Figure 1: Location of the Piuquenes Pass in the Main Andes of Argentina and Chile.
On Darwin's footsteps across the Andes: Tithonian-Neocomian fossil invertebrates from the Piuquenes pass
Beatriz Aguirre-Urreta and Verónica Vennari
Laboratorio de Bioestratigrafía de Alta Resolución, Departamento de Ciencias Geológicas, Universidad de Buenos Aires, Ciudad Universitaria, Buenos Aires. Email: aguirre@gl.fcen.uba.ar
ABSTRACT
The aim of this work is to summarize the modern knowledge of the geology of the Piuquenes Pass, in the Main Andes of Argentina and Chile, and to describe a small fauna of Tithonian-Neocomian invertebrates mostly represented by ammonites. The present knowledge of the region is compared with Darwin's, as expressed in his famous book on the Geology of South America.
Keywords: Main Andes; Mesozoic; Ammonites; Piuquenes Pass; Darwin.
RESUMEN: Tras las huellas de Darwin a través de los Andes: invertebrados fósiles del Tithoniano-Neocomiano del paso de Piuquenes. El objetivo de este trabajo es presentar una breve revisión moderna de la geología del Paso de Piuquenes en los Andes Principales de Chile y Argentina y describir una pequeña fauna de invertebrados del Tithoniano-Neocomiano compuesta principalmente por amonites. Se compara también el conocimiento de esta región con la referida por Darwin en su famoso libro sobre la geología de América del Sur.
Palabras clave: Andes Principales; Mesozoico; Amonites; Paso de Piuquenes; Darwin.
INTRODUCTION
During his 5 years' voyage around the
world on board the HMS Beagle, Darwin
spent more than 3 years on shore.
This was most probably due to his interest
in exploring new regions, but also to
avoid the strong sea-sickness that annoyed
him when sailing. During the Beagle
years and some time later Darwin was
foremost a geologist, though we are used
to think of him as a pure biologist. Thus,
his enormous collection of fauna and
flora came second to his interest as can
be seen in the fact that he gave most of
it to different specialists after his return
to England.
In March-April 1835, Darwin undertook
a long ride to examine the geology of the
Andes. He started the journey in Valparaiso
on the Pacific coast, crossed the
Andes along the Piuquenes and Portillo
passes, reached Mendoza city and went
back to Chile by the Cumbre or Uspallata
Pass (Fig. 1). This trip was probably one
of the most valuable to him from a
scientific point of view as can be seen in
the long and detailed letters he sent both
to Professor Henslow and to his sister
Susan. The letter to Henslow begins
thus: "I have just returned from Mendoza,
having crossed the Cordilleras by two passes.
This trip has added much to my knowledge of
the geology of the country. Some of the facts, of the truth of which I in my own mind feel fully
convinced, will appear to you quite absurd& incredible", followed by several paragraphs
devoted to the structure of the cordillera
(Burkhardt and Smith 1985).
Figure 1: Location of the Piuquenes Pass in the Main Andes of Argentina and Chile.
One hundred and sixty years later, in the summer of 1995 one of us (BAU) followed the steps of Darwin along the Piuquenes Pass, checking the stratigraphy and collecting fossils. Thus, the objective of this work is to summarize the modern knowledge of the geology and palaeontology of the region of the Piuquenes Pass, in the Main Andes of Argentina and Chile, comparing these views with Darwin's (1846), as expressed in his famous book on the Geology of South America.
REGIONAL GEOLOGICAL SETTING
The Piuquenes (Darwin always spelled it
as Peuquenes) Pass runs across the boundary
between Argentina and Chile in the
Main Central Andes (33°38'S, 69°52'W, Fig. 1). It reaches 4,030 m a.s.l., and
though it is the international divide, the
highest pass is Portillo, located some 24
km to the east, with 4,374 m a.s.l.
The geology of this part of the Main
Andes was studied in detail by Polanski
(1964) as part of the mapping program
of the Geological Survey of Argentina.
It is characterized by a normal subduction
zone, where the oceanic Nazca plate
subducts beneath South America at an
angle of 30° (Jordan et al. 1983). As a
result of that the Mesozoic sequence is
heavily deformed in a series of thrust slices
(see Giambiagi et al. 2009). These
thrusts produced an imbricated sequence,
where the strata are repeated several
times, creating a complex structure.
The basement of the Andes at this latitude
is composed of metamorphic rocks of
Precambrian age. Carboniferous marine
successions devoid of fossils and some
rhyolitic tuffs and lavas of Permian and
Triassic age are unconformably overlying
the basement (Polanski 1964). The Mesozoic
sequences, which record a Pacific
marine transgression, constitute different
stratigraphic cycles described by Groeber
(1953), Malumián and Ramos (1984) and
Riccardi (1983, 1988).
These sedimentary successions are covered
by extensive Cenozoic volcanic and
pyroclastic rocks, including some Recent
volcanic centers, such as San José and
Marmolejo volcanoes. Isolated patches
of colluvial and alluvial deposits are widely
distributed in the region (Fig. 2).

Figure 2: Geological map of the region of the Piuquenes Pass, Main Andes (from Pángaro et al. 1996).
THE GEOLOGY OF THE PIUQUENES PASS
A geological map of the Piuquenes Pass
region is shown in figure 2 (Pángaro et al.
1996), where Darwin's itinerary is marked,
and the rock succession according
to Darwin (1846) is also shown. He described the following: "The ridge of Peuquenes, which divides the waters
flowing into the Pacific and Atlantic oceans,
extends in a nearly N.N.W. and S.S.E. line;
its strata dip eastward at an angle of between
30° and 45°, but in the higher peaks bending
up and becoming almost vertical. Where the
road crosses this range, the height is 13,210 feet
above the sea-level, and I estimated the neighbouring
pinnacles at from 14,000 to 15,000 feet.
The lowest stratum visible in this ridge is a red
stratified sandstone [P]; on it are superimposed
two great masses [Q and S] of black, hard,
compact, even having a conchoidal fracture, calcareous,
more or less laminated shale, passing
into limestone: this rock contains organic remains,
presently to be enumerated. The compacter
varieties fuse easily in a white glass; and this
I may add is a very general character with all the
sedimentary beds in the Cordillera: although this
rock when broken is generally quite black, it
everywhere weathers into an ash-grey tint. Between
these two great masses [Q and S], a bed
[R] of gypsum is interposed, about 300 feet in
thickness, and having the same characters as
heretofore described. I estimated the total thickness
of these three beds [Q, R, S] at nearly
3000 feet; and to this must be added, as will be
immediately seen, a great overlying mass of red
sandstone.
In descending the eastern slope of this great central
range, the strata, which in the upper part dip
eastward at about an angle of 40°, become more
and more curved, till they are nearly vertical; and
a little further onwards there is seen on the further
side of a ravine, a thick mass of strata of
bright red sandstone [T], with their upper extremities
slightly curved, showing that they were
once conformably prolonged over the beds [S]: on
the southern and opposite side of the road, this
red sandstone and the underlying black shaly
rocks stand vertical, and in actual juxtaposition" (Darwin 1846, p. 179-180, pl. 1,
sketch 1, see Giambiagi et al. 2009).
The red sandstones of P correspond to
the Río Damas Formation in Chile
(Tordillo Formation in Argentina). Q and
S of Darwin (1846) are the Lo Valdés
Formation in Chile or the Mendoza
Group in Argentina, where it is composed
by the Vaca Muerta, Chachao and
Agrio Formations. R, the gypsum bed, is
the Auquilco Formation of Argentina
(Río Colina Formation in Chile) of Oxfordian-Kimmeridgian age, and T is
again the Tordillo Formation. It is obvious
now that Darwin did not quite
understand the complex structure of the
region, as can be seen in the geological
map of figure 2, where a western backthrust
puts the Auquilco Formation on
top of the Agrio Formation and a
second fore-thrust repeats again the
Agrio and Chachao Formations (Pángaro
et al. 1996, Giambiagi et al. 2009).
A section was measured on the Argentinean
slope (Fig. 3), along the northern
bank of the Arroyo Piuquenes (Aguirre-Urreta 1996; see location in Fig. 2). The
top beds of the Tordillo Formation
(Kimmeridgian) are sandstones interbedded
with siltstones and green shales. The
Vaca Muerta Formation begins with a
few meters of limestone with algal lamination,
followed in its lower part by black
papyraceous shales with abundant calcareous
nodules with three fossiliferous
levels, from bottom to top with: Virgatosphinctes
aff. V. denseplicatus rotundus
Spath, 1931, Pseudolissoceras zitteli (Burckhardt,
1903) and Aulacosphinctes proximus
(Steuer, 1897). These shales grade upwards
to dark grey, thinly laminated limestones,
reaching more than 110 metres
(Fig. 3). The age of the Vaca Muerta
Formation in the area is Early to Middle-Late? Tithonian. The Chachao Formation
- 40 metres thick - is composed by
massive oyster coquinas interbedded
with shales bearing Parodontoceras calistoides
(Behrendsen, 1891) of Late Tithonian
age in its lower part. The Agrio Formation
is some 120 metres thick in the measured
section but is incomplete, as it is
cut by a thrust that puts the Chachao
coquinas on top of its upper part. The
lower part is mostly composed by intercalations
of massive and finely laminated
grey and yellowish limestones, bearing -
in some levels- Steinmanella sp., Thalassinoides
isp. and poorly preserved flattened
ammonoids. In the upper part there are
non-fossiliferous olive green shales, limestone
with small oysters, silty limestones
and black shales with Spitidiscus riccardii
Leanza and Wiedmann, 1992 (Aguirre-Urreta 2001), Protohemichenopus neuquensis
Camacho, 1953, Lucinidea indet.,
and callianassid crustaceans.


Figure 3: Stratigraphic section of the Mendoza Group in the Argentine slope of the Piuquenes Pass.
The fossils that Darwin collected in Q
and S (presently Lo Valdés Formation/
Mendoza Group) were studied by d'Orbigny
and cited by Darwin (1846, p. 181)
as follows:
"The fossils above alluded to in the black calcareous
shales are few in number, and are in an
imperfect condition; they consist, as named for me
by M. d'Orbigny, of
1. Ammonite, indeterminable, near to A. recticostatus, D'Orbig. Pal. Franc. (Neocomian formation).
2. Gryphæa, near to G. Couloni, (Neocomian formations of France and Neufchatel).
3. Natica, indeterminable.
4. Cyprina rostrata, D'Orbig. Pal. Franc. (Neocomian formation).
5. Rostellaria angulosa (?), D'Orbig. Pal. de l'Amer. Mer.
6. Terebratula ?"
However, it seems that d'Orbigny gave
Darwin slightly different information
(see fig. 4). According to his notes, the
specimens collected by Darwin in the "Cordillera Centrale du Chili" comprised:
"617/792. Gryphaea - Voisine du Gryphaea
Couloni, du terrain Neocomien de France et
de Neuchatel
613. Natica (indeterminable)
619. Ammonite indeterminable, voisine de l'A.
recticostatus, d'Orb (Paleont. française) Du
terrain Neocomien
614. B. Cyprina rostrata, d'Orb. Paleont.
franç. Terrain Neocomien (non Lucina)
C. Terebratula ?
A. Rostellaria angulosa, d'Orb. Paleont. de
l'Am. Mer. Pl. 18 fig. 4 ?"

Figure 4 : List of fossils identified by d'Orbigny for Darwin from his collection of the Central Cordillera
of Chile (from www.darwin-online.org.uk).
d'Orbigny also listed :
"790. Arca peut être Arca gabrielis, d' Orb.
(Paleontologie francaise) du terrain Neocomien" This fossil is not mentioned by Darwin
from the Piuquenes Pass, and is almost
certainly from Puente del Inca (Darwin 1846, p. 193), as this specimen and the
"792 (Gryphaea - Voisine du Gryphaea
Couloni)" have a note in the left margin
stating Cumbre, probably handwritten by Darwin (Fig. 4).
D'Orbigny assigned a bivalve to Cyprina
rostrata d'Orbigny, 1853 pointing out that
this is not Lucina Lamarck, 1799, which is
interesting as here this fossil is referred
to Lucinidae indet. (D. Lazo, oral comm.)
(Fig. 5, p-q). He also added the figure of
Rostellaria angulosa d'Orbigny, 1842 (d'Orbigny
1842, p. 80, pl. 18, fig. 4) which was
very helpful in the assignation of our
specimens to Protohemichenopus neuquensis
Camacho, 1953 (Fig. 5 o). It is worth
mentioning here that, besides the ammonites
that will be described in detail below,
we have also found chelipeds of
callianassid crustaceans in both the Vaca
Muerta and Agrio Formations.
Besides Darwin (1846) the other only published
mention of fossils in the Piuquenes
range is that of Polanski (1964, p.
48) who listed the following Tithonian
fossils: Corongoceras sp., Aulacosphinctes sp.,
Trigonia carricurensis Leanza, Lucina sp. and
Exogyra sp.

Figure 5: a-h) Pseudolissoceras zitteli (Burckhardt, 1903); a) lateral view CPBA 20552.1; b) lateral and ventral views CPBA 20552.5; c) lateral and ventral views CPBA
20552.4; d) lateral view CPBA 20552.3; e) lateral view CPBA 20552.2; f) lateral view CPBA 20552.6; g) apertural and lateral views CPBA 20552.7; h) lateral and ventral
views CPBA 20552.8; i-m) Virgatosphinctes aff. Virgatosphinctes denseplicatus rotundus Spath, 1931; i) lateral view CPBA 20551.3; j) lateral view CPBA 20551.4; k) ventral
and lateral views CPBA 20551.2; l) lateral view CPBA 20551.1; m) apertural and lateral views CPBA 20551.5; n) Spitidiscus riccardii Leanza and Wiedmann, 1992, lateral
view CPBA 20557; o) Protohemichenopus neuquensis Camacho, 1953, apertural and lateral views CPBA 20556.1; p-q) Lucinidae indet. lateral views CPBA 20555.1-2; r-t)
Aulacosphinctes proximus (Steuer, 1897); r) lateral view CPBA 20553.2; s) lateral and ventral views CPBA 20553.1; t) Parodontoceras calistoides (Behrendsen, 1891), lateral
view CPBA 20554. All specimens from the Piuquenes Pass, Mendoza. All natural size. Specimens coated with ammonium chloride.
SYSTEMATIC PALAEONTOLOGY
Family Haploceratidae Zittel, 1884
Genus Pseudolissoceras Spath, 1925
Pseudolissoceras zitteli (Burckhardt, 1903) Figs. 5 a-h
? 1897 Oppelia perlaevis Steuer, p. 73, pl. 6,
figs. 7-9
1900 Oppelia aff. perlaevis Steuer; Burckhardt,
p. 46, pl. 26, figs. 5, 6; pl. 29, fig. 2
1903 Neumayria zitteli Burckhardt, p. 55,
pl. 10, figs. 1-8
1907 Neumayria zitteli Burckhardt; Haupt,
p. 200, pl. 7, figs. 3a, b, 4a-c
? 1921 Oppelia perlaevis Steuer, p. 102, pl.
6, figs. 7-9
? 1921 Oppelia perglabra Steuer, p. 104, pl.
7, figs. 13-15
1925 Pseudolissoceras zitteli (Burckhardt);
Spath, p. 113 (Gen. nov.)
1926 Pseudolissoceras zitteli Burckhardt;
Krantz, p. 436, pl. 17, figs. 4, 5
1928 Pseudolissoceras zitteli Burckhardt;
Krantz, p. 18, pl. 1, fig. 6; pl. 4, figs. 9a, b 1931 Pseudolissoceras zitteli Burckhardt;
Weaver, p. 401, pl. 43, fig. 291
1942 Pseudolissoceras cf. Pseudolissoceras zitteli
(Burckhardt); Imlay, p. 1443, pl. 4,
figs. 1-4, 7, 8, 11, 12
1980 Pseudolissoceras zitteli (Burckhardt);
Leanza, p. 17, pl. 1, figs. 1a, b, 2a, b
2001 Pseudolissoceras zitteli (Burckhardt);
Parent, p. 23, figs. 3a, b, 5a, b, 7a-f
Material: 10 specimens (CPBA 20552.1- 20552.10) from the Piuquenes Pass, Mendoza.
Description: Well preserved, small to medium size specimens (largest specimen is 62.2 mm diameter); body chamber occupies at least half a whorl, usually laterally crushed. Shell discoidal, involute (U c. 21% of diameter) with deep umbilicus, rounded umbilical border and an abrupt umbilical slope on the outermost whorls while on the innermost it becomes gently inclined. Subelliptic transversal section with convex flanks converging on a rounded and slightly acuminated venter. Whorls always higher than wide, with maximum width near the middle of the flanks. Shell surface almost smooth. Only where test is preserved is it possible to distinguish extremely soft, falcoid ribs densely packed, that start at the base of the umbilical slope. No dimorphic sexual structures observed. Suture line not completely preserved.
Dimensions of specimens

Remarks: Specimens described above
can certainly be assigned to Pseudolissoceras
zitteli (Burckhardt 1903, p. 55, pl. 10,
figs. 1-8), and it is also possible that specimens
figured as Oppelia perlaevis Steuer
(1897, p. 73, pl. 6, figs. 7-9; 1921, Spanish
translation of 1897 edition, p. 102, pl. 6,
figs. 7-9) and Oppelia perglabra Steuer
(1921, p. 104, pl. 7, figs. 13-15) from the
lowermost Middle Tithonian of Mendoza,
belong also in the same species.
This is due to the fact that there is a
strong similarity in general form, ornamentation,
and suture line between them
and Burckhardt's original form. Nevertheless,
we can not confirm this until we
have the chance of comparing our specimens
with Steuer's holotypes, although
we can confidently assert that they do
not belong in Oppelia Waagen, 1869
because they show no trace of even a
feeble keel preserved on the illustrated
material.
Specimens described and figured by
Haupt (1907, p. 200, pl. 7, fig. 3a, b, 4ac),
Krantz (1926, p. 436, pl. 17, figs. 4, 5;
1928, p. 18, pl. 1, fig. 6; pl. 4, fig. 9a, b),
Weaver (1931, p. 401, pl. 43, fig. 291) and
Leanza (1980, p. 17, pl. 1, figs. 1a, b, 2a,
b), were all found at Cerro Lotena, Neuquén,
although some specimens have
also been recovered from Picún Leufú (Weaver), Rodeo Viejo, Bardas Blancas,
Arroyo Cienaguitas and from a locality
between Cajón del Burro and Río Choica,
in Mendoza (Krantz 1926). Specimens
studied by Parent (2001, p. 23, fig.
3a, b, 5a, b, 7a-f) are from Cañadón de los
Alazanes, a locality exposed at the Vaca
Muerta Range in Neuquén, while Imlay's material (1942, p. 1443, pl. 4, figs. 1-4, 7,
8, 11, 12) is from Cuba. In all cases specimens
are highly similar; there is a notable
concordance in U/D and H/W ratios
with only some minimum differences in
transversal section -which can be sometimes
more inflated or depressed- or in
having a more or less acuminate or rounded
venter. In all cases, specimens are
found in association with a similar fauna
composed mainly of other haploceratid
ammonoids, from which they can be
generally differentiated by the absence of
a marked groove on the flanks, as occurs
in Hildoglochiceras Spath, 1924, or its rounded
venter and lesser degree of involution
than in Parastreblites Donze and Enay,
1961.
On the other hand, many authors (Haupt
1907, Krantz 1926, Weaver 1931, Leanza
1980) distinguish Pseudolissoceras pseudoolithicum
(Haupt, 1907) recorded in the same
beds that contain Pseudolissoceras zitteli.
That species never exhibits an umbilical
edge, it is only moderately involute,
and its flanks are more inflated with
whorls as high as wide in transverse section.
Ocurrence: Lowermost Middle Tithonian. Pseudolissoceras zitteli Zone.
Family Perisphinctidae Steinmann, 1890 Subfamily Virgatosphinctinae Spath, 1923 a Genus Virgatosphinctes Uhlig, 1910
Virgatosphinctes aff. Virgatosphinctes denseplicatus rotundus Spath, 1931 Figs. 5 i-m
? 1890 Perisphinctes contiguus Catullo; Toucas,
p. 581, pl. 14, fig. 4
? 1900 Perisphinctes contiguus Catullo; Burckhardt,
p. 45, pl. 24, fig. 1
1903 Perisphinctes contiguus Catullo; Burckhardt,
p. 38, pl. 4, figs., 7-10
? 1931 Virgatosphinctes denseplicatus (waagen)
var. rotunda Spath, p. 532, pl. 96, figs.
3a-b; pl. 102, fig 4.
1954 Virgatosphinctes cf. denseplicatus (Waagen)
var. rotunda Spath; Indans, p. 106,
pl. 21, fig. 1
1972 Virgatosphinctes denseplicatus Waagen;
Fatmi, p. 346, pl. 8, figs. 5a, b
1980 Virgatosphinctes denseplicatus rotundus
Spath; Leanza, p. 31, pl. 2, figs. 2, 3
Material: 6 specimens (CPBA 20551.1-20551.6) from the Piuquenes Pass, Mendoza.
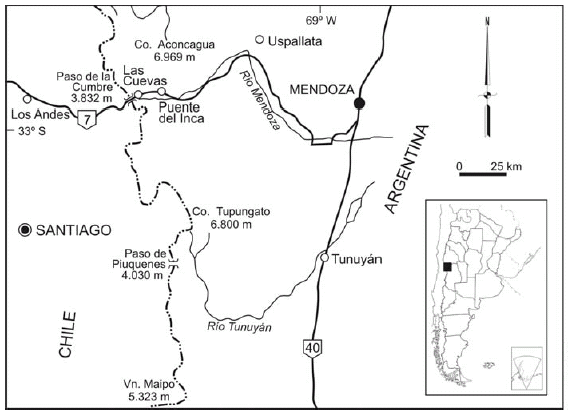
Description: Medium size discoidal phragmacone (largest specimen is 45.9 mm in diameter), body chamber not preserved. Slightly involute (U c. 33% of diameter), with moderately deep umbilicus and umbilical border not very inclined. Venter slightly planar with gently curved flanks. Whorl section somewhat inflated, slightly wider than high, with maximum width at umbilical margin. Fine, sharp ribs, regularly distributed, beginning at the middle of the umbilical slope, with a slightly prorsiradiate tendency and then bifurcate at the external third of the flank, crossing the venter without interruption. A slight weakness occurs in the ribs in specimens that are less than 38 mm in diameter. At least two shallow constrictions are observed in the last whorl preserved, parallel to the rib pattern and bordered posteriorly by a simple asymmetrical rib. Suture line not observed.
Dimensions of specimens
Remarks: Specific assignation of the
specimens to Virgatosphinctes denseplicatus
rotundus Spath (1931, p. 532, pl. 96, figs.
3a-b; pl. 102, fig. 4) is quite difficult due
to the fact that ornamentation suffers a
radical change in the last whorl, just as
illustrated by Waagen in Virgatosphinctes
denseplicatus (1873, p. 201, pl. 46, fig. 3a, b;
pl.55, figs. 1a. b), where the innermost
fine ribs become stronger over the inner
third of the last whorl, and then differentiate
in the middle of the flank into a
bundle of 4 to 10 secondary ribs that
cross the venter without any interruption.
It is important to state here that the
gradual differentiation from biplicate
ribs, into triplicate, virgatotome, and finally
fasciculate ribs that constitutes a
diagnostic character of Virgatosphinctes
Uhlig, 1910 is not evident in Waagen's
illustrated specimens from the Lower Tithonian
of Kutch.
Perisphinctes contiguus Catullo (in Toucas
1890, p. 581, pl. 14, fig. 4, Burckhardt
1900, p. 45, pl. 24, fig. 1, Burckhardt
1903, p. 38, pl. 4, figs. 7-10) closely resembles
the specimens described above,
with similar rib pattern and alternation of
triplicate ribs on one side with biplicate
ribs on the other; there is also a slight
attenuation of the ornamentation on the
venter of some specimens. Toucas' material
comes from the Lower Tithonian of
the Carpathians, the Alps, the Apennines
and from Andalucia, while Burkhardt's
specimens are from the Lower Tithonian
from Casa Pincheira in Mendoza.
Virgatosphinctes cf. denseplicatus (Waagen)
var. rotunda Spath (in Indans 1954, p. 106,
pl. 21, fig. 1) from Cerro Alto in southern
Mendoza also resembles the studied specimens,
as well as the specimen from
western Pakistan identified as Virgatosphinctes
denseplicatus Waagen by Fatmi
(1972, p. 346, pl. 8, figs. 5a, b). However,
there appears to be a difference in the
ornamentation pattern with the Argentinian
specimens, as there are simple posterior ribs bordering the constrictions,
not joining the immediately anterior
biplicate ribs.
Finally, there is a strong resemblance between
the specimens studied here and
Virgatosphinctes denseplicatus rotundus described
and figured by Leanza from the
Lower Tithonian of Cerro Lotena, Neuquén
(1980, p. 31, pl. 2, figs. 2, 3), though
his specimens show a more inclined umbilical
slope and a more subelliptical
whorl section.
Virgatosphinctes denseplicatus rotundus is
usually found in association with other
Virgatosphinctes, like Virgatosphinctes andesensis
(Burckhardt), from which it can be
differentiated due to its whorl section
only slightly wider than high, its softer
ornamentation and the lack of intercalatory
ribs and primary ribs describing an
inflection on the middle of the flanks.
Ocurrence: Basal section of Vaca Muerta Formation, just above the biolaminated carbonate levels. Lower Tithonian, Virgatosphinctes mendozanus Zone.
Family Perisphinctidae Steinmann, 1890 Subfamily Himalayitinae Spath, 1925 Genus Aulacosphinctes Uhlig, 1910
Aulacosphinctes proximus (Steuer, 1897) Figs. 5 r-t
1897 Reineckeia proxima Steuer, p. 34, pl. 8,
figs. 7-11
? 1897 Perisphinctes colubrinus Reinecke;
Steuer p. 62, pl. 15, fig. 11
1900 Perisphinctes colubrinus Reinecke;
Burckhardt, p. 44, pl. 24, figs. 5, 6; pl. 26,
fig. 4
1907 Perisphinctes proximus Steuer; Haupt,
p. 192
1921 Reineckeia proxima Steuer, p. 61, pl. 8,
figs. 7-11
? 1921 Perisphinctes colubrinus Reinecke;
Steuer, p. 90, pl. 15, fig. 11
1928 Aulacosphinctes wanneri Krantz, p. 42,
pl. 2, figs. 6a, b
? 1931 Aulacosphinctes proximus (Steuer);
Weaver, p. 411, pl. 44, figs. 298, 299
1931 Aulacosphinctes colubrinus (Reinecke);
Weaver, p. 413, pl. 44, figs. 301-303
1980 Aulacosphinctes proximus (Steuer);
Leanza, p. 44, pl. 6, figs. 2a, b, 4a, b, 5a, b
1981 Aulacosphinctes proximus (Steuer);
Leanza, p. 587, pl. 2, figs. 9, 10
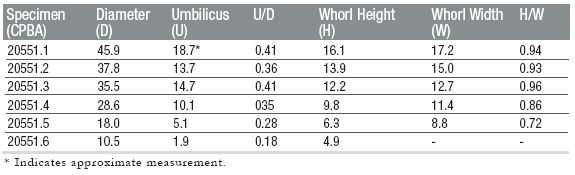
Material: 2 specimens (CPBA 20553.1- 20553.2) from the Piuquenes Pass, Mendoza.
Description: Medium size planulate
shell (largest specimen is 52.3 mm in diameter),
body chamber partially preserved,
of at least 1/4 whorl. Strongly evolute
(U c. 45% of diameter), with shallow
umbilicus, rounded umbilical border and
gradually inclined umbilical slope. Subquadrate
transversal section with slightly
convex flanks and flat venter, whorls
slightly higher than wide. Prominent, fine, sharp and distant ribs start at the
base of the umbilical slope in a rursiradiated
mode, at the umbilical margin they
become prorsiradiated, crossing the
flanks in a prorsiradiated to rectiradiated
pattern. Number of ribs decreases as diameter
increases; in a 52.3 mm diameter
specimen there are 25 ribs in the last
whorl and 20 in the preceding one. Ribs
on the innermost whorls are interrupted
over the venter by a narrow and shallow
furrow, which tends to disappear in outer
whorls although ornamentation there
may suffer an important weakening.
Simple primary ribs are irregularly intercalated
with biplicate ribs that bifurcate
at the middle of the flanks without developing
any tubercle at the bifurcation
point. Some intercalatory ribs are also
observed, not extending beyond the middle
part of the flanks. Two wide but shallow
constrictions occur near the shell
end; they are posteriorly bounded by a
simple, acute and prominent primary rib.
Suture line not visible.
Dimensions of specimens
Remarks: Diagnostic characters observed
in the specimens studied, such as
evolution degree, rib pattern and the presence
of a ventral furrow allow its assignation
to Aulacosphinctes proximus (Steuer
1897, p. 34, pl. 8, figs. 7-11 as Reineckeia
proxima; 1921 Spanish translation, p. 61,
pl. 8, figs. 7-11). However, in the same
publication Steuer describes another species,
from Arroyo Cienaguitas in Mendoza,
and from contiguous beds. This
species, (Perisphinctes colubrinus (Reinecke)
p. 62, pl. 15, fig. 11), is very similar to
Aulacosphinctes proximus (Steuer, 1897),
both in general form and ornamentation
pattern, but the ventral furrow is not visible
on the last whorl preserved, although
it is present on previous ones. Nevertheless
the specimen illustrated is 83 mm in
diameter, so this character may not be
noticeable at that size, although it might
be quite important in smaller shells as
can be clearly seen on the specimens studied
by Burckhardt (1900, p. 44, pl. 24,
figs. 5, 6; pl. 26, fig. 4) and Weaver (1931,
p. 413, pl. 44, figs. 301-303), from Mendoza
and Neuquén, respectively.
Weaver (1931, p. 411, pl. 44, figs. 298,
299) also described a specimen that was
assigned to Aulacosphinctes proximus. However,
his description does not mention
any rib interruption over the venter at
any shell size, an important character in
the definition of the genus that casts
doubts on its assignation.
Aulacosphinctes wanneri Krantz (1928, p.
42, pl. 2, figs. 6a, b) from Arroyo La
Manga, Mendoza, closely resembles Aulacosphinctes
proximus. The development of
a trifurcated rib is the only difference
between the two species, an unimportant
character that may be considered as
intraspecific variation.
Finally, the studied specimens are comparable
with those studied by Leanza
(1980, p. 44, pl. 6, figs. 2a, b, 4a, b, 5a, b; 1981, p. 587, pl. 2, figs., 9, 10) from the
Middle Tithonian of Cerro Lotena in
Neuquén.
Ocurrence: Middle Tithonian, Aulacosphinctes proximus Zone.
Family Neocomitidae Salfeld, 1921
Subfamily Berriasellinae Spath, 1922
Genus Parodontoceras Spath, 1923 a
Parodontoceras calistoides (Behrendsen, 1891) Fig. 5 t
1891 Hoplites calistoides Behrendsen, p.
402, pl. 23, figs. 1a, b
1897 Odontoceras calistoides Behrendsen;
Steuer, p. 41, pl. 17, figs. 13-16
1921 Odontoceras calistoides Behrendsen;
Steuer, p. 69, pl. 17, figs. 13-16
1922 Hoplites calistoides Behrendsen, p.
184, pl. 1, figs. 11a, b
1923a Parodontoceras callistoides Behrendsen;
Spath, p. 305 (Gen. nov.)
1925 Berriasella calistoides Behrendsen;
Gerth, p. 88
1928 Berriasella (Parodontoceras) calistoides
(Behrendsen); Krantz, p. 24
1945 Parodontoceras calistoides (Behrendsen);
Leanza, p. 41, pl. 5, figs. 5, 6
1988 Paradontoceras calistoides (Behrendsen);
Riccardi, pl. 3, figs. 3, 4
Material: 1 specimen (CPBA 20554) from the Piuquenes Pass, Mendoza.
Description: Medium to large phragmacone (97.2 mm diameter), laterally crushed. Shell ellipticone, involute (U c. 22 % of diameter), with poorly developed umbilical border and quite inclined umbilical slope. Flanks are gently convex and venter somewhat flattened. Whorls are higher than wide, with the maximum width probably located at the umbilical border. The whorl height increases considerably with diameter, resulting in an eccentric shaped shell. Ornamentation consists of moderately strong ribs, regularly spaced, that begin at least from the middle part of the umbilical slope in a rursiradiate manner, then bend forwards at the umbilical border describing an inflection, and then another inflection at the bifurcation point in the middle of the flanks. These biplicate ribs are intercalated with simple ribs and some intercalatory ones do not go beyond the external third of the flank. All ribs are interrupted over the venter by a relatively wide and shallow furrow, which becomes less visible as diameter increases until it disappears completely. Suture line is very badly preserved.
Dimensions of specimen

Remarks: Specific assignation of the specimen to Parodontoceras calistoides (Behrendsen, 1891, p. 402, pl. 23, figs. 1a, b) is quite safe despite its rather poor preservation, especially taking into account its eccentric shape, rib pattern and ventral furrow. This species is known from many localities in Neuquén and Mendoza, almost always in association with Substeueroceras koeneni (Steuer, 1897) characterized by a finer and denser rib pattern than Parodontoceras calistoides. Nevertheless, they seem to be very close forms and only a future more detailed study will prove if they are not just morphotypes of the same species.
Ocurrence: Uppermost Tithonian, Substeueroceras
koeneni Zone.
Family Holcodiscidae Spath, 1923 b
Genus Spitidiscus Kilian, 1910
Spitidiscus riccardii Leanza and Wiedmann, 1992 Fig. 5 n
1992 Spitidiscus riccardii Leanza and Wiedmann,
p. 33, figs. 4a-b
1992 Spitidiscus aff. S. rotula (J. de C. Sowerby);
Leanza and Wiedmann, p. 32, figs.
3a-b
1992 Spitidiscus aff. S. gastaldianus (d'Orbigny);
Leanza and Wiedmann, p. 32,
figs. 6a-b
1995 Spitidiscus riccardii Leanza and Wiedmann;
Aguirre-Urreta, p. 407, pl. 1, figs.
1-23
1999 Spitidiscus riccardii Leanza and
Wiedmann; Aguirre-Urreta et al., pl. 1,
fig. 4.
2005 Spitidiscus riccardii Leanza and Wiedmann;
Aguirre-Urreta et al., figs. 7 e-f
Material: 1 specimen (CPBA 20557) and several fragments and impressions from the Piuquenes Pass, Mendoza.
Description: Shell small, involute (approx. diameter 30 mm), with rounded umbilical slope and slightly rounded flanks converging towards a curved venter. Ornament is composed by ribs and constrictions. The ribs are fine, arise from the umbilical slope and bifurcate in the lower third of the flank where they curve into a falcoid shape. They cross the venter without interruption bending slightly towards the aperture. The constrictions are less curved than the ribs and they dissect them. Suture line not preserved.
Remarks: Although the preservation of the specimens is rather poor, they can be assigned to Spitidiscus riccardii Leanza and Wiedmann, 1992 due to the size, involution of the shell and ribbing pattern. The studied specimens are comparable with those studied by Aguirre-Urreta (1995, p. 407, pl. 1, figs. 1-23) from Agua de la Mula and Agrio del Medio in Neuquén where the species has been studied in detail.
Occurrence: Upper Hauterivian. Spitidiscus riccardii Zone.
CONCLUDING REMARKS
When one compares the geological section of the Piuquenes Pass by Darwin
(1846, pl. 1, skecth 1) with the present
knowledge of the area (Pángaro et al.
1996, Fig. 2 herein), it can be seen that
Darwin was able to clearly identify the
succession of Meso-Cenozoic rocks and
some structural features. He only partially
recognized the highly complex
structure of thrusts that affects the sedimentary
rocks, repeating parts of the
sequence.
Regarding the fossil content, it must be
understood that Darwin just rode along
the trail, suffering the effects of high altitude
and that he was being hurried by the "baqueanos" who were afraid of bad weather.
He wrote to Henslow: "It was late in
the Season, & the situation particularly dangerous
for Snow storms. I did not dare to delay,
otherwise a grand harvest might have been reaped" (Burkhardt and Smith 1985). He was
able to collect the most common fossil,
his big oyster Exogyra (Gryphacea) couloni
(presently Aetostreon), other bivalves, some
gastropods and ammonites. It is difficult
to know precisely from where he got
his specimens, though it seems he stopped
several times along the trail collecting
loose material, a fact that would explain
why the fossils were poorly preserved.
D'Orbigny (manuscript, see Fig. 4, and
also Darwin 1846) regarded the assemblage
as Neocomian age, based not only
in the oysters but also in the presence of
an indeterminate ammonite resembling
Ammonites recticostatus d'Orbigny, 1840.
This species is presently assigned to the
genus Macroscaphites Meek, 1876 of Barremian-Early Aptian age, and is completely
unknown in the Neuquén Basin,
where the youngest ammonites are of
Early Barremian age (Aguirre-Urreta et
al. 2005). Most probably d'Orbigny confused
it with some Tithonian perisphinctids
which are common in the area.
In spite of the poor preservation of the
specimens and the lack of knowledge of
faunas of this age in South America, it
should be emphasized that Darwin was
correct in assigning the whole succession
to the Neocomian, following d'Orbigny's
advice. This fauna was one of the first to
be documented in the High Cordillera of
the Main Andes, which now we know
that also includes the Tithonian as seen
by the ammonites described above.
ACKNOWLEDGEMENTS
The authors wish to acknowledge Dario Lazo for his information on the bivalves and Victor A. Ramos for his assistance in the field and his advice on the structure of the region. The reviewers Estanislao Godoy, Héctor Leanza and Amaro Mourgues improved the original manuscript with their comments.
WORKS CITED IN THE TEXT
1. Aguirre-Urreta, M.B. 1995. Spitidiscus riccardii Leanza y Wiedmann (Ammonoidea) en el Hauteriviano del Neuquén. Ameghiniana 32: 407-410.
2. Aguirre-Urreta, M.B. 1996. El Tithoniano marino de la vertiente argentina del paso de Piuquenes, Mendoza. 13° Congreso Geológico Argentino y 3° Congreso de Exploración de Hidrocarburos (Buenos Aires), Actas 2: 185.
3. Aguirre-Urreta, M.B. 2001. Marine Upper Jurassic-Lower Cretaceous Stratigraphy and Biostratigraphy of the Aconcagua-Neuquén Basin, Argentina and Chile. Journal of Iberian Geology 27: 71-90.
4. Aguirre-Urreta, M.B., Concheyro, A., Lorenzo, M., Ottone, E.G. and Rawson, P.F. 1999. Advances in the biostratigraphy of the Agrio Formation (Lower Cretaceous) of the Neuquén basin, Argentina: ammonites, palynomorphs, and calcareous nannofossils. Paleogeography, Palaeoclimatology, Palaeoecology 150: 33-47.
5. Aguirre-Urreta, M.B., Rawson, P.F., Concheyro, G.A., Bown, P.R. and Ottone, E.G. 2005. Lower Cretaceous Biostratigraphy of the Neuquén Basin. In Veiga, G., Spalletti, L., Howell, J.A. and Schwarz, E. (eds.) The Neuquén Basin: A case study in sequence stratigraphy and basin dynamics. The Geological Society, Special Publication 252: 57-81, London.
6. Behrendsen, O. 1891-1892. Zur Geologie des Ostabhanges der argentinischen Kordillere. Deutsche Geologische Gesellschaft 43 (1891): 369-420; 44 (1892): 1-42.
7. Behrendsen, O. 1922. Contribución a la geología de la pendiente oriental de la Cordillera Argentina. Academia Nacional de Ciencias, Actas 7: 157-227, Córdoba.
8. Burckhardt, C. 1900. Profils géologiques transversaux de la Cordillère Argentino-Chilienne. Museo de La Plata, Sección Geológica y Mineralógica, Anales 2: 1-136.
9. Burckhardt, C. 1903. Beitrage zur Kenntniss der Jura und Kreide formation der Cordillere. Palaeontographica 50: 1-144.
10. Burkhardt, F. and Smith, S. 1985. The Complete Correspondence of Charles Darwin. Vol. 1. 1821-1836. Cambridge University Press, 752 p., Cambridge.
11. Camacho, H. H. 1953. Algunas consideraciones sobre los "Aporrhaidae" fósiles argentinos. Revista de la Asociación Geológica argentina 8: 183- 194.
12. Darwin, C. 1846. Geological observations on South America, being the third part of the geology of the voyage of the Beagle, under the command of Capt. FirtzRoy, RN, during the years 1832 to 1836. Smith, Elder and co., 268 p. London.
13. d'Orbigny, A. 1840-1841. Paléontologie Francaise. Description des Mollusques et Rayonnés Fossiles. Terrains Crétacés. Masson, 662 p., Paris.
14. d'Orbigny, A. 1842. Voyage dans L'Amérique Méridionale. Tome 3, 4° part: Paleontologie, 188 p., Paris & Strasbourg.
15. Fatmi, A.N. 1972. Stratigraphy of the Jurassic and Lower Cretaceous rocks and Jurassic ammonites from northern areas of West Pakistan. British Museum of Natural History 20: 297-380.
16. Gerth, E. 1925. La fauna Neocomiana de la Cordillera argentina, en la parte meridional de la provincia de Mendoza. Academia Nacional de Ciencias, Actas 9: 57-132, Córdoba.
17. Giambiagi, L., Tunik, M., Ramo, V. A. and Godoy, E. 2009. The High Andean Cordillera of Central Argentina and Chile along the Piuquenes Pass- Cordón del Portillo transect: Darwin´s pioueering observations compared with modern geology. Revista de la Asociación Geológica Argentina 64 (1): 44-54.
18. Groeber, P. 1953. Mesozoico. In Geografía de la República Argentina, Sociedad Argentina de Estudios Geográficos GAEA 2(1): 9-541, Buenos Aires
19. Haupt, O. 1907. Beiträge zur Fauna des oberen Malm und der unteren Kreide in der argentinischen Cordillere. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 23: 187-236.
20. Imlay, R.W. 1942. Late Jurassic fossils from Cuba and their economic significance. Geological Society of America, Bulletin 53: 1417-1478.
21. Indans, J. 1954. Eine Ammonitenfauna aus dem Untertithon der argentinischen Kordillere in Süd-Mendoza. Palaeontographica 105 A: 96- 132.
22. Jordan, T., Isacks, B., Ramos, V.A. and Allmendinger, R.W. 1983. Mountain building model: The Central Andes. Episodes 1983(3): 20-26.
23. Kilian, W. 1910. Erste Abteilung: Unterkreide (Palaeocretacicum). Lieferung 2: Das bathyale Palaeocretacicum im südostlichen Frankreich; Valendis-Stufe; Hauterive-Stufe; Barreme-Stufe; Apt-Stufe. In Frech, F., Lethaea Geognostica. II. Das Mesozoicum, Band 3 (Kreide) (1907-1913), Schweizerbart, 169-288, pls. 1-8, Stuttgart.
24. Krantz, F. 1926. Die ammoniten des Mittel-und Obertithons. In Jaworski, E., Krantz, F. and Gerth, E. (eds.) Beiträge zur Paläontologie und Stratigraphie des Lias, Doggers, Tithons und der Unterkreide in der Kordillere im Süden der Provinz Mendoza (Argentinien). Geologische Rundschau 17a: 427-462.
25. Krantz, F. 1928. La fauna del Titono superior y medio en la parte meridional de la provincia de Mendoza. Academia Nacional de Ciencias, Actas 10: 1-57, Córdoba.
26. Leanza, A.F. 1945. Ammonites del Jurásico Superior y del Cretácico Inferior de la Sierra Azul, en la parte meridional de la provincia de Mendoza. Anales del Museo de La Plata NS 1: 1-99.
27. Leanza, H.A. 1980. The Lower and Middle Tithonian Ammonite Fauna from Cerro Lotena, Province of Neuquén, Argentina. Zitteliana 5: 3-49.
28. Leanza, H.A. 1981. Faunas de ammonites del Jurásico superior y del Cretácico inferior de América del Sur, con especial consideración de la Argentina. In Volkheimer, W. and Musacchio, E.A. (eds.) Cuencas sedimentarias del Jurásico y Cretácico de América del Sur. Comité Sudamericano del Jurásico y Cretácico 2: 559-597, Buenos Aires.
29. Leanza, H.A. and Wiedmann, J. 1992. Nuevos Holcodiscidae (Cephalopoda-Ammonoidea) del Barremiano de la Cuenca Neuquina, Argentina y su significado estratigráfico. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1992, 1: 24-38.
30. Malumián, N. and Ramos, V.A. 1984. Magmatic intervals, transgression regression cycles and oceanic events in the Cretaceous and Tertiary of Southern South America. Earth and Planetary Science Letters 67: 228 237.
31. Pángaro, F., Ramos, V.A. and Godoy, E. 1996. La Faja plegada y corrida de la Cordillera Principal de Argentina y Chile a la latitud del Cerro Palomares (33°35'S). 13° Congreso Geológico Argentino y 3° Congreso Exploración de Hidrocarburos (Buenos Aires), Actas 2: 315-324.
32. Parent, H. 2001. The Middle Tithonian (Upper Jurassic) ammonoid fauna of Cañadón de los Alazanes, southern Neuquén-Mendoza Basin, Argentina. Boletín del Instituto de Fisiografía y Geología 71: 19-38, Rosario.
33. Polanski, J. 1964. Descripción geológica de la Hoja 25a Volcán San José, provincia de Mendoza. Dirección Nacional de Geología y Minería, Boletín 98: 1-94, Buenos Aires.
34. Riccardi, A. 1983. The Jurassic of Argentina and Chile. In Moullade, M. and Nairn, A.E.M. (eds.) The Phanerozoic geology of the world II, The Mesozoic B, Elsevier, 201-263, Amsterdam.
35. Riccardi, A.C. 1988. The Cretaceous System of southern South America. Geological Society of America, Memoir 168: 1-161.
36. Salfeld, H. 1921. Kiel- und Furschenbildung auf der Schalenaussenseite der Ammonoifrrn in ihrer Bedeutung für die Systematik und Festlegung von Biozonen. Zentralblatt für Mineralogie, Geologie und Paläontologie 1921: 343-347.
37. Spath, L.F. 1922. On Cretaceous Ammonoidea from Angola, collected by Professor J.W. Gregory, D. Sc., F.R.S. Transactions of the Royal Society of Edinburgh 53: 91-160.
38. Spath, L.F. 1923 a. On ammonites from New Zealand. In Trechmann, Ch.T. (ed.) The Jurassic Rocks of New Zealand. Geological Society of London, Quarterly Journal 79: 246-312.
39. Spath, L.F. 1923 b. A monograph of the Ammonoidea of the Gault. Part 1. Palaeontographical Society, Monographs 75 (353): 1-72.
40. Spath, L.F. 1925. Ammonites and Aptychi. In Wyllie, B.N.K and Smellie, W.E. (eds.) The collection of fossils and rocks from Somaliland. Hunterian Museum, Glasgow University, Monograph 7: 111-164.
41. Spath, L.F. 1927-1933. Revision of the Jurassic Cephalopod Fauna of Kachh (Cutch). Paleontologia Indica, NS 9, 1 (1927): 1-72, 2 (1928): 73-161, 3 (1928): 163-278, 4 (1931): 279-550, 5 (1931): 551-658, 6 (1933): 659-945.
42. Steinmann, G. 1890. Cephalopoda. In G. Steinmann and L. Döderlein (eds.) Elemente der Paläontologie, 344-475, W. Engelmann, Leipzig.
43. Steuer, A. 1897. Argentinische Jura - Ablagerungen: Eine Beitrage zur Kenntniss der Geologie und Paleontologie der argentinischen Anden. Palaeontologische Abhandlungen NS 7: 127-222, Jena.
44. Steuer, A. 1921. Estratos Jurásicos Argentinos. Contribución al conocimiento de la Geología y Paleontología de los Andes Argentinos entre el Río Grande y el Río Atuel. Academia Nacional de Ciencias, Actas 7: 5-128, Córdoba.
45. Toucas, A. 1890. Étude sur la fauna des couches tihoniques de l'Ardèche. Bulletin de la Société Géologique de France (Serie 3) 18: 560-629.
46. Uhlig, V. 1910. The Fauna of the Spiti Shales. Palaeontologia Indica 15: 133-395.
47. Waagen, W. 1873-1875. Jurassic Fauna of Kutch. Palaeontologia Indica 1 (1873): 1-22, 2 (1875): 23-76, 3 (1875), 77-106, 4 (1875): 107-247.
48. Weaver, C. 1931. Paleontology of the Jurassic and Cretaceous of West Central Argentina. Memoirs of the University of Washington 1: 1-496.
49. Zittel, K.A. 1884. Cephalopoda. In Zittel, K.A. Handbuch der Palaeontologie, R.Oldenbourg, Band 1, Abteilung 2, Lieferung 3, 893 p., Munich and Leipzig..
50. Zittel, K.A. 1895. Grundzüge der Palaeontologie. R. Oldenbourg, viii + 971 p., 2.048 figs, Munich and Leipzig.
Recibido: 10 de septiembre de 2008
Aceptado: 4 de noviembre de 2008