











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCTION
Wetlands concentrate a large part of the continental biodiversity and therefore play a fundamental role in macroevolutionary pro cesses (Greb et al., 2006). Today, South American land ecosystems associated with the Amazon, Paraguay, and the southernmost Paraná-del Plata fluvial systems are part of the largest wet lands of the world (Orfeo & Neiff, 2008). The early history of these ecosystems dates back to the early Neogene, when they developed in tec tonically controlled environments linked to the compressive Andean orogeny, which remodeled the foreland basin landscape (Wesselingh et al., 2001; Hoorn et al., 2010, 2022; Tineo, 2020; Tineo et al., 2021). South American Neogene wetlands and associated fluvial systems are mainly known from several Miocene geological outcroppings such as the exceptional depositional and fossil records of Pebas-Solimões formations (close to the Peruvian-Colombian-Brazilian borders), the Honda Group (La Venta, Colombia), the Urumaco Formation (Urumaco, Venezuela), the Yecua Formation (Argentinian-Bolivian Sub-Andean zone) and the “Conglomerado osífero” (Ituzaingó Formation, Entre Ríos Province, Argentina) (e.g., Langston, 1965; Cione et al., 2000; Wesselingh et al., 2001; Hoorn et al., 2010, 2022; Riff et al., 2010; Salas-Gismondi et al., 2015, 2016, 2018; Tineo, 2020; Tineo et al., 2021). These sedimen tary beds had been deposited in a temporal range that goes approximately from the Early Miocene (ca. 23 Ma) to the Late Miocene (ca. 10.5-7/5 Ma) (e.g., Uba et al., 2009; Hoorn et al., 2010, 2022), and are mostly composed of shallow aquatic set tings of varying salinity and marine influence, such as lakes, swamps, and rivers, that repre sent different sequences of inland mega wet lands (e.g., Early to Middle Miocene sections of the Peruvian Pebas Formation and Late Miocene Yecua Formation; Nutall, 1990; Wesselingh et al., 2001; Boonstra et al., 2015; Hoorn et al., 2022; Uba et al., 2009; Tineo, 2020; Tineo et al., 2021), and predominantly fluvial or delta plain depos its (e.g., the Late Miocene Urumaco, Tranquitas, and Ituzaingó formations; Tineo et al., 2022; Quiroz & Jaramillo, 2010; Hoorn et al., 2010; Brandoni et al., 2019).
All these Miocene deposits achieve astonish ing associations of continental fossil vertebrates. Among them, Miocene crocodyliform assemblag es are characterized by the presence of a great di versity of amphibian neosuchians, the Crocodylia, and the much less diverse and abundant ter restrial sebecosuchians, such as Langstonia huilensis Langston (1965) (= Sebecus huilen sis; Paolillo & Linares, 2007) from the Middle Miocene of La Venta, Colombia. Crocodylia is the best represented group, with a large diversity of gavialoids (e.g; Siquisiquesuchus venezuelensisBrochu & Rincón, 2004; Aktiogavialis caribesiSalas-Gismondi et al., 2018; Dadagavialis gunai Salas-Gismondi et al., 2018) and several species of Gryposuchus (Gürich, 1912) and alligatoroids (e.g., Gnatusuchus pebasensis Salas-Gismondi et al., 2015; Kuttanacaiman iquitosensis Salas-Gismondi et al., 2015; Globidentosuchus brachy rostrisScheyer et al., 2013; Centenariosuchus gilmoreiHastings et al., 2013; Culebrasuchus mesoamericanus Hastings et al., 2013; Acresuchus pachytemporalisSouza-Filho et al., 2018; PaleosuchusGray, 1862; several species of CaimanSpix, 1825, PurussaurusBarbosa-Rodrigues, 1892, and MourasuchusPrice, 1964). These crocodylians show a high range of mor phological disparity and body sizes indicative of the ecological diversity that this group achieved during the Neogene (an event that in terms of morphological disparity and taxonomic diversity occurs for the only time during the entire evolu tionary history of these lineages). Salas-Gismondi et al. (2015, 2016) analyzed part of these hyper diverse crocodylian fossil records. These authors concluded that several Miocene assemblages are probably vinculated with different paleoenviron ments developed in relation to the geological evo lution of basins during the Neogene in Northern South America. They proposed that crocodylian associations in shallow water paleoenvironments, like lagoons, swamps, and marginal marine em bayments (e.g., several early Middle Miocene de posits) are characterized by a predominance of gavialoids with body morphotypes typical of shal low water piscivores forms (e.g., adults with or bits not protruded such as Gryposuchus pachaka mue Salas-Gismondi et al., 2016, Piscogavialis jugaliperforatus and Siquisiquesuchus venezuel ensis) and medium sized alligatorids, including caimanines with crushing dentitions with more grinding feeding habits (e.g., Gnatusuchus pe basensis, Kuttanacaiman iquitosensis, Caiman wannlangstoni Salas-Gismondi et al., 2015). Among these assemblages, large forms of al ligatorids are less frequent, although the enig matic but probably “gulp feeding” Mourasuchus atopus Langston, 1966 (Langston, 1965; Riff et al., 2010; Tineo et al., 2015; Cidade et al., 2017), and the predator Purussaurus neivensisMook (1941) are also registered in some Middle Miocene South American localities (such as in the late Middle Miocene of La Venta, Colombia; e.g., Langston, 1965). These interpretations sug gest that although most of early Middle Miocene caimanine alligatorids would have been endemic and became regionally extinct, others such as Mourasuchus and Purussaurus persisted in the Middle-Late Miocene in fluvial dominated pa leoenvironments (e.g., Mourasuchus arendsiBocquentin-Villanueva, 1984, from Venezuela, Brazil and Argentina; M. amazonensis Price, 1964, from Brazil; M. pattersoni Cidade et al., 2017, from Venezuela; Purussaurus brasilien sis Barbosa-Rodrigues, 1892, from Brazil and P. mirandaiAguilera et al., 2006, from Venezuela, and Mourasuchus sp. from Bolivia) and would have coexisted with large specialized piscivores (e.g., IkanogavialisSill, 1970; Hesperogavialis Bocquentin-Villanueva & Buffetaut, 1981; Gryposuchus colombianus Langston, 1965; G. croizati Riff & Aguilera, 2008; G. neogaeusBurmeister, 1885). Crushing caimans, such as Globidentosuchus brachyrostris Scheyer et al., 2013 and Caiman wannlangstoni Salas-Gismondi et al., 2015, were also found in rocks of ~7-9 Ma of the Urumaco Formation, indicating that niche was also occupied by caimanines in what is be lieved to be the last relict of the Pebas System en vironments (Hoorn et al., 2010; Salas-Gismondi et al., 2016).
The southernmost record of these neogene South American crocodylians is in the Late Miocene of northeastern Argentina (see below). In general terms, this crocodylian association is taxonomically similar to that of lower latitudes, with a greater diversity of Caimaninae, domi nated by medium to large sized caimans (e.g., Caiman australisBurmeister (1883); C. lute scensRovereto (1912), C. gasparinaeBona & Paulina Carabajal, 2013 and Caiman cf. C. lat irostris, Bona & Barrios, 2015; Bona et al., 2013) and a gavialoid, Gryposuchus neogaeus, that was interpreted as a highly aquatic and underwater feeding piscivorous crocodylian given its reduced skull pneumaticity (Bona et al., 2017). Among the large forms, Mourasuchus arendsi (= M. na tivusGasparini, 1985; Cidade et al., 2018) and generalist animal eating caiman like forms are also registered. Although several huge caimanine species have been described (e.g., Bravard, 1858; Burmeister, 1885; Rovereto, 1912; Rusconi, 1933; Gasparini, 1981, 1985), only C. lutescens and C. gasparinae (represented by a skull table and a partial skull, respectively) are currently recog nized and the presence of the Miocene mega predator Purussaurus was never mentioned in these latitudes (e.g., Bona & Barrios, 2015; Bona et al., 2013).
We present herein the first record of Purussaurus in the Late Miocene of Argentina together with a detailed description of this mate rial and a discussion of its paleogeographic and paleoenvironmental implications.
Geological and Paleontological Settings
In the Entre Ríos Province, the “Conglomerado osífero” crops out discontinuously in the river banks of the Paraná River and its tributar ies, from the city of Paraná northwards to Hernandarias (Fig. 1A). The tooth here described was collected in the Toma Vieja locality (31° 42’ 10’’S, 60° 28’ 35’’W), where this conglomerate is clearly distinguished by a visible erosive uncon formity from the mudstone and sandstone beds of the underlying Paraná Formation (Fig. 1B). The “Conglomerado osífero” is characterized by well consolidated levels of fine cross stratified gravel sandstone and well sorted fine to coarse grained cross stratified sandstone beds, and clayed mud stone lenses, which concentrated abundant fos sil bones and teeth of fragmented and disasso ciated vertebrates (Brandoni & Noriega, 2013). These skeletal fragments are heavy and well mineralized and belong mostly to continental vertebrates, although a few mostly reworked ma rine forms can be represented (e.g., Cione et al., 2000). Traditionally, the “Conglomerado osífero” was considered as part of the fluvial system of the Ituzaingó Formation that outcrops in Entre Ríos Province (Frenguelli, 1920; Aceñolaza, 1976, 2000; Brunetto et al., 2013; Schmidt et al., 2020). Particularly, Brunetto et al. (2013) and Brandoni et al. (2019) regarded the “Conglomerado osífe ro” as part of the Lower Member of the Ituzaingó Formation and interpreted that it corresponds to a continental paleoenvironment dominated by a braided fluvial succession. However, Pérez (2013a) considered that these levels correspond to fluvial tide influenced channels of a marginal marine paleoenvironment and assigned them to the upper levels of the Paraná Formation.
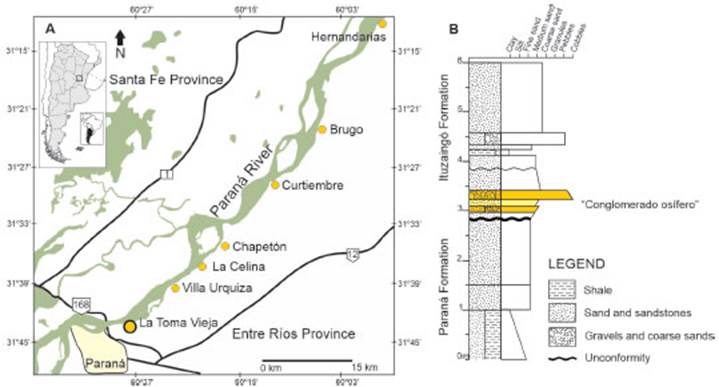
In the absence of radiometric dating, the age of the “Conglomerado osífero” has been estimated by biostratigraphic correlation to other Neogene units, based on its mammal content and stratigraphic position. Thus, the age of the “Conglomerado osífero” is mainly interpreted to be Huayquerian (ca. 9-6.8 Ma sensuCione et al., 2000), or alternatively to the Chasicoan-Huayquerian lapse (ca. 10-6.8/5.3 Ma, Brandoni, 2013), and consequently corre lated to the Tortonian-Messinian (Late Miocene) (International Chronostratigraphic Chart, 2022). 86Sr/87Sr dating of mollusks from the underly ing Paraná Formation in the locality of Punta Gorda (Entre Ríos Province; Fig. 1A), indicates ages of 9.47 Ma (Tortonian; Pérez, 2013b) and 7.55-6.67 Ma (Tortonian-Messinian; del Río et al., 2018). Considering the relationship between the association of vertebrates recovered from the “Conglomerado osífero” and that of other Neogene units (see Cione et al., 2000; Brandoni, 2013; Schmidt et al., 2020), as well as the proposed age for the Paraná Formation (Pérez, 2013b; del Río et al., 2018), a Late Miocene age can be sug gested for the “Conglomerado osífero”.
MATERIAL AND METHODS
MAS-PV 386 was described macro and micro scopically. For the macroscopic anatomical analy sis comparisons with teeth of extant and Miocene caimanines were made, with first hand materi als, photographs taken from literature (Tab. 1), or own photographs. Terminology used for the spatial orientation of the tooth follows Smith & Dodson (2003); however, we decided to add the term “vestibular” as synonym of “labial”, since reptiles do not have lips. The microscopic analy sis was made by the Environmental Scanning Electron Microscope ESEM-FEI Quanta 200 of the LIMF. In addition, chemical element map ping tools (major elements) were used, and Backscattered Electrons (BE) method. The lat ter, based on the detection of the atomic num ber (Z), provides an image of the distribution of chemical elements in the material that makes up the entire piece, with the darkest areas be ing those with the lowest Z and the lightest and bright ones with higher Z (Fig. 2).
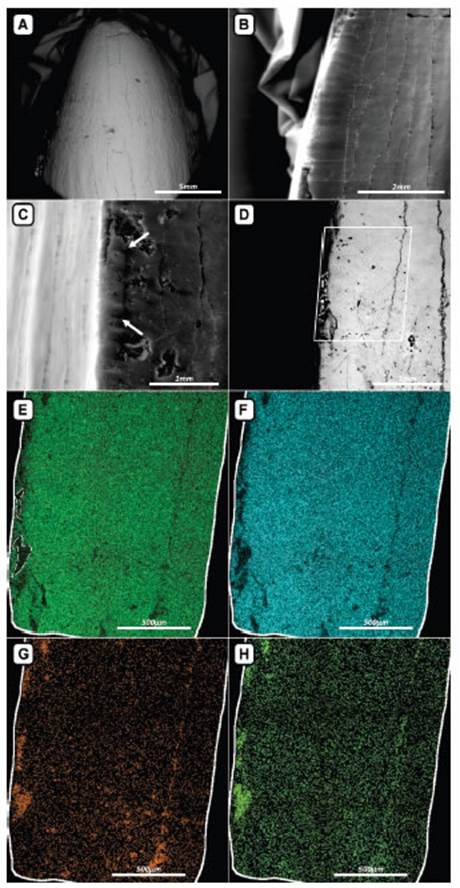
Fig. 2 Images of the tooth taken from the Environmental Scanning Electron Microscope (ESEM). (A) general view of the labial surface without metal coverage, using the low vacuum technique showing the vertical fractures and the cavities in the surface of the piece. (B) detail of the crenulated margin on the lingual aspect of the tooth, with vertical and transverse fractures. (C) detail of the mesio-distal edge of the tooth in vestibular (labial) view showing the crenulation typical of pseudozifodont teeth (indicated by arrows). (D) vestibular aspect of the tooth observed with Backscattered electrons showing fractures (in black) and positive surfaces (in white); the box delimits the area of mapping of chemical elements, carried out with EDAX. (E-H), details of the distribution of the chemical elements after the EDAX analysis: (E) phosphorus; (F) calcium; (G) iron; (H) silicon distributions. Note that the iron and the silicon are present mainly in the fractures and cavities of the tooth.
Institutional Abbreviations. AMU-CURS, Alcaldía del Municipio de Urumaco, Urumaco, Venezuela; DGM, Divisão Geologia y Mineralogia, Rio de Janeiro, Brazil; MACN-PV, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Colección Paleontología Vertebrados, Ciudad Autónoma de Buenos Aires, Argentina; MAS-PV, Museo de Ciencias Naturales y Antropológicas “Prof. Antonio Serrano”, Colección Paleontología Vertebrados, Paraná, Entre Ríos, Argentina; MCC-URU, Museo de Ciencias Naturales de Caracas-Urumaco, Caracas, Venezuela; MLP, Museo de La Plata, La Plata, Buenos Aires, Argentina; LIMF, Laboratorio de Investigaciones de Metalurgia Física “Ing. Gregorio Cusminsky”, Facultad de Ingeniería, Universidad Nacional de La Plata, Buenos Aires, Argentina, UFAC, Universidade Federal do Acre, Rio Branco, Brazil.
RESULTS
Systematic Paleontology
CrocodyliformesHay, 1930 (Benton & Clark, 1988)
CrocodyliaGmelin, 1789
ALLIGATORIDAE Gray, 1844
CaimaninaeBrochu, 1999
PurussaurusBarbosa-Rodrigues, 1892
Purussaurus sp.
Type species. Purussaurus brasiliensisBarbosa-Rodrigues, 1892.
Referred material. MAS-PV 386, incomplete tooth preserving the apex sector of an almost well preserved crown (Fig. 3).

Fig. 3 Isolated tooth of Purussaurus sp. MAS-PV 386 in (A) vestibular; (B) lingual; and (C) mesio-distal views. Abbreviations: cr, crenulations. Scale bar = 20 mm.
Locality and horizon of the new specimen. Toma Vieja locality, near Paraná City (Entre Ríos Province, Argentina), “Conglomerado osífero” (traditionally considered as the base of Ituzaingó Formation, Brunetto et al., 2013), Late Miocene (Brandoni, 2013; Brunetto et al., 2013; Schmidt et al., 2020).
Description. MAS-PV 386 consists of a frag mentary tooth that preserves most of the apical sector of the crown, which is conical but slightly compressed and lingually curved. In lingual and vestibular views, the mesial and distal margins are straight in the apical region but more curved in the base of the preserved part of the crown, where the tooth is subcircular in transverse sec tion, with a vestibular-lingual width equal to 16.6 mm, and a mesiodistal width equal to 19.7 mm (both measured at the base of the piece). This minimal but abrupt increase in tooth width due to curving of the mesial and distal margins is present, although more marked, in other teeth assigned to Purussaurus (e.g., DGM 1128, Souza et al., 2016, figure 3 F-H).
The tooth apex is rounded, showing a partial wear of the enamel apical surface, a condition commonly seen in worn or shed teeth of all croc odiles. The enamel of the lingual and vestibular (or labial, sensuSmith & Dodson, 2003) surfaces of the crown are uniformly ornamented with fine anastomosed striae or ridges separated by smooth grooves, extending from the apex to the base along the entire surface of the crown. These fine apicobasal ridges are transversely crossed by thin irregular lines, which observed under mag nification give the enamel a cracked appearance.
The crown shows a conspicuous carina that extends continuously along the mesial and dis tal faces of the tooth dividing the crown into a vestibular and a lingual subequal sectors. This crenulated carina is preserved in several sectors of the mesial and distal margins, it is exclusively formed by the enamel and bears fine but very marked parallel crenulations, oriented perpen dicularly to the carina. Under magnification (SEM) it can be verified that these crenulations are formed by rounded parallel short crests sepa rated by a shallow valley of enamel, distributed on both the labial and lingual surfaces of the carina (Figs. 2B-C, 3C). The density of these car inae is 5 per millimeter. This condition is called pseudoziphodonty (or false ziphodonty; Prasad & de Broin, 2002) and differs from the ziphodont condition (serrated teeth) present in other archo saurs, in which the crenulations are formed by both the dentine and the enamel (Prasad & de Broin, 2002).
Several features acquired during the fossil ization and depositional processes of MAS-PV 386 can be observed. In a macroscopic view, dam aged areas are present at the base and the mesial and distal margins of the tooth, with worn sur faces and fractures (Fig. 3). Under magnification (SEM) an evident wear can be observed at the apex of MAS-PV 386, and several excoriations associated with longitudinal microfractures run ning from the apex to the base and connected to each other with finer subhorizontal microfrac tures (Fig. 2A). The longitudinal micro fractures are arranged in the smooth grooves or valleys between the apicobasal striae that constitute the general ornamentation of the crown (Figs. 2A, 3). The vestibular surface of the tooth shows a process of micro alveolarization, identified from the presence of small depressions distributed all over the surface of the enamel. Finally, the tooth is broken at the base and, given the nature of this fracture, it can be inferred that it was sepa rated from the skull after fossilizing.
Comparisons. Teeth preserved implanted in the upper and lower jaws are scarce in the croco dylian fossil record. In the case of Purussaurus, few teeth were recovered in situ and described in the literature (e.g., Langston, 1965; Aguilera et al., 2006; Aureliano et al., 2015; Souza et al., 2021). A gradual transition in the shape and size of teeth has also been described for this taxon, from taller and pointed anterior teeth to broader, lower, and more bulbous posterior ones (Langston, 1965; Aureliano et al., 2015). The teeth of Purussaurus have been described as rounded in a basal section but with crowns tending to be compressed (Langston, 1965) and bearing pseudoziphodont ridges. Anteriormost, larger, conical and sharp teeth of both upper and mandibular series of Purussaurus have been de scribed as slightly compressed and lingually and distally curved (Langston, 1965; Aguilera et al., 2006; Aureliano et al., 2015). In P. mirandai the premaxillary teeth are sharp, anteroposteriorly and labiolingually curved; these teeth are rela tively compressed as MAS-PV 386 (with a ves tibular-lingual width of 15-30 mm and a mesi odistal width of 19-39 mm, sensu Aguilera et al., 2006). Due to its dimensions and proportions, MAS-PV 386 is more similar to the premaxillary second tooth of P. mirandai (e.g., AMU-CURS-135, described by Aguilera et al., 2006), although given that MAS-PV 386 is an isolated tooth, its relative size and position in the tooth row cannot be specified since the body size of the individual is unknown. Some posterior maxillary teeth of P. mirandai are preserved in some specimens (e.g., AMU-CURS-135, MCC URU-115-72V) and as in most of caimanines, these teeth are blunt, more rounded and lower than the anterior ones, with a general morphology clearly different from MAS-PV 386. The dentary teeth arrangement of P. mirandai is similar to other caimanines with the anterior teeth larger, taller and sharper, being the mandibular tooth 4 the tallest of the first teeth preserved (e.g., AMU-CURS-135). In this species mandibular teeth are more compressed than the teeth of the upper jaw and than in MAS-PV 386. Following this reasoning, MAS-PV 386 could cor respond to a tooth of the upper series.
In relation to the crown ornamentation, the teeth of Purussaurus have been described as pseudoziphodont (see below). All these teeth pres ent mesiodistal carinae, sometimes incompletely preserved and with fine striations perpendicular to the surface of the crown. In P. mirandai, the third premaxillary teeth of MCC URU-115-72V and some isolated teeth associated with AMU-CURS-135 show a striae density of 3 per 5 mm, different from MAS-PV 386 in which the density of these striae is 5 per millimeter. Finally, the enamel of isolated teeth assigned to Purussaurus has been described by several authors (e.g., Langston, 1965; Aguilera et al., 2006; Aureliano et al., 2015; Souza et al., 2016, 2021) which con clude that in smaller teeth (especially in the posterior and more bulbous ones), the enamel is generally wrinkled whereas larger teeth gener ally show smooth enamel but with longitudinal and transverse lines along the crown, conditions similar to those of MAS-PV 386.
DISCUSSION
Among crocodyliforms, ziphodont teeth are characteristic of carnivorous species with in ferred terrestrial habits (as well as a few marine metriorhynchids; Gasparini et al., 2006). Ziphodont teeth are common among predatory species within the diverse clade Notosuchia, such as peirosaurids (e.g., MontealtosuchusCarvalho et al., 2007, Lomasuchus Gasparini et al., 1991) and baurusuchids (e.g., BaurusuchusPrice, 1945; e.g., Riff & Kellner, 2011) from the Cretaceous and the sebecids from the Paleogene-Neogene of South America (e.g., SebecusSimpson, 1937; Sahitysuchus Kellner et al., 2014; e.g., Colbert, 1946). Ziphodont teeth also occur in eusuchians of supposedly terrestrial habits such as the planocraniids (e.g., PlanocraniaLi, 1976 and BoverisuchusKuhn, 1938; e.g., Brochu, 2012) and mekosuchines (e.g., QuinkanaMolnar, 1981 and KambaraWillis et al., 1993; e.g., Brochu, 2001; Buchanan, 2009). Pseudoziphodont teeth are less common in non eusuchian crocodyli forms, being reported in some peirosaurids (e.g., BarcinosuchusLeardi & Pol, 2009) and atopos aurids (Venczel & Codrea, 2019). The presence of pseudoziphodont teeth is even less frequent within Crocodylia. In particular, the only record of pseudoziphodont teeth for the Cenozoic of South America is restricted to the large caima nine Purussaurus and, in fact, this feature has been regarded as a diagnostic character for this genus (Souza et al., 2021).
The general morphology of MAS-PV 386 is like the anterior teeth of specimens assigned to Purussaurus from northern regions (e.g., Brazil, Colombia), in which the anterior teeth have slightly flattened crowns subcircular at their bases (Langston, 1965; Aguilera et al., 2006; Aureliano et al., 2015; Souza et al., 2016, 2021). Considering the general structure of the enamel of MAS-PV 386, it also resembles specimens as signed to Purussaurus in having longitudinal and transverse striae along the crown (e.g., DGM 1128-R and DGM 1194-R, Souza et al., 2016, fig ure 3 F-I; Aureliano et al., 2015, figure 5). The pseudoziphodonty, the macro and microscopic ap pearance of the enamel, together with the general shape of the tooth, allows us to assign MAS-PV 386 to the genus Purussaurus. However, the lack of species level diagnostic features in the teeth of each species of this genus precludes assigning MAS-PV 386 to any of the known Purussaurus species. Furthermore, MAS-PV 386 differs from teeth assigned to P. mirandai, P. neivensis, and P. brasiliensis, which have been described as curving backwards and slightly inwards in these species (Langston, 1965; Aguilera et al., 2006; Aureliano et al., 2015; Souza et al., 2021). As oc cur in most generalist alligatorids, the dentition of Purussaurus is heterodont in shape and size, showing a general transition from taller conical pointed anterior teeth (with some hypertrophied teeth such as the 4th dentary tooth, 3rd/4th pre maxillary teeth and 2nd/3rd maxillary teeth) to lower and rounded posterior teeth which are more button like shaped (e.g., Aguilera et al., 2006; Aureliano et al., 2015). After the anatomi cal comparisons here made, MAS-PV 386 is with in the range of size variation of Purussaurus and the morphology of the crown indicates that it is probably an anterior tooth. As it is an isolated tooth, the position of MAS-PV 386 in the teeth row cannot be specified. However, compared with the proportions of the less compressed upper teeth and the relatively more compressed lower teeth of Purussaurus mirandai, the proportions of MAS-PV 386 indicate that it could be a tooth from the upper series (premaxillary tooth or an terior maxillary tooth).
In addition to the anatomical features, the preservation traits observed in MAS-PV 386 al low us to recognize the effects of the taphonomic processes that affected the material. From the BE analysis of the SEM, differences in the chemical elements and density of the materials that com pose the surface of the tooth are identified (Fig. 2 EH). By mapping the chemical elements that conform the surface of the tooth, it was possible to verify the typical presence of calcium (Ca2+) and phosphorus (P5+), which constitute the original components of the tooth (hydroxyapa tite), but also the presence of silicon (Si) and iron (Fe). In MAS-PV 386, Si and Fe fill the fractures and probably come from the silicoclastic sedi ment of the conglomerate (terrigenous material) and from authigenic precipitation as ferruginous cement (hematite, Fe2O3), respectively, during the fossil-diagenetic processes (Figs. 2G-H). The presence of longitudinal, transverse, and perpen dicular fractures (respect of the apicobasal axis of the tooth) (Fig. 2AD) is characteristic of a fragile tooth that loses the organic component that gives it the resistance of the structure. These fractures together with the small depressions distributed all over the vestibular surface of the enamel (mi cro alveolarization) would indicate that the piece suffered some abrasion by roll of little magnitude generated by the silicoclastic granular sediment of the bottom of the fluvial channel where it was accumulated.
Analyzing crocodylian assemblages from other contemporary South American localities, such as Solimões and Urumaco formations, Purussaurus is one of the taxa that cohabited in the Late Miocene mega wetland systems. Within these associations, caimanines (such as Mourasuchus and different Caiman species) and gavialoids (such as Gryposuchus) are always registered showing differences in their sizes and snout, jaws and teeth shapes. This morphologi cal variability has been interpreted as evidence of niche partitioning, which results in optimizing the availability of resources and avoiding compe tition (Aureliano et al., 2015). In addition to the large size of Purussaurus species, the pseudozi phodont teeth suggest that they would have been active predators that included large vertebrates in their diets (e.g., Pujos & Salas-Gismondi, 2020). This study confirms that Purussaurus would have been a component of the Late Miocene Argentinian crocodylofauna occupying a separate niche from piscivorous gavialoids, animalivore caimans and Mourasuchus (Tab. 2).

Table 2 List of crocodylians of the “Conglomerado osífero” (Ituzaingó Formation, Paraná Province, Argentina).
The sedimentological characteristics ob served in the “Conglomerado osífero” in the Toma Vieja locality are mainly given by channel fill deposits interpreted as a braided fluvial sys tem (e.g., Brunetto et al., 2013; Brandoni et al., 2019). As mentioned above, these levels rest un conformably on the shallow marginal marine de posits of the Paraná Formation through an ero sive boundary, interpreted by some authors as a regional erosive surface related to the basal sec tion of the Ituzaingó Formation (Brunetto et al., 2013; Brandoni et al., 2019). The erosive coarse-grained deposits of the “Conglomerado osífero”, together with the taphonomic characteristics of most of its fossils (which are disarticulated, frag mentary, and with evidence of transport) suggest that this association represents a mixture of fau nas, mainly continental with few marine taxa of different ages (Cione et al., 2000; Schmidt et al., 2020). Beyond this scenario, the now documented presence of Purussaurus in the “Conglomerado osífero” suggests that ecosystems associated with the southernmost South American Miocene wetlands would have been taxonomically simi lar to those of lower latitudes (e.g., Hoorn et al., 2010, 2022; Tineo et al., 2015; Tineo, 2020). Particularly the presence of Gryposuchus neo gaeus and caimanines such as Mourasuchus ar endsi, Purussaurus, and several species for the moment assigned to Caiman (Bona & Barrios, 2015; Bona et al., 2013) in the Late Miocene of Paraná (Tab. 2) proposes a wide geographic dis tribution of these Neogene fluvial systems with a similar ecological complexity for the entire re gion in South America. Among the fossil materi al of crocodylians of the “Conglomerado osífero”, Gryposuchus neogaeus is the most complete specimen recovered so far (i.e., MLP 26-413, an almost complete skull; Gasparini, 1968). Caimanines, on the other hand, are represented by isolated skulls and postcranial fragments which would indicate that most of these specimens will have been transported from their source area. In this scenario, MAS-PV 386 is another example of this taphonomical features. Although a regional envi ronmental reconstruction is necessary to inter pret the different continental sub-environments that would have developed in this part of the basin, the taxonomic diversity of crocodylians recognized in the “Conglomerado osífero” indi cates the presence of warm humid environments with large amounts of water, capable of to with stand large piscivores gavialoids and huge cai manines along the megapredator Purussaurus.
CONCLUSIONS
The record of the tooth MAS-PV 386 in the “Conglomerado osífero” in Entre Ríos Province allows us to propose for the first time the pres ence of Purussaurus in the Late Miocene of Argentina. Although isolated indeterminate re mains of large crocodylians have been described in the “Conglomerado osífero” (such as cranial and postcranial fragments of vertebrae, ribs, long bones, and osteoderms; Bravard, 1858; Burmeister 1883, 1885; Rovereto, 1912; Rusconi, 1933), at the moment this genus was geo graphically restricted to Miocene beds of Brazil, Colombia, Perú, and Venezuela (Aguilera et al., 2006; Aureliano et al., 2015).
This new finding represents the southern most record of one of the largest known preda tors of neosuchian crocodiles. The presence of this mega carnivore alligatorid suggests a greater taxonomic diversity for the huge caimanines in Late Miocene wetlands in northeast Argentina. The potential faunistic association of large croco dylians such as Gryposuchus, Purussaurus, and giant caimans allows to explore the hypothesis of a niche partitioning already inferred for other Miocene northernmost South American localities given by the coexistence of piscivorous and other animal eating crocodylians. This possible setting proposes the existence of more complex environ ments capable of sustaining all this crocodilian fauna and therefore a more complex scenario for inland wetland and fluvial ecosystems devel oped in the Late Miocene in the South American Chaco Parana Basin.

