











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Los humedales cubren el 6 % de la su perficie terrestre del mundo y contienen aproximadamente el 12 % del depósito de carbono global, desempeñando un papel importante en el ciclo global del carbo no (Erwin, 2009; Ramsar, 2016). Estos ecosistemas se encuentran entre los más productivos del mundo, son ambientes de alta diversidad biológica, proporcio nando agua, productividad primaria y servicios ecosistémicos, e innumerables especies dependen de ellos para sobrevi vir (Malvárez & Bó, 2004; Laterra et. al, 2011; Ramsar, 2016).
El sitio Ramsar Lagunas de Guanaca che, Desaguadero y del Bebedero es un importante humedal de la región cuyana inserto en la diagonal árida de Argenti na (Secretaría de Ambiente y Desarrollo Sustentable, 2010; Martínez Carretero, 2013). Este sistema ha sufrido un con tinuo proceso de desecamiento debido a causas naturales y antrópicas que han actuado en forma aislada o en conjunto (Torres, 2015).
Las lagunas temporarias o semiper manentes son sistemas acuáticos que se caracterizan por alternar fases de inun dación y desecación que condicionan el desarrollo de las comunidades acuáticas que las habitan (Bronmark & Hansson 2005; Díaz-Paniagua et al., 2011). Du rante muchos años, estos ambientes han sido infravalorados y han tenido escasa o nula protección (Díaz-Paniagua et al., 2011). El objetivo del presente trabajo es caracterizar el ensamble de aves de un ambiente lagunar temporario del sitio Ramsar Lagunas de Guanacache, Des aguadero y del Bebedero de acuerdo a su diversidad taxonómica y funcional (gremios tróficos), abundancia específi ca, estatus de residencia y estado de conservación.
Material y método
Área de estudio
El Humedal Desaguadero del Bermejo se encuentra ubicado en el departamen to 25 de Mayo, provincia de San Juan, a 562 m s.n.m. (coordenadas de referen cia: 32° 12’ 21,1’’ S - 67° 36’ 16,4’’ O). Se halla aproximadamente a 135 km al su reste de la ciudad capital, a 14 km de la localidad El Encón y se accede por ruta nacional No 20.
Constituye un sistema de lagunas y bañados encadenados, alimentados por el río San Juan y esporádicamente por los desagües del Bermejo (Figura 1).

Figura 1: Mapa de localización del área de estudio y cuadrantes de muestreo. Figure 1: Location map of the study area and sampling quadrants.
Este humedal alcanza una extensión de 305.000 hectáreas aproximadamente, pertenece a sitio RAMSAR (Humedal de Importancia Mundial). El 5 de junio de 2007 se amplió el sitio Lagunas de Guanacache, pasando a denominarse “Lagunas de Guanacache, Desaguadero y Del Bebedero”, incluyendo 962.370 ha de las provincias de Mendoza, San Juan y San Luis. En este humedal, las aves son el grupo más abundante y uno de los componentes faunísticos principales que justifican la declaración del área como Sitio Ramsar (Secretaría de Ambiente y Desarrollo Sustentable, 2010). El sector de estudio corresponde a un ambiente lagunar temporario del sistema, locali zado a ambos lados de la ruta nacional N° 20. Este se forma algunos años como consecuencia de mayores caudales de los ríos San Juan y Mendoza, como también de agua meteórica de precipitaciones lo cales.
Diseño de muestreo
El trabajo de campo consistió en un muestreo por mes (de 2 días consecuti vos) durante 4 meses (época reproduc tiva), desde enero hasta abril de 2008. El método de muestreo seleccionado para llevar a cabo el estudio consistió en re levamientos por registros visuales por medio de puntos de observación (Bibby et al., 1998; Codesio, 2000). Se realizó un barrido con un radio variable, de terminado por la distancia a la cual se detectaron las aves, siguiendo el criterio de visibilidad (Bibby et al., 1998). Para minimizar los errores por sobreestima ción (Tellería, 1986) se dividió el área de estudio en 4 cuadrantes, cada uno con un punto de observación sobre la ruta nacional N°20 debido a la imposibilidad de un recorrido interno. Los cuadrantes I y II (en dirección SO y SE respectiva mente), con vista hacia el sur; los cua drantes III y IV (en dirección NE y NO respectivamente), con vista hacia el nor te (Figura 1). Los puntos de observación estuvieron separados aproximadamente 10 metros de distancia entre sí. Las aves contactadas en la franja imaginaria (de 10 metros) que se formó en la delimita ción de los cuadrantes no se tuvieron en cuenta para el muestreo.
El avistaje se realizó de forma orde nada: fue sistemático y en sentido anti-horario. Luego de realizar el barrido del cuadrante I de derecha hacia el centro, se caminó el ancho de la faja para llevar a cabo la observación en el cuadrante II del centro hacia la izquierda. Después de esto, se cruzó la ruta y se registraron los dos cuadrantes restantes. El cuadrante III, de espaldas al cuadrante II, de dere cha al centro; posteriormente se cami nó el ancho de la faja para llegar al IV cuadrante y realizar el barrido del centro hacia la izquierda. El tiempo de observa ción por cuadrante fue de 10 minutos. Todos los individuos que se encontraron dentro de los mismos fueron contados y todas las observaciones fueron inde pendientes. Los datos fueron tomados siempre por el mismo observador, para evitar el error existente entre diferentes observadores (Tellería, 1986). Fuera de los horarios de muestreo, se realizaron observaciones asistemáticas para rele vamiento de especies. Con la finalidad de poder detectar la mayor cantidad de especies en función de la variación de ac tividad diaria de las diversas especies, se realizaron muestreos a las 8, 10, 12, 14 y 16 horas, cubriendo casi la totalidad del tiempo de actividad de las aves del hu medal.
Análisis de datos
Composición taxonómica
Se identificaron todas las aves cotejando con la guía de campo de Narosky & Yzu rieta (2010) y se siguió la clasificación utilizada por el Ministerio de Ambiente y Desarrollo Sustentable y Aves Argenti nas (2017) que incorpora recientes arre glos sistemáticos.
Ensamble trófico
Para la selección de los gremios tróficos se siguió a Narosky & Izurieta (2010) y Rodriguez Matta et al. (2008). Se consi deraron las siguientes categorías: carro ñeros (C), zoófagos (Z), granívoros (G), herbívoros (H), insectívoros (I), piscívo ros (PI) y omnívoros (O).
Estado de residencia
Se clasificaron las especies tomando como base referencias bibliográficas. Se gún Narosky & Izurieta (2010), se con sideran residentes (R), migradoras A (MA), migradoras B (MB) y migradoras C (MC).
Conservación de especies
Se utilizó la última categorización de aves del Ministerio de Ambiente y De sarrollo Sustentable y Aves Argentinas (2017), donde se indica el estado de con servación de cada especie.
Proporción de individuos para cada especie
Se calculó dividiendo el número de in dividuos registrados de la especie i en el número total de individuos, multiplica do por 100 para obtener el porcentaje (pi = ni / ΣnT , donde pi representa la abun dancia relativa de la especie i, ni el nú mero de individuos de la especie i, ΣnT el número total de individuos) (Pettingil, 1969).
La dominancia: d= pi máx., donde pi máx. representa la proporción de la es pecie i que con mayor cantidad de indi viduos contribuyó a la abundancia total (May, 1975; Torres, 1995; Ordano, 1996). Las especies subordinadas deben tener un pi ≥ 0,05 para diferenciarlas de las raras.
La riqueza específica: S, es el número total de especies registradas durante el muestreo o estudio (Soave et al., 1999; Moreno, 2001).
Diversidad específica
Se determinó mediante el Índice de Di versidad de Simpson: D = 1 / Σ [ni (ni-1) / N (N-1)] = 1 / Σ (pi2), donde pi es la proporción de los individuos pertenecientes a la especie i: pi= ni / NT, donde ni es el número de individuos pertenecientes a la especie i y NT corresponde al número total de individuos (May, 1975). El Índice de Di versidad de Simpson (D) varía entre 1 (para una muestra con una sola especie) y S (riqueza) cuando todas las especies tienen exactamente el mismo número de individuos (Begon et al., 1999; Feinsin ger, 2001).
La equitatividad se cuantificó en base al Índice de Diversidad de Simpson (D), como una proporción del máximo valor que podría asumir D si los individuos estuvieran distribuidos de modo total mente uniforme entre las especies. En este caso, la diversidad máxima resul taría igual a la riqueza (Dmax= S). Por consiguiente: E= D / Dmax= [1 / Σ (pi2)] x (1 / S). La equitatividad adopta un va lor comprendido entre 0 y 1 (Begon et al., 1999).
Resultados
Composición taxonómica
Se registró un total de 45 especies de aves (39 especies en los muestreos siste máticos y 6 especies fuera de ellos). Estas especies pertenecen a 34 géneros, 20 fa milias y 11 órdenes. La composición del ensamble estuvo dominada, en términos de riqueza, por la familia Anatidae con 14 especies. Le siguieron en importancia las familias Podicipedidae y Ardeidae con 4 especies cada una (Tabla 1).
Tabla 1: Especies de aves relevadas en el humedal durante el estudio. Se indican con asterisco (*) las aves registradas fuera de los muestreos siste máticos. Se indica también Abundancia absoluta y relativa, Gremios tróficos (I = insectívoros, PI = piscívoros, O = omnívoros, H = herbívoros, C = carroñeros, Z = zoófagos, G = granívoro), estatus de residencia (MA = migrador A, MC = migrador C, R = residente) y categoría de conservación (EC = En peligro crítico EN = En peligro, AM = Amenazada, VU = Vulnerable, NA = No amenazada, IC = Insuficientemente conocida). Table 1: Bird species surveyed in the wetland during the study. Birds recorded outside of systematic sampling are indicated with an asterisk (*). Absolute and relative abundance are also indicated, trophic guilds (I = insectivores, PI = piscivores, O = omnivores, H = herbivores, C = scavengers, Z = zoophagus, G = granivore) and residence status (MA = migrant A, MC = migrant C, R = resident) and and conservation category (EC = Critically endangered EN = Endangered, AM = Threatened, VU = Vulnerable, NA = Not threatened, IC = Insufficiently known).
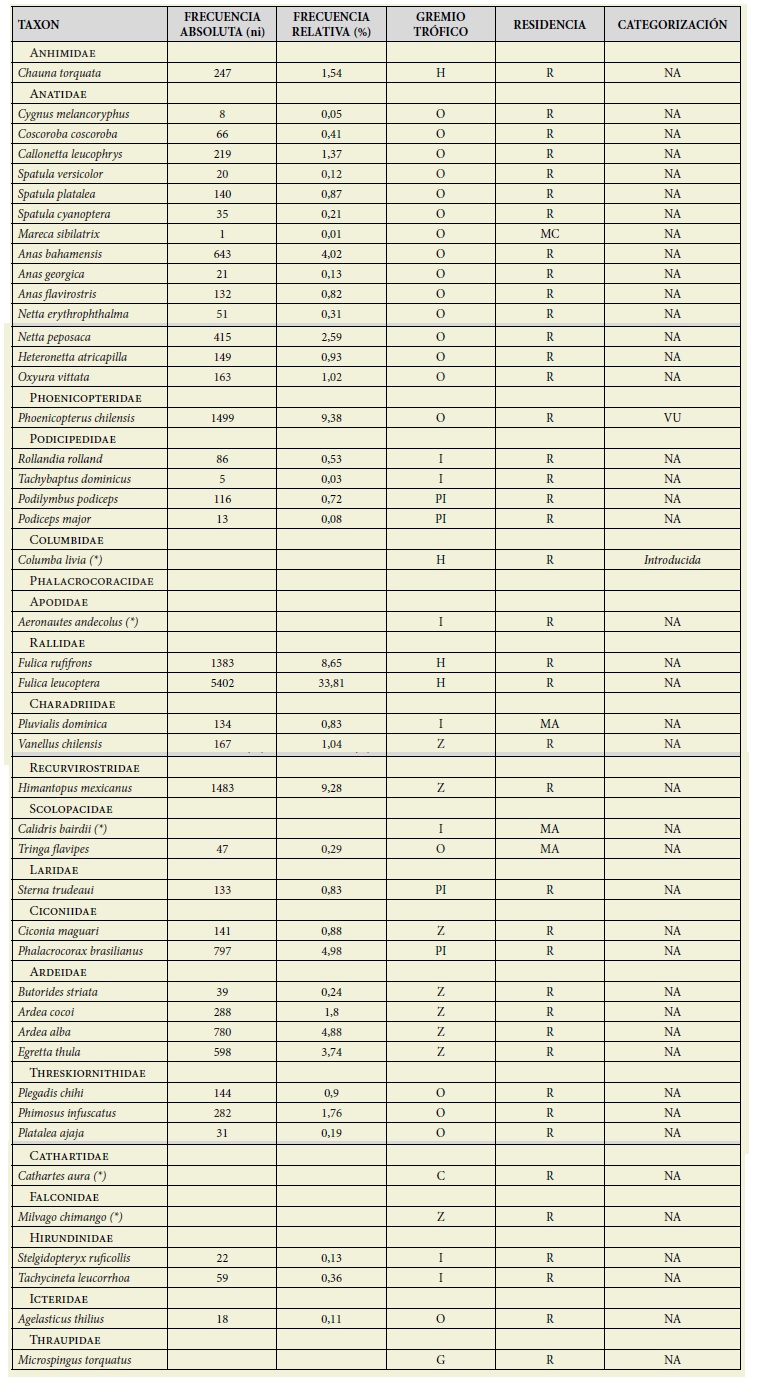
Para citar las especies de aves se siguieron la listas taxonómica propuesta por el Comité de Clasificación de América del Sur (Remsen et al., 2021), considerando los más recientes cambios.
Bibliografía: Remsen, J. V., Jr., J. I. Areta, E. Bonaccorso, S. Claramunt, A. Jaramillo, D. F. Lane, J. F. Pacheco, M. B. Robbins, F. G. Stiles, and K. J. Zimmer. 2021. A classification of the bird species of South America. American Ornithological Society. http://www.museum.lsu.edu/~Remsen/SA CCBaseline.htm [SACC Classification Version 19 January 2021]
Representación de los ensambles tróficos y estatus de residencia
El ensamble estuvo representado mayo ritariamente por aves omnívoras con un 47 % (21 especies); le siguieron en im portancia las aves zoófagas con un 38 % (17 especies); el 15 % restante son insec tívoras (7 especies) (Tabla 1).
El 91,1 % de las especies fueron residen tes y el resto, migratorias (6,67 % migrato rias A y 2,22 % migratorias C) (Tabla 1).
Especies dominantes, subordinadas y raras
Fulica leucoptera fue la especie dominan te, d, con un pi máximo= 0,34. Contribu yó así con la mayor cantidad de indivi duos a la abundancia total. Las especies subordinadas fueron Phoenicopterus chilensis, Himantopus mexicanus y Fulica rufifrons; mientras que las especies res tantes se clasificaron como raras.
Estado de conservación
El 95 % de las especies (43) se encuen tran en la categoría NA (no amenazada) a nivel nacional. Phoenicopterus chilensis es la única especie registrada categoriza da como vulnerable (Tabla 1).
Diversidad, equitatividad, riqueza
La diversidad fue relativamente baja (D=6,68) comparada con la diversidad máxima esperada Dmax=S, que en este caso sería 39. La equitatividad también resultó relativamente baja (E=0,17). La curva de dominancia-diversidad se ase meja al modelo de la serie logarítmica (Figura 2).

Discusión
La riqueza de especies observada (45 especies) fue mucho menor a la infor mada para otras lagunas y bañados del sitio Ramsar; Lucero (2013) informa un registro total de 205 especies para el sis tema de laguna del Toro, laguna de Gua nacache, bañados del Carau, laguna Seca y bañados del Tulumaya. En cada uno de estos cinco humedales se contabilizó una riqueza de más 110 especies (Lucero, 2013). La baja riqueza observada proba blemente esté altamente condicionada por los períodos de desecamiento pro longados en el tiempo que impiden un desarrollo de vegetación palustre y por consiguiente generación de hábitat y re cursos para otras especies. La baja diver sidad, baja equitatividad y el patrón de dominancia-diversidad observados pre dicen un número pequeño de especies abundantes y una gran proporción de especies raras, lo que es aplicable cuando uno o unos pocos factores dominan la estructura de la comunidad, como en las etapas iniciales de la sucesión (Moreno, 2001). La riqueza funcional se presentó simplificada con dominancia de especies omnívoras y zoófagas, probablemente esto también como consecuencia de ser un sistema en etapa inicial de sucesión por el reciente ingreso de agua al sistema después de un periodo de desecación. Algunos estudios de diversidad de aves para otros ecosistemas en diversos esta díos sucesionales sugieren que un incre mento en la estructura de la vegetación (riqueza y estructura vertical) favorecen el incremento de la diversidad funcional del ensamble (Salas-Correa & Mancera- Rodríguez, 2020).
La mayoría de las especies (95 %) se encuentran categorizadas como espe cies no amenazadas, solo encontrándose como vulnerable Phoenicopterus chilen sis con un alto número de individuos observados (1499, Tabla 1). A pesar de que estos ambientes permanecen mayor mente secos durante años (no se cuenta con registros documentados) y en perío dos particulares ingresa agua, pueden ser muy relevantes para las dinámicas poblaciones de las diversas especies que lo utilizan sobre todo en un contexto de ambientes áridos, donde el agua es un re curso altamente limitante.
Finalmente, los patrones de baja diver sidad, baja equitatividad, baja riqueza de gremios tróficos, patrón de diversidad-dominancia de tipo logarítmico, son consistentes con etapas sucesionales iniciales del sistema. Se resalta la nece sidad de evaluar la importancia de estos sistemas lagunares semipermanentes de ambientes áridos en las dinámicas pobla cionales del ensamble de aves. Esta infor mación cuyos muestreos se realizaron en el año 2008 puede considerarse una línea de base para futuros relevamientos com parativos y ser de interés para el diseño de acciones de manejo y conservación del humedal y su biodiversidad.

