agente tafonómico. La
Figura 5b exhibe la comparación entre los elementos
con marcas de digestión en la muestra avifaunística
y los elementos óseos hallados en egagrópilas de T. alba y B. v. magellanicus (Fernández et al. 2009). En
este sentido, los porcentajes obtenidos muestran tendencias
similares a los valores de muestras actuales de T. alba. Las marcas de corrosión por digestión en la
superficie de los huesos presentan una incidencia de
aproximadamente el 25% de NISP total de la muestra
de aves Passeriformes. Asimismo, los especímenes
arqueológicos presentan la mayor corrosión de
ácidos gástricos en los huesos largos. Por otra parte,
la ausencia de marcas de ácidos gástricos en cráneo
y vértebras en las muestras arqueológicas contrasta
con las muestras actuales. Sin embargo, esta ausencia
podría deberse problemas de preservación diferencial,
ya que encontramos en este sitio una baja frecuencia
de dichos elementos, que puede ser consecuencia de
su escasa densidad mineral (Savanti 1994).

Figura 5. Valores de corrosión digestiva en restos de reptiles
(a) aves (b) y micromamíferos (c) recuperados de LS-3. Datos
de Tyto alba y Bubo virginianus magellanicus tomados de
Fernández et al. (2009).
Estudios actualísticos sobre tafonomía de varias
especies de Strigiformes revelan que estas aves se
caracterizan porque la acción de sus ácidos gástricos
afecta de manera ligera a moderada a los restos óseos
y dentarios de micromamíferos (e.g., Andrews 1990;
Gómez 2007). En congruencia, a lo largo de toda la
secuencia de LS-3 se observaron principalmente restos
óseos con corrosión digestiva ligera y moderada
(Figura 5c). Por un lado, investigaciones realizadas sobre
restos óseos de micromamíferos consumidos por T. alba (e.g., Andrews 1990; Gómez 2007; Fernández et al. 2009) concluyen que esta rapaz produce niveles
de digestión correspondientes a la categoría ligera.
Por otro lado, Gómez (2005) y Montalvo y Tejerina
(2010) hallaron que B. v. virginianus y A. cunicularia, respectivamente, causaban mayor acción digestiva, y
las ubican entre las categorías ligera y moderada. Esto
indica que los restos de micromamíferos de LS-3 podrían
haber sido acumulados por varias aves rapaces.
Actividad antrópica
En cuanto a las aves medianas, entre las cuales se
incluyen aves de ambientes acuáticos (Podicipedidae,
Anatidae, Charadriidae) y aves rapaces (Cathartidae,
Falconidae, Accipitridae, Strigidae, Tytonidae), se esperaba
que tuvieran evidencias de consumo antrópico,
sin embargo, esto no sucedió. Estas muestran
evidencias de otros agentes tafonómicos (i.e., acción
de carnívoros, corrosión digestiva y la escasa selección de partes anatómicas). Además, la presencia de especímenes
óseos de T. alba en este sitio podría sugerir
la muerte natural in situ (Pardiñas 1999) o como presa
de otros depredadores "predators as preys" (Andrews
1990: 27) y que, como consecuencia de esto último,
en ciertas oportunidades aparecen mezclados depósitos
con restos de depredadores y presas.
En lo que respecta a los micromamíferos, también
esperábamos evidencias de consumo antrópico debido al contexto cronológico en el que se encuentran los
depósitos de este sitio, es decir, debido a la existencia
del mencionado proceso de intensificación. Sin
embargo, en ninguna unidad arqueológica se halló
evidencia directa de consumo humano (e.g., marcas
de corte). Asimismo, si bien se observa un leve aumento
de restos termoalterados en las unidades más
profundas, estos nunca alcanzan el 3% del NISP total,
y corresponden a roedores sigmodontinos. Relacionado
con esto, Pardiñas (1999) plantea que un conjunto de
huesos termoalterados de micromamíferos, asociados
al consumo humano, presenta entre otras cosas un
elevado porcentaje de restos afectados y una dominancia
de roedores histriocognatos. Por ende, la posible
vinculación de los restos quemados provenientes de
las unidades arqueológicas al consumo humano queda
prácticamente descartada. Procesos naturales, limpieza
del recinto, el encendido de fuego sobre las egagrópilas
y su posible uso como combustible, pueden ser
una mejor explicación al tema (ver Pardiñas, 1999).
Las lechuzas y búhos posiblemente sean los depredadores
que más contribuyen a generar acumulaciones
de microvertebrados en sitios arqueológicos y paleontológicos
de cuevas o refugios rocosos (e.g., Andrews
1990), y dentro de este grupo, T. alba ocupa un lugar
preponderante (Andrews 1990; Saavedra y Simonetti
1998, entre otros). Los taxones hallados a lo largo de
la secuencia están dominados por micromamíferos de
tamaño pequeño y mediano (sensu Pardiñas 1999), lo
cual coincide con el rango de tamaño de presas de
las aves Strigiformes (Pardiñas 1999 y literatura allí
citada). En tal sentido, cabe mencionar que T. alba, B. v. magellanicus y A. cunicularia son las especies más
frecuentes habitando la zona en la actualidad.
El hallazgo de restos óseos de Strigiformes indeterminados
en las unidades 3 y 4 y de T. alba en las
unidades 2 y 3 constituye un elemento de cabal importancia.
La presencia de restos de aves rapaces en
sitios arqueológicos no establece una evidencia directa
de que el lugar haya servido de habitación para estas.
Sin embargo, en ciertas ocasiones, las lechuzas, búhos,
halcones, águilas, etc., mueren por causas naturales
en sus sitios de nidificación y/o descanso, donde sus
restos pueden acumularse con los de sus presas (e.g.,
Andrews 1990; Pardiñas 1999).
En cuanto a los vertebrados de tamaño mediano y
grande, tal como mencionamos previamente, fueron
registradas una pelvis de guanaco y siete vértebras
articuladas en posición anatómica. Las evidencias sobre
el origen antrópico de estos especímenes se presentan
ambiguas y por eso discutimos cuáles podrían
haber sido los agentes responsables de la acumulación
de los especímenes de este grupo taxonómico.
Para ello utilizamos algunos de los criterios definidos
por L´Heureaux y Borrero (2002) para reconocer conjuntos
óseos antrópicos y no antrópicos de guanaco
en Patagonia. Estos son: presencia de modificaciones
culturales en los huesos como indicador de actividad
antrópica, asociación de los elementos con artefactos,
proporción diferencial de huesos del esqueleto axial/
apendicular y proporción diferencial de elementos
enteros/fragmentados.
En cuanto a la presencia de modificaciones culturales
en los huesos, se ha observado que los restos
de Camelidae del sitio LS-3 presentan únicamente un
negativo de impacto en uno de sus huesos largos.
Aunque efímera, esta evidencia sería concordante con
un origen cultural de estos restos. Lo mismo ocurre con
lo concerniente a la asociación del conjunto óseo con
artefactos, ya que se recuperó un sólo artefacto lítico
y restos de carbón en todo el sitio.
En lo concerniente a la proporción diferencial de
huesos del esqueleto axial/apendicular, se sostiene que
en los sitios arqueológicos hay una predominancia
del esqueleto apendicular sobre el axial, por ello, en
aquellos sitios en los que haya una importante proporción
de elementos del esqueleto axial existen mayores
probabilidades de que sea de origen no antrópico. En
el sitio LS-3, la mayor proporción de los elementos
pertenecen al esqueleto axial (N = 13). En cuanto al
esqueleto apendicular, tres elementos corresponden
a carpianos, y cuatro, a huesos largos. Es decir que,
según este criterio, el conjunto en cuestión muy probablemente
sea de origen natural. En cuanto a la proporción
diferencial de elementos enteros/fragmentos, en
los sitios arqueológicos la mayoría de los elementos se
encuentran fragmentados como producto de su procesamiento
antrópico. Una alta proporción de elementos
completos o una baja fragmentación de ellos indicarían
mayores probabilidades de que estemos ante un
conjunto depositado naturalmente. En el caso de LS3,
la mayoría de los elementos de guanaco se encuentran
completos, como los carpianos y tarsianos y un metacarpo.
Algunos se encuentran casi completos, como
la pelvis y vértebras articuladas halladas en la misma
unidad. Es decir que, excepto por la diáfisis de fémur
con fractura helicoidal y negativo de impacto, el estado
de baja fragmentación que presentan los elementos
indicaría un origen natural de estos.
En síntesis, de los cuatro criterios mencionados
para identificar el origen cultural o natural de este
conjunto, dos se encuentran a favor de un origen cultural
y dos a favor de un origen natural. El sentido del
análisis de los criterios mencionados no es aplicarlos
para ver hacia dónde se inclina la balanza y definir
automáticamente si se trata de un conjunto antrópico
o natural, sino evaluarlos en conjunto con las otras
líneas de evidencia del sitio. En este sentido, sostenemos
que la existencia de algunos de los elementos
de guanaco tiene un origen antrópico, pero que el
aprovechamiento de estos ha sido poco intensivo.
Procesos posdepositacionales
La ausencia de huesos de microvertebrados con
evidencias de meteorización a lo largo de toda la secuencia
indica que posiblemente los restos han tenido
un enterramiento rápido. Andrews (1990) observó,
sobre la base de estudios experimentales sobre restos
de micromamíferos, que el pisoteo es un proceso muy
destructivo y del cual emergen patrones de fractura,
tales como la ausencia de cráneos completos, reducción
del número de maxilares e importante grado de
fractura, pero con los elementos más pequeños en
buen estado de integridad. Al respecto, gran parte de
los restos presentaron fracturas con ángulos agudos y
bordes ásperos sin huellas de digestión, y la mayoría
de los elementos de menor tamaño (i.e., vértebras,
calcáneos, astrágalos, metapodios y falanges) estaban
completos. De esta manera, posiblemente este patrón
esté más asociado a procesos posdepositacionales
como el pisoteo que a los agentes acumuladores.
Además, la exigua cantidad de huesos con evidencia
de meteorización a lo largo de toda la secuencia
sugiere que los restos posiblemente hayan tenido un
enterramiento rápido, luego de haber sido alterados
por el pisoteo. Eventos de estos tipos son habituales
en las cuevas o refugios rocosos, debido a que son
unidades espaciales con circulación restringida, donde
los restos fósiles están en parte protegidos de los
agentes meteóricos (i.e., sol, lluvia, viento y cambios
de temperatura), pero a su vez están más expuestos a
los efectos del pisoteo (e.g., Andrews 1990; Pardiñas
1999).
Con respecto a otros procesos posdepositacionales,
cabe destacar que tanto las marcas de raíces como la
corrosión sedimentaria evidenciaron un incremento a
medida que aumentó la profundidad del sitio, que es
más notorio en las unidades 3 y 4. Andrews (1990) observó
que bajo condiciones de permanente humedad
en el sedimento, los huesos de micromamíferos son
afectados en su totalidad, mientras que en condiciones
de sequedad, son mejor preservados. En congruencia,
se registraron restos afectados por óxido de manganeso
sólo en la parte más profunda del sitio (U-4). En este
alero, la alta humedad del suelo y los ácidos húmicos
de las raíces (principalmente en las unidades 3 y
4) produjo una importante alteración en los huesos,
que afectó toda su superficie, contrariamente a lo que
ocurre con la corrosión por digestión, en que los efectos
son localizados (Andrews 1990; Fernández-Jalvo y
Andrews 1992).
En cuanto a la fauna de tamaño grande, el individuo
de camelidae articulado hallado en este sitio
presenta marcas producidas por carnívoros. El tamaño
de dichas marcas y su disposición coinciden con
las que puede producir el zorro gris. Según estudios
actualísticos, un carnívoro de este tamaño podría
haber modificado la superficie a partir de actividades
de carroñeo, pero no haber causado la muerte
ni el desplazamiento de esta porción de la carcasa
(Mondini 1995; Martin 1998 y 2005). Por otra parte,
existen registros de mortandad de guanacos en
reparos en momentos de estrés ambiental durante
el invierno en sitios de Patagonia (Rindel y Belardi
2006). Consideramos que es posible que este animal
haya muerto por causas naturales.
CONCLUSIONES
La importancia del sitio LS-3 a nivel regional radica
en que su cronología es contemporánea con momentos
en los que se ha registrado un proceso de intensificación
en la región, donde se observa la incorporación
de nuevos espacios a los circuitos de movilidad y un
mayor espectro de recursos en la dieta humana, que
incluye fauna de pequeño tamaño y menor rendimiento
económico. Sin embargo, a partir de los análisis
realizados en este sitio concluimos que este no es
adecuado para la evaluación del modelo de intensificación
planteado por Neme (2007), debido a que casi
la totalidad de los restos habrían ingresado al sitio por
causas naturales.
A partir de lo observado en todo el conjunto depositado
en LS-3 sostenemos que los humanos habrían
tenido cierta participación en su formación, pero de
manera muy efímera.
Asimismo, todos los indicadores analizados sugieren
un origen no antrópico de los de microvertebrados
(reptiles, aves y roedores pequeños), los cuales habrían
llegado al sitio por la actividad depredadora de aves
Strigiformes, siendo T. alba la principal especie involucrada,
aunque posiblemente hayan actuado otras especies
tales como B. v. magellanicus y A. cunicularia.
Con respecto al conjunto de vertebrados de tamaño
mediano y grande, de acuerdo con los análisis realizados,
algunos de los restos de Camelidae depositados
en el sitio LS-3 tienen un origen natural, y otros, un
origen cultural.
Concluimos que, al no existir evidencias de consumo
antrópico de la fauna de tamaño mediano y pequeño
y al no observarse un aprovechamiento intensivo
de la fauna de tamaño grande, es incorrecto utilizar
las evidencias de este sitio para evaluar el modelo de
intensificación. Sin un análisis exhaustivo de los procesos
tafonómicos que intervinieron en la formación
de este sitio, la presencia de vertebrados de tamaño
pequeño habría sido interpretada como producto del
consumo humano, hecho que nos hubiese conducido
a una interpretación errónea del registro arqueológico
del sitio LS-3.
Agradecimientos
A Fernando Franchetti, Miriam Ayala y Germán
Moreira por haber ayudado en la separación de los
materiales óseos. A Adolfo Gil y Gustavo Neme por
la lectura crítica del manuscrito. Estas investigaciones
pudieron realizarse gracias al apoyo financiero del
CONICET y de ANPCyT: proyectos IDAC-ICES y PICT
2006-00046. Además queremos agradecer a los tres
evaluadores de este artículo, sus valiosos comentarios
ayudaron a mejorarlo.
REFERENCIAS CITADAS
1. Andrews, P.
1990 Owls, caves and fossils. University of Chicago
Press, Chicago.
2. Behrensmeyer, A. K.
1978 Taphonomic and ecologic information on bone
weathering. Paleobiology 4: 150-162.
3. Behrensmeyer, A. K., C. T. Stayton y R. E. Chapman
2003 Taphonomy and ecology of modern avifaunal
remains from Amboseli Park, Kenya. Paleobiology 29:
52-70.
4. Bellocq, M. I.
2000 A review of the trophic ecology of the Barn Owl in
Argentina. Journal of Raptor Research 34: 108-119.
5. Bettinger, R.
1991 Hunter–Gatherers: Archaeological and Evolutionary
Theory. Plenum Press, Nueva York.
6. Binford, L. R.
1981 Bones: ancient men and modern myths. Academic
Press, Nueva York.
7. Bocheński, Z. M. y T. Tomek
1997 Preservation of bird bones: erosion versus digestion
by owls. International Journal of Osteoarchaeology 7:
372-387.
8. Bocheński, Z. M., K. Huhtala, P. Jussila, E. Pulliainen, R.
Tornberg y P. S. Tunkkari
1998 Damage to bird bones in pellets of Gyrfalcon
Falco rusticolus. Journal of Archaeological Science 25:
425-433.
9. Bocheński, Z. M. y R. Tornberg
2003 Fragmentation and preservation of bird bones in
uneaten food remains of the Gyrfalcon Falco rusticolus.
Journal of Archaeological Science 30: 1665-1671.
10. Bovy, K. M.
2002 Differential avian skeletal part distribution:
Explaining the abundance of wings. Journal of
Archaeological Science 29: 965-978.
11. Cabrera, A. L.
1976 Regiones fitogeográficas argentinas. Enciclopedia
Argentina de Agricultura y Jardinería 1: 1-85.
12. Castillo, C., E. Martín y J. J. Coello
2001 Small vertebrate taphonomy of La Cueva del Llano,
a volcanic cave of Fuerteventura (Canary Islands,
Spain). Palaeogeography, Palaeoclimatology and
Palaeoecology 166: 277-291.
13. Ericson, P. G. P.
1987 Interpretations of archaeological birds remains
a taphonomic approach. Journal of Archaeological
Science 14: 65-75.
14. Fernández, F. J., G. J. Moreira, G. A. Neme y L. J. M. De
Santis
2009 Microvertebrados exhumados del sitio arqueológico
"Cueva Arroyo Colorado" (Mendoza, Argentina):
aspectos tafonómicos y significación paleoambiental.
Archaeofauna 18: 99-118.
15. Fernández, P., I. Cruz y A. I. Forlano
2010 Sitio 37: Una Madriguera de Carnívoro en el
Norte de la Patagonia Andina (Cholila, Provincia de
Chubut, Argentina) En Zooarqueología a principios del
siglo XXI. Aportes teóricos, metodológicos y casos de
estudio, editado por M. A. Gutiérrez, M. De Nigris, P.
Fernández, M. Giardina, A. Gil, A. Izeta, G. Neme, H.
Yacobaccio, pp. 409-417. Del Espinillo, Buenos Aires.
16. Fernández-Jalvo, Y. y P. Andrews
1992 Small Mammal Taphonomy of Gran Dolina,
Atapuerca (Burgos), Spain. Journal of Archaeological
Science 19: 407-428.
17. Fernández-Jalvo, Y. y P. Andrews 2003 Experimental effects of water abrasion on bone
fragments. Journal of Taphonomy 1: 147-163.
18. Giardina, M. A.
2010 El aprovechamiento de la avifauna entre las
sociedades cazadoras-recolectoras del Sur de Mendoza,
un enfoque arqueozoológico. Tesis Doctoral inédita.
Facultad de Ciencias Naturales y Museo, Universidad
Nacional de La Plata, La Plata.
19. Gómez, G.
2005 Analysis of bone modification of Bubo virginianus
pellets from Argentina. Journal of Taphonomy 3: 1-16.
20. Gómez, G. 2007 Predators categorizations based on taphonomic
analysis of micromammals bones: a comparison to
proposed models. En Taphonomy and Zooarchaeology
in Argentina, editado por M. A. Gutiérrez, L. Miotti,
G. Barrientos, G. Mengoni Goñalons, M. Salemme, pp.
89-103. BAR International Series 1601. Archaeopress,
Oxford.
21. Haynes, G.
1980 Evidence of carnivore gnawing on Pleistocene and
Recent mammalian bone. Paleobiology 6 (3): 341-351.
22. Kelly, R. L.
1995 The foraging spectrum. Diversity in hunter-gatherer
lifeway. Simthsonian Institution Press, Washington.
23. Korth, W.
1979 Taphonomy of Microvertebrate Fossil Assemblages.
Annals of Carnegie Museum 15: 235-285.
24. L´Heureux, G. L y L. A. Borrero
2002 Pautas para el reconocimiento de conjuntos óseos
antrópicos y no antrópicos de guanaco en Patagonia.
Intersecciones en Antropología 3: 29-40.
25. Llano, C.
2013 Aprovechamiento de los recursos vegetales entre
las sociedades cazadoras-recolectoras del sur de
Mendoza. Tesis Doctoral inédita. Universidad Nacional
del Comahue, Neuquén.
26. Livingston, S. D.
1989 The taphonomic interpretation of avian Skeletal
Part frequencies. Journal of Archaeological Science 16:
537-547.
27. Lyman, R. L.
1994 Vertebrate taphonomy. Cambridge University Press,
Cambridge.
28. Lyman, R. L. 2008 Quantitative Paleozoology. Cambridge manuals in
Archaeology. Cambridge University Press, Nueva York.
29. Martin, F.
1998 Madrigueras, dormideros y letrinas. En Arqueología
de la Patagonia Meridional. Proyecto Magallania,
compilado por L. A. Borrero, pp. 73-96. Búsqueda de
Ayllu, Concepción del Uruguay.
30. Martin, F. 2005 Frozen Storage, en The Taphonomist´s Corner.
Journal of Taphonomy 4: 55-56.
31. Medina, E. M., P. Teta, y D. Rivero
2012 Burning damage and small-mammal human
consumption in Quebrada del Real 1 (Cordoba,
Argentina): an experimental approach. Journal of
Archaeological Science 39: 737-743.
32. Mengoni Goñalons, G. L.
1999 Cazadores de guanacos de la estepa patagónica.
Sociedad Argentina de Antropología, Buenos Aires.
33. Mondini, N. M.
1995 Artiodactyl prey transport by foxes in Puna rock
shelters. Current Anthropology 36: 520-524.
34. Mondini, N. M. 2000 Tafonomía de abrigos rocosos de la Puna.
Formación de conjuntos escatológicos por zorros y sus
implicaciones arqueológicas. Archaeofauna 9: 151-164.
35. Mondini, N. M. 2003 Modificaciones óseas por carnívoros en la Puna
argentina. Una mirada desde el presente a la formación
del registro arqueofaunístico. Mundo de Antes 3:
87-108.
36. Montalvo, C. I. y P. Tejerina
2009 Análisis tafonómico de los huesos de anfibios
y roedores depredados por Athene cunicularia
(Strigiformes, Strigidae) en La Pampa, Argentina. En
Mamül Mapu: pasado y presente desde la arqueología
pampeana, editado por M. Berón, L. Luna, M. Bonomo,
C. I. Montalvo, C. Aranda y M. Carrera Aizpitarte, pp.
323-334. Del Espinillo, Buenos Aires.
37. Nabte, M. J., S. L. Saba y U. F. J. Pardiñas
2006 Dieta del búho magallánico (Bubo magellanicus)
en el Desierto del Monte y la Patagonia Argentina.
Ornitología Neotropical 17: 27-38.
38. Nagaoka, L.
2002 The effects of resource depression on foraging
efficiency, diet breadth, and patch use in southern New
Zealand. Journal of Anthropological Archaeology 21:
419-442.
39. Neme, G.
2007 Cazadores recolectores de altura en los andes
meridionales: el Alto valle del río Atuel. BAR
International Series 1591. Archaeopress, Oxford.
40. Neme, G. y A. Gil
2008 Faunal exploitation and agricultural transitions in
the South American agricultural limit. International
Journal of Osteoarchaeology 18: 293-306.
41. Outram, A. K.
2002 Distinguishing bone fat exploitation from other
taphonomic proceses: what caused the high level of
bone fragmentation at the middle Neolithic site Ajvide,
Gotland? En The Zooarchaeology of Fats, Oils, Milk
and Dairying. Proceedings of the 9th ICAZ Conference,
editado por J. Mulville y A. K. Outram, pp. 32-43.
Oxbow Press, Durham.
42. Pardiñas, U. F. J.
1999 Tafonomía de microvertebrados en yacimientos
arqueológicos de Patagonia. Arqueología 9: 265-308.
43. Pianka, E
1983 Evolutionary Ecology. Harper and Row, Nueva York.
44. Rindel, D. y J. B. Belardi
2006 Mortandad catastrófica de guanacos por estrés
invernal y sus implicaciones arqueológicas: el sitio
Alero los guanacos 1, Lago Cardiel (Provincia de Santa
Cruz, Argentina). Magallania 34 (1): 139-155.
45. Saavedra, B. y J. A. Simonetti
1998 Small mammals taphonomy: Intraspecific bone
assemblage comparison between South and North
American Barn Owl, Tyto alba, populations. Journal of
Archaeological Science 25: 165-170.
46. Savanti, F.
1994 Las aves en la dieta de los cazadores recolectores
terrestres de la costa fueguina. Temas de Arqueología,
CONICET - Programa de Estudios Prehistóricos, Buenos
Aires.
47. Stephens, D. W. y J. R. Krebs
1986 Foraging Theory. Monographs in Behaviour and
Ecology. Princeton University Press, Nueva Jersey.