











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
Debido a la magnitud que han alcanzado los cambios producidos por el ser humano en la Tierra, los ecosistemas que se desarrollan espontáneamente, como respuesta natural a las nuevas condiciones ambientales creadas por la actividad humana son, desde una perspectiva ecológica, cada vez más relevantes (Lugo, 2009). Dichos ecosistemas, conocidos como neoecosistemas (“novel ecosystems”), resultan de la coocurrencia de especies en combinaciones y abundancias relativas que no se han verificado previamente para un dado bioma (Hobbs et al., 2006). En muchos casos, se trata de áreas en las que las especies exóticas invasoras son las dominantes o más frecuentes, mientras que las especies de plantas nativas se encuentran como acompañantes (Morello et al., 2000). Actualmente los neoecosistemas ocupan alrededor del 40% de la tierra libre de hielo, por lo que se han vuelto críticos para la conservación de la biodiversidad y la provisión de servicios ecosistémicos (Bridgewater et al., 2011). De este modo, es necesario entender estos sistemas para poder predecir su estado futuro y su correcto manejo adaptativo (Seastedt et al., 2008).
Las áreas naturales protegidas (ANP) son esenciales para la conservación de la diversidad biológica y el mantenimiento de un ambiente saludable para las personas (Naughton-Treves et al., 2005; UNEP-WCMC & IUCN, 2016), siendo la biodiversidad local mayor dentro de las ANP que fuera de ellas (Gray et al., 2016), inclusive sin haberse desarrollado todo su potencial de conservación (Gaston et al., 2008; Watson et al., 2014). Muchas veces se han cuestionado las posturas de conservación que consideran a los ecosistemas urbanos, a menudo asociados a un gran número de especies no nativas. Sin embargo, es prometedor considerar sus servicios ecosistémicos asociados, sus beneficios sociales y su contribución a la conservación de la diversidad biológica (Dearborn & Kark, 2010; Kowarik, 2011). El establecimiento de las especies exóticas en ecosistemas urbanos está asociado a su cercanía a centros urbanos, tamaño y densidad poblacional humanos (Kuhman et al., 2010; Gaertner et al., 2017). En particular, las áreas naturales urbanas brindan la oportunidad de llevar a cabo tareas de conservación y de educación ambiental sobre los procesos ecosistémicos que allí tienen lugar. Además, promueven una mayor participación e involucramiento de los ciudadanos a través de la ciencia ciudadana, la restauración y el monitoreo ambiental (Dearborn & Kark, 2010).
Las ANP de la costa bonaerense del Río de la Plata se encuentran declaradas tanto sobre ambientes remanentes como sobre sustratos jóvenes (Cabrera & Dawson, 1944; Barbetti et al., 1985; Faggi & Cagnoni, 1987; Kalesnik et al., 2005) y se encuentran influenciadas por tres regiones fitogeográficas distintivas: Delta del Paraná, Espinal y Pampeana (Burkart et al., 1999; Oyarzabal et al., 2018). Según Passarelli et al. (2014), en los humedales próximos al Río de la Plata pueden observarse comunidades herbáceas y leñosas de especies nativas, que incluyen también la presencia de algunas exóticas. Las ANP rioplatenses se encuentran, por un lado, insertas o muy próximas a centros urbanos muy densamente poblados y, por otro lado, próximos al Río de la Plata, cuyas márgenes constituyen un importante biocorredor que facilita el intercambio de fauna y propágulos vegetales (Capllonch et al., 2008; Guerrero et al., 2017).
La línea de costa de la ciudad de Buenos Aires ha sufrido modificaciones ambientales desde su primera fundación hace más de 500 años (Athor, 2012; Brailovsky, 2018). Su perfil es producto de sucesivos rellenos realizados desde el año 1836 sobre la planicie estuárica del Río de la Plata (Nabel & Becerra Serial, 2008). Esta técnica de expansión de la costa por relleno es en la actualidad una tendencia mundial, mayormente mediada por factores antropogénicos, que tiende a la reducción del área ocupada por humedales costeros (Sengupta et al., 2018) y la pérdida de biodiversidad (Yang et al., 2011; Duan et al., 2016). En el caso particular de la costa rioplatense, en cada progradación la hidrodinámica natural del río fue generando nuevos ecosistemas adaptados a la configuración costera creada por los distintos sistemas de relleno (Marcomini & López, 2004).
Uno de los últimos rellenos antrópicos sobre el Río de la Plata, realizado durante las décadas de 1960 y 1970, corresponde al terreno donde hoy se asienta la Reserva Ecológica Ciudad Universitaria - Costanera Norte (RECU-CN). Esta ANP fue creada mediante la Ley Nº 4467 en el año 2012, pero no fue hasta marzo del año 2018, cuando se firmó el convenio entre los organismos que deben cogestionar el área (la Universidad de Buenos Aires y el Gobierno de la Ciudad Autónoma de Buenos Aires), que entró en vigencia dicha Ley. Para cumplir con los objetivos particulares de cualquier ANP es necesario contar con una línea de base de los recursos naturales del lugar, permitiendo así caracterizar los aspectos naturales y planificar el manejo de los mismos (APN, 2010). El área ha sido recorrida y utilizada con fines académicos desde sus orígenes, principalmente por docentes, investigadores y estudiantes de la Universidad de Buenos Aires. En cuanto a la vegetación del área, hasta el momento se realizó un listado preliminar de la flora vascular de un sector de 5 ha (Santo Domingo Sartori & Picca, 2009) y una recopilación de la riqueza acumulada históricamente (Sirolli et al., 2018), pero no existen trabajos publicados generados en forma sistemática que abarquen toda el área. Por lo tanto, el objetivo del presente trabajo fue describir la vegetación de plantas vasculares de la RECU-CN mediante la identificación de gradientes ambientales en los que se ordena la vegetación y la clasificación de unidades homogéneas. Se planteó como hipótesis que las especies de plantas vasculares presentes en la RECU-CN se encuentran en combinaciones distintas a las de las comunidades de plantas naturales descriptas en la bibliografía de la región, conformándose como neoecosistemas.
MATERIALES Y MÉTODOS
Área de estudio
El área de estudio se encuentra al norte de la Ciudad Autónoma de Buenos Aires, dentro de la Comuna 13, a orillas del Río de la Plata, y abarca una superficie total de 23 ha (Fig. 1). Sus límites son el muro de contención de la Ciudad Universitaria, la desembocadura del arroyo White, el Río de la Plata y el Parque de la Memoria. En 1936 el predio ocupado actualmente por Ciudad Universitaria y la RECU-CN no existía, llegando el río hasta las vías del ferrocarril General Belgrano (Prezzi et al., 2011). Este terreno fue ocupado por rellenos sucesivos que ocuparon la plataforma de abrasión que afloraba en toda la costa de la ciudad, correspondiente al paleoacantilado que conformaban las barrancas de Belgrano y Nuñez (Prezzi et al., 2011). En particular, el área de estudio corresponde a un proceso parcial de polderización, realizado mediante la construcción de un terraplén unido a la costa sólo en un extremo, que delimitó una península y un humedal costero estuárico conectado al río por un canal de mareas ubicado al norte del área (Marcomini & López, 2004). El clima es subtropical húmedo con una temperatura media anual de 17,9 °C y una precipitación anual de 1082 mm (estación Aeroparque Jorge Newbery, Servicio Meteorológico Nacional, 2020). El área es afectada por el nivel del río, dominado mayormente por los vientos, dándose fenómenos climatológicos como las sudestadas. Este fenómeno se presenta como vientos provenientes del sector sudeste con persistencia regular e intensidades de moderadas a fuertes, que producen inundaciones en la costa del Río de la Plata con duraciones de uno a tres días (Kreimer et al., 2001).
Obtención de datos
El tamaño de la parcela se definió en 5 x 5 m (25 m2) luego de realizar un muestreo piloto en ocho puntos seleccionados al azar procurando tener en cuenta las distintas fisonomías vegetales detectadas “a priori” (comunidades herbáceas, arbóreas, terrestres y acuáticas). En cada punto se contabilizó el número de especies presentes en tamaños de subparcelas crecientes desde 0,3 x 0,3 m hasta 10 x 10 m en saltos que duplicaban la superficie de la parcela anterior, y se estableció el tamaño óptimo por la determinación del punto de inflexión de la curva de acumulación de especies (Mueller-Dombois & Ellenberg, 1974). Una vez determinado el tamaño de parcela óptimo se procedió con el muestreo, disponiendo las parcelas de manera sistemática en toda el área de estudio en las intersecciones de una grilla de 50 x 50 m (Fig. 1). En total se estudiaron 79 parcelas, correspondiente a una superficie de 0,1975 ha, de las cuales dos no fueron consideradas por estar cubiertas sólo por hormigón. El muestreo se realizó de junio a agosto de 2018 durante un total de 14 jornadas, incluidas aquellas correspondientes al muestreo piloto.
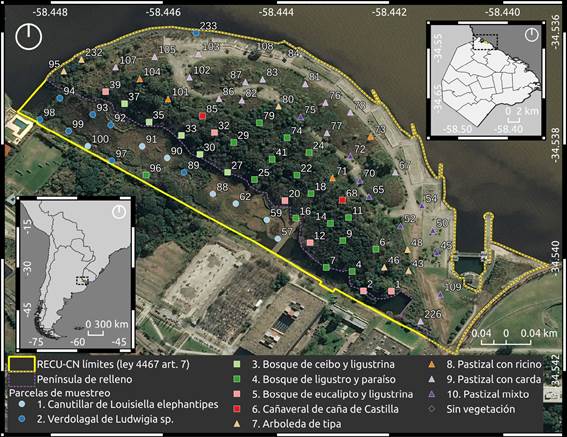
Fig. 1 Ubicación de la Reserva Ecológica Ciudad Universitaria - Costanera Norte (RECU-CN). Se indican los límites de la RECU-CN (línea amarilla), la ubicación de las parcelas de muestreo (números, no consecutivos), las unidades de vegetación Humedal (círculos), Bosque (cuadrados) y Pastizal con arboledas (triángulos) y las 10 comunidades de plantas (distintos colores, caña de castilla = Arundo donax, carda = Dipsacus fullonum, ceibo = Erythrina crista-galli, eucalipto = Eucalyptus sp., ligustrina = Ligustrum sinense, ligustro = L. lucidum, ricino = Ricinus communis, tipa = Tipuana tipu). Figura en color en la versión en línea http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/908/1194
La cobertura de cada especie se estimó visualmente mediante el método de Braun-Blanquet considerando las siguientes nueve categorías de rangos porcentuales 0-0,1; 0,1-1; 1-5; 5-10; 10-25; 25-33; 33-50; 50-75 y 75-100 (van der Maarel & Franklin, 2013). Aquellos ejemplares cuya identificación no pudo ser resuelta a campo fueron herborizados para su posterior determinación en el laboratorio. Para la determinación de los materiales se consultaron las floras regionales de Buenos Aires (Cabrera, 1963, 1965a, 1965b, 1967, 1968, 1970) y de Entre Ríos (Burkart, 1969, 1974, 1979, 1987; Burkart & Bacigalupo 2005). De los 154 taxones registrados, 124 fueron identificados a nivel de especie, 20 a nivel de género y 10 a nivel de familia u otra jerarquía. Algunas especies no pudieron ser determinadas y son tratadas como morfotipos. La nomenclatura de las especies empleada en este trabajo siguió el criterio de Zuloaga et al. (2019), disponible en línea en el sitio web del Instituto de Botánica Darwinion (http://conosur.floraargentina.edu.ar/ [consultado marzo-noviembre 2019]). En la Tabla 1 se detalla la lista completa de especies con sus autores.
Análisis de datos
A escala global, de unidad de vegetación y de comunidad se calculó la riqueza como el número acumulado de taxones. Para cada taxón, la cobertura media correspondió al promedio de las coberturas registradas en las parcelas correspondientes, y la frecuencia relativa al total de parcelas en las que el taxón estuvo presente respecto al total de parcelas consideradas. Ambos valores se calcularon tanto para las unidades de vegetación como para las comunidades. Para la cobertura se utilizó la marca de clase de los rangos porcentuales utilizados. La riqueza total del área se estimó con la función iNEXT del paquete iNEXT (Hsieh et al., 2016) en R (R Core Team, 2019) a partir de la matriz de incidencia de los datos recolectados a campo (77 parcelas y 154 taxones). La riqueza, cobertura y frecuencia también fueron analizadas agrupando los taxones según su origen cuando pudo ser determinado (nativo o exótico) y su forma de vida (árbol, arbusto, subarbusto, hierba y trepadora).
Se realizaron análisis multivariados de la vegetación de la RECU-CN mediante R (R Core Team, 2019), previa transformación de los datos de cobertura a escala ordinal (1 a 9 para las nueve categorías porcentuales antes mencionadas, cero para ausencia). Dicha transformación se realizó con el objetivo de disminuir el riesgo de sobreenfatizar los taxones dominantes (Jensen 1978) y de corregir en forma aceptable los valores de cobertura para su análisis numérico (van der Maarel, 2007). La similitud en la composición de taxones de las parcelas y los gradientes ambientales en los que se ordenaban las parcelas fueron analizados mediante un Análisis de Correspondencia Destendenciado (DCA) realizado con la función decorana del paquete vegan (Oksanen et al., 2019). Con el mismo paquete y la función hclust se realizó la clasificación de las parcelas en función de su vegetación mediante una clasificación jerárquica aglomerativa, utilizando la distancia de Bray-Curtis y el ligamiento de Ward. Se aplicaron dos niveles de corte considerando los 30 procedimientos provistos en el paquete NbClust (Charrad et al., 2014). Las unidades de vegetación correspondieron a los grupos formados por el nivel de corte más frecuente de todos los considerados. Las comunidades de plantas fueron establecidas mediante el nivel de corte sugerido por los métodos Duda y Pseudot2 (Duda & Hart, 1973) debido a que han demostrado mejores desempeños en comparación con otros índices según Milligan & Cooper (1985) y Chouikhi et al. (2015). Debido a que algunas de las parcelas presentaron el césped cortado, sólo aquellas gramíneas que por sus características vegetativas y ecológicas resultaron inconfundibles (Arundo donax, Cortaderia selloana, Echinochloa polystachya y Louisiella elephantipes) se trataron independientemente en los análisis multivariados, mientras que las restantes fueron tratadas en conjunto dentro de la categoría “Poaceae spp.”, dando como resultado una matriz de 140 taxones por 77 parcelas.
RESULTADOS
Riqueza de plantas vasculares de la RECU-CN En el relevamiento se registraron 154 taxones correspondientes a un total de 144 especies, 123 géneros y 58 familias de plantas vasculares. Las especies Ligustrum sinense, Ligustrum lucidum y Melia azedarach resultaron ser las más abundantes y frecuentes, y las familias Poaceae y Asteraceae, las que tuvieron más representantes. Se registraron 79 especies nativas de la región (58% del total de las especies determinadas), que representaron cerca del 30% de la cobertura total. La riqueza total estimada de la RECU-CN fue de 238 especies con un intervalo de confianza del 95% de entre 195 y 327 especies. En cuanto a las formas de vida se registraron árboles, arbustos, subarbustos, hierbas y trepadoras, siendo las de mayor cobertura los árboles (46%) y las hierbas (44%) mientras que los arbustos, subarbustos y trepadoras se encontraron en baja proporción (3%, 3% y 4% respectivamente).
Tabla 1. Listado de especies registradas en la Reserva Ecológica Ciudad Universitaria - Costanera Norte. Se indica si la especie es nativa en la región, su forma de vida, frecuencia y cobertura en cada comunidad por unidad de vegetación. Abreviaturas: H, Humedal; B, Bosque; P, Pastizal con arboledas; comunidades: 1, Canutillar de Louisiella elephantipes; 2, Verdolagal de Ludwigia sp.; 3, Bosque de ceibo y ligustrina; 4, Bosque de ligustro y paraíso; 5, Bosque de eucalipto y ligustrina; 6, Cañaveral de caña de Castilla; 7, Arboleda de tipa; 8, Pastizal con ricino; 9, Pastizal con carda; 10, Pastizal mixto; E, exótica; N, nativa; ND, no determinada; FV, forma de vida; A, árbol; B, arbusto; H, hierba; S, subarbusto; T, trepadora; frecuencias: a, 1-20%; b, 21-40%; c, 41-60%; d, 61-80%; e, 81-100%; coberturas: 1, 0-0,1%; 2, 0,1-1%; 3, 1-5%; 4, 5-10%; 5, 10-25%; 6, 25-33%; 7, 33-50%; 8, 50-75% y 9, 75-100%.
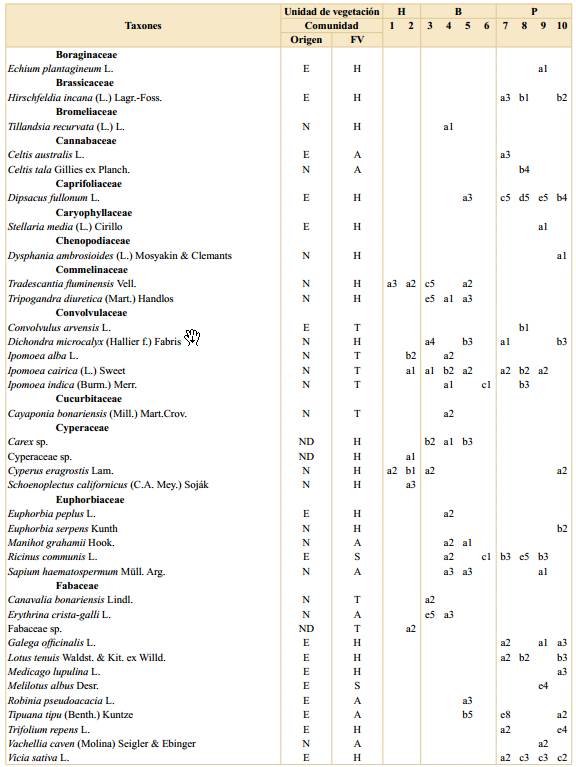
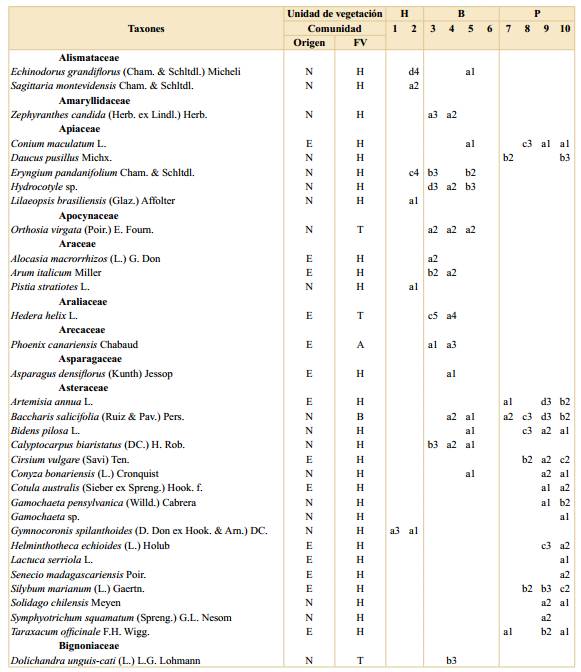
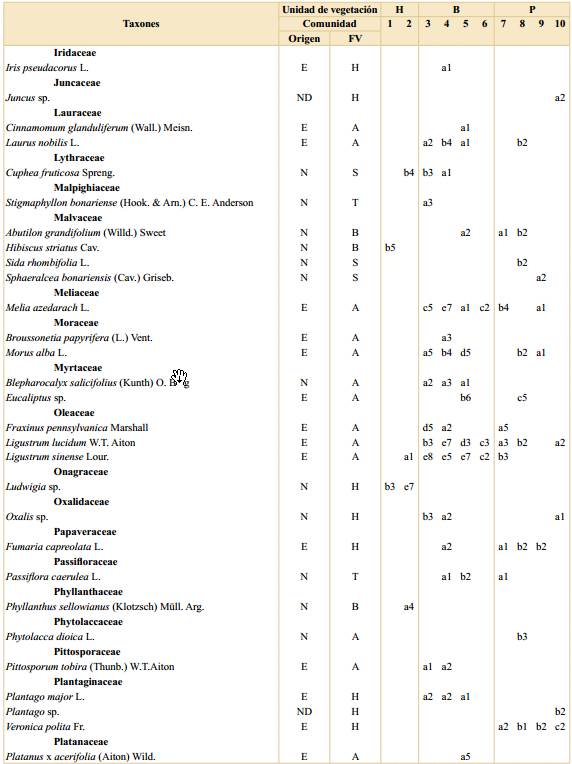
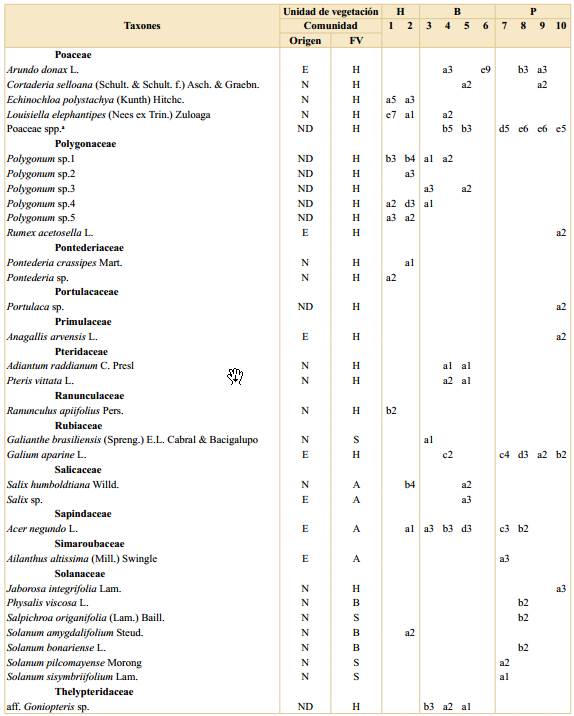
a Dentro de Poaceae spp. se identifcaron las especies nativas Bromus catharticus Vahl, Paspalum notatum Flüggé y Paspalum urvillei Steud. y las exóticas Cynodon dactylon (L.) Pers., Lolium sp., Megathyrsus maximus (Jacq.) B.K. Simon & S.W.L. Jacobs, Poa annua L., Polypogon sp., Setaria sp., Sorghum halepense (L.) Pers. Otros cinco morfotipos de gramíneas sólo se identifcaron a nivel de familia.
Unidades de vegetación y comunidades de la RECU-CN
Los ejes DCA1 y DCA2 tuvieron autovalores de 0,9 y 0,4 y explicaron el 6% y el 3% de la variabilidad de los datos respectivamente (Fig. 2). En el extremo positivo del eje DCA1 se observó un conglomerado de parcelas que pertenecen a la porción anegada del área de estudio, ubicadas entre el murallón de contención de Ciudad Universitaria y la porción elevada del terreno que corresponde a la península de relleno y, también, sobre la porción inundable costera del área. En el sector central del ordenamiento se encontraron las parcelas ubicadas sobre la península, desde la orilla inundable hasta el centro de la misma, donde el terreno pasa a ser más elevado y disminuye la posibilidad de ingreso de agua ante eventos extraordinarios de sudestada. En el extremo negativo del eje DCA1 se agruparon las parcelas correspondientes a la porción de la península más cercana al río, cuyos rellenos son más elevados que en la porción antedicha, y se encuentran completamente aislados de la dinámica estuarial, siendo a su vez la porción más antropizada. De esta forma se identificó en forma indirecta un gradiente ambiental de inundación hacia la derecha de la figura y, en sentido opuesto al anterior, un gradiente de antropización que incluye tanto la existencia de un sustrato antrópico, como la de disturbios producidos por el humano (Fig. 2).
El nivel de corte más frecuente de todos los considerados en el análisis de clasificación indicó la formación de tres grandes grupos de parcelas (distintos trazos en la figura 2). Con base en esta clasificación se definieron las unidades de vegetación “Humedal”, “Bosque” y “Pastizal con arboledas”, con 15, 29 y 33 parcelas respectivamente. El segundo nivel de corte indicó la formación de 10 grupos (Fig. 2) que definieron las 10 comunidades de plantas de entre dos y 16 parcelas cada uno. La formación de estos 10 grupos de parcelas se dio en un contexto de variabilidad explicada de los ejes del ordenamiento cercana al 10 % y son gráficamente menos evidentes que los tres grupos correspondientes a las unidades de vegetación (Fig. 2). Tanto las unidades de vegetación como las comunidades fueron denominadas en función de sus formas de vida y especies dominantes y se describen a continuación.
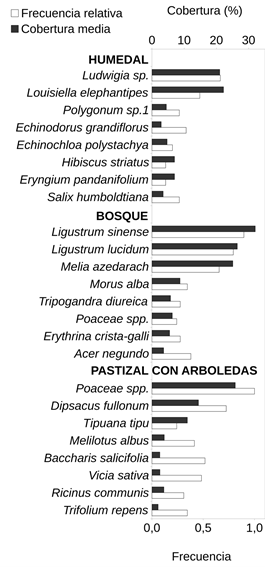
Humedal. Esta unidad se ubicó principalmente entre el murallón de contención de Ciudad Universitaria y la península, con una parcela sobre la costa (Fig. 1). Esta unidad de vegetación tuvo el menor número de especies totales (32) con un promedio de cinco especies por parcela. Por otro lado, tanto el porcentaje de número de especies nativas (90%) como el de cobertura de dichas especies (99%) resultaron ser mayores que en las otras unidades. Esta unidad de vegetación presentó mayor cobertura de especies herbáceas (77%), siendo las más abundantes y frecuentes Ludwigia sp. y Louisiella elephantipes (Fig. 3). Otras especies de herbáceas registradas con coberturas y frecuencias intermedias resultaron ser distintas especies de Polygonum, Echinodorus grandiflorus, Echinochloa polystachya y Eryngium pandanifolium; también se registraron especies arbustivas y arbóreas como Hibiscus striatus y Salix humboldtiana respectivamente (Fig. 3).
Con una cobertura intermedia, pero una baja frecuencia se registró Schoenoplectus californicus, mientras que por el contrario la trepadora Ipomoea alba y la herbácea Cyperus eragrostis se registraron en una frecuencia intermedia, pero con baja cobertura (Tabla 1). En esta unidad se identificaron las comunidades (1) Canutillar de Louisiella elephantipes y (2) Verdolagal de Ludwigia sp. con un total de 7 y 8 parcelas respectivamente. La composición de especies de cada comunidad se detalla en la Tabla 1.
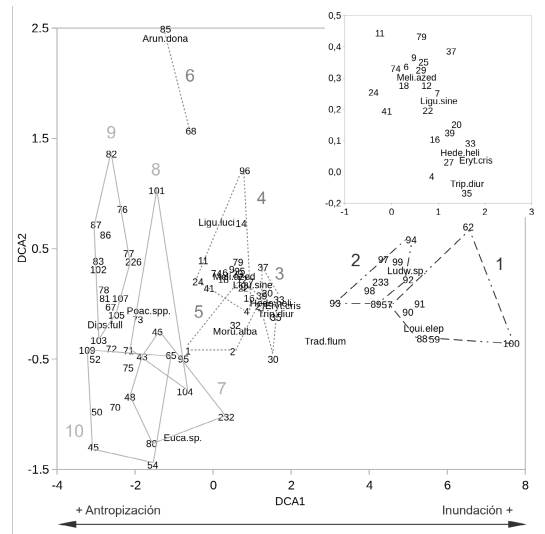
Fig. 2 Ordenamiento y clasificación de la vegetación de la RECU-CN. En el ordenamiento se indican las 15 especies de mayor cobertura (letras), las parcelas de muestreo (números pequeños), una ampliación de la zona central congestionada (esquina superior derecha) y el gradiente ambiental identificado (flecha abajo). En la clasificación se indican los grupos de parcelas correspondientes a las tres unidades de vegetación (Humedal, líneas de rayas y puntos; Bosque, líneas punteadas; Pastizal con arboledas, líneas continuas) y a las 10 comunidades vegetales (números grandes, ver figura 1). Arun.dona = Arundo donax, Dips.full = Dipsacus fullonum, Eryt.cris = Erythrina crista-galli, Euca.sp. = Eucalyptus sp., Hede.heli = Hedera helix, Ligu.luci = Ligustrum lucidum, Ligu.sine = Ligustrum sinense, Loui.elep = Louisiella elephantipes, Ludw.sp. = Ludwigia sp., Meli.azed = Melia azedarach, Moru.alba = Morus alba, Poac.spp. = Poaceae spp., Tipu.tipu = Tipuana tipu, Trad.flum = Tradescantia fluminensis, Trip.diur = Tripogranda diuretica.
Bosque. Esta segunda unidad de vegetación se ubicó en la porción central del área (Fig. 1). Se registraron 77 especies con un promedio de 10 especies por parcela, de las cuales el 55% correspondieron a especies nativas de la región con una cobertura total del 18%. En promedio las parcelas se encontraron cubiertas en un 72% por árboles. Las especies que se encontraron con mayor cobertura y frecuencia fueron Ligustrum sinense, Ligustrum lucidum y Melia azedarach, dominando sobre otras especies arbóreas como Morus alba, Erythrina crista-galli y Acer negundo (Fig. 3). Además, se registraron hierbas como Tripogandra diuretica y gramíneas (Poaceae spp.). En esta unidad se identificaron las comunidades (3) Bosque de ceibo y ligustrina, (4) Bosque de ligustro y paraíso, (5) Bosque de eucalipto y ligustrina y (6) Cañaveral de caña de Castilla, con un total de 5, 16, 6 y 2 parcelas respectivamente. La composición de especies de cada comunidad se detalla en la Tabla 1.
Pastizal con arboledas. Esta tercera unidad de vegetación se ubicó en la península hacia el río, ocupando la superficie ovalada rodeada por cemento y las zonas lindantes que limitan con la franja boscosa (Fig. 1). Se registraron 77 especies en total y aproximadamente 9 por parcela. Esta unidad de vegetación presentó el menor porcentaje de número de especies nativas de las tres (43%) y de cobertura total correspondiente a este grupo de especies (11%). La forma de vida predominante, con un 66% de cobertura, correspondió a las herbáceas, quedando el restante 34% mayormente comprendido por árboles. Esta unidad de vegetación se encontró dominada, tanto en cobertura como en frecuencia, por diversas especies de gramíneas (Poaceae spp.), como Megathyrsus maximus y Sorghum halepense, acompañadas principalmente por Dipsacus fullonum (Fig. 3). Otras herbáceas fueron registradas en una frecuencia intermedia, pero con coberturas menores, como por ejemplo Melilotus albus, Vicia sativa y Trifolium repens. En cuanto a otras formas de vida se registraron considerables cobertura y frecuencia de la especie arbórea Tipuana tipu y los arbustos Baccharis salicifolia y Ricinus communis. En esta unidad se identificaron las comunidades (7) Arboleda de tipa, (8) Pastizal con ricino, (9) Pastizal con carda y (10) Pastizal mixto, con un total de 6, 4, 14 y 9 parcelas respectivamente. La composición de especies de cada comunidad se detalla en la Tabla 1.
DISCUSIÓN Y CONCLUSIONES
Riqueza de plantas vasculares de la RECU-CN En este trabajo se describió la vegetación de la Reserva Ecológica Ciudad Universitaria - Costanera Norte (RECU-CN), encontrándose una riqueza global de 154 especies, las cuales representan el 65% de las especies totales que habría en el área de estudio según la estimación realizada (238 especies). Esta diferencia puede deberse a que el muestreo se realizó durante la temporada de invierno, pudiendo resultar en una subestimación de especies al pasar desapercibidas aquellas especies anuales que podrían ser reconocidas en primavera y verano. La subestimación tampoco resulta sorpresiva ya que durante la realización del trabajo de campo se observaron especies que no se registraron dentro de ninguna parcela, como por ejemplo Pontederia cordata y Enterolobium contortisiliquum. A su vez, el valor estimado de especies totales del área fue de 238, número cercano a lo registrado colectivamente y de forma histórica para el área, donde se alcanzó un total de 263 especies (Sirolli et al., 2018). De las 58 familias registradas, aquellas con más representantes resultaron ser Poaceae y Asteraceae, lo cual concuerda con lo registrado por Burkart (1957) para el delta del río Paraná y por Faggi & Cagnoni (1990) para la costa de la ciudad de Buenos Aires.
En comparación con otras áreas naturales costeras cercanas, relevadas con una metodología similar (Faggi & Cagnoni, 1987; Kalesnik et al., 2005), la riqueza global registrada en la RECU-CN fue intermedia respecto a las 104 especies registradas en el Refugio Natural Educativo Ribera Norte (RN) y a las 242 especies registradas en la Reserva Ecológica Costanera Sur (RECS). Considerando la superficie total de estas tres áreas (23, 10 y 350 ha respectivamente), la riqueza de plantas vasculares de la RECU-CN estuvo en el rango de lo esperado según la curva clásica de relación entre número de especies y área (Arrhenius, 1921). Por el contrario, el porcentaje de especies exóticas registradas en la RECU-CN (42%) fue mucho mayor que en las otras dos áreas mencionadas (RN 14%, RECS 27%). Dicho porcentaje guardó la misma relación que el porcentaje de sustrato antrópico constituido por la península en la RECU-CN, por los terraplenes en la RECS y casi nula en el RN (Faggi & Cagnoni, 1987, 1990; Kalesnik et al., 2005; Sirolli et al., 2018). Si se considera que en las tres áreas se encontró mayor proporción de exóticas en dichos sustratos, que coinciden con las posiciones topográficamente más altas y transitadas, entonces los resultados son consistentes con que la actividad humana favorece la colonización por especies exóticas generando condiciones más secas en contextos ribereños y ayudando a la dispersión de especies exóticas, especialmente aquellas de ambientes terrestres (Catford et al., 2011; Jansen et al., 2011).
Unidades de vegetación y comunidades de la RECU-CN
La vegetación de la RECU-CN se ordenó a lo largo de dos gradientes principales: inundabilidad, creciente de izquierda a derecha, y antropización, creciente de derecha a izquierda (Fig. 2). Ambos factores son considerados estructuradores de la vegetación de suma relevancia en contextos ribereños (Webb et al., 2012; Osterkamp & Hupp, 2010; You et al., 2015). Este resultado se reflejó no sólo a escala de las tres unidades de vegetación sino también dentro de las mismas, donde se diferenciaron comunidades de especies más o menos tolerantes al anegamiento, por ejemplo dentro del Humedal se diferenciaron el Canutillar de Louisiella elephantipes del Verdolagal de Ludwigia sp. y, dentro del Bosque, el Bosque de ceibo y ligustrina versus el Bosque de ligustro y paraíso. En un extremo, los sectores más bajos e inundables coinciden con los sectores menos transitados y disturbados por el ser humano y, en el otro, los sectores más altos poseen un sustrato de origen artificial y mayores frecuencia e intensidad de disturbios antrópicos (Sirolli et al., 2018). Estas condiciones también explican las principales diferencias fisionómicas de las unidades de vegetación, que se intercalan espacialmente entre fisonomías herbáceas y arbóreas con relativo equilibrio en extensión.
Las comunidades registradas en la unidad de Humedal constituyen fisonomías y elementos ampliamente reconocidos en la bibliografía del río Paraná, su delta y la ribera platense (Burkart, 1957; Neiff, 1986; Cabrera & Zardini, 1993; Kalesnik et al., 2005; Sabattini & Lallana, 2007; Passarelli et al., 2014). Según dichos autores, Louisiella elephantipes es una especie de gramínea flotante asociada a corrientes bajas a moderadas que crece con gran desarrollo en márgenes, marismas, lagunas y otros ambientes similares a donde se la registró en la RECU- CN. Por su lado, Ludwigia sp. es una herbácea arraigada que se describe creciendo en terrenos inundables internos y costeros, como se observó en la RECU-CN. Además, la parcela ubicada sobre la costa (233) presentó gran cobertura de Phyllanthus sellowianus, una especie característica del matorral ribereño (Burkart, 1957; Cabrera & Zardini, 1993). A pesar de no haberse registrado dentro de las parcelas realizadas en la unidad de Humedal, durante el muestreo también se observaron espadañales de Zizaniopsis bonariensis, totorales de Typha latifolia y juncales de Schoenoplectus californicus, todas comunidades también características de la región (Burkart, 1957; Passarelli et al., 2014). Las especies exóticas fueron escasas en esta unidad, sin embargo, a pesar de haberse registrado con baja cobertura y frecuencia, es destacable mencionar la presencia de Iris pseudacorus debido a su gran capacidad invasora (Kandus & Quintana, 2018; Chen, 2019). De esta forma, las comunidades registradas presentaron una combinación de especies muy similar a la descrita por bibliografía en los ambientes naturales y con casi nula presencia de especies exóticas, en contraposición con la hipótesis de trabajo planteada.
En cuanto a la unidad de vegetación de Bosque, a pesar de la alternancia de la especie dominante en el dosel, se destacó la abundancia de Ligustrum sinense en el estrato medio. Esta especie asiática constituye una invasora exitosa en la región, que se desarrolla en etapas tempranas a intermedias en la sucesión (Kalesnik et al., 2005; Kalesnik & Aceñolaza, 2008; Sirolli & Kalesnik, 2015). Entre las causas de dicho éxito se mencionan la dispersión por aves durante la colonización inicial del área, la gran producción de frutos y la formación de un importante banco de semillas y de plántulas (Montaldo, 1993; Kalesnik et al., 2013). A su vez, se observó una baja cobertura de especies nativas (18%) que, a diferencia de las comunidades del Humedal, propone una composición novedosa en comparación con sus contrapartes descritas en la bibliografía para ambientes naturales de la región pudiéndose considerar, en consecuencia, neoecosistemas.
La comunidad de Bosque de ceibo y ligustrina fue la única que se asemejó a una de las formaciones que se describen en la bibliografía de la región: el ceibal (Burkart, 1957; Cabrera & Zardini, 1993). El dosel de esta comunidad estuvo dominado por Erythrina crista-galli y se registraron algunos componentes en común como las enredaderas Passiflora caerulea y Cissus palmata. Sin embargo, el estrato medio dominado por Ligustrum sinense y la ausencia de un sotobosque de ciperáceas marcan una importante diferencia explicada, probablemente, por el rápido cambio topográfico impuesto por la península, que determina baja superficie inundable. Por su lado, las especies dominantes del dosel de la comunidad de Bosque de ligustro y paraíso representan dos de las invasoras más emblemáticas de la región (Söyrinki, 1991; Ghersa et al., 2002; Torresín et al., 2013; Franco et al., 2018). A diferencia de su congénere, L. lucidum es más tolerante a la sombra, aunque comparte su elevado éxito reproductivo, explicado en parte por el bajo daño foliar que presenta en esta región, donde no es nativa (Montaldo, 2000; Montti et al., 2016). En cuanto al Bosque de eucalipto y ligustrina, si bien en Argentina se mencionan algunas especies naturalizadas del género Eucalyptus (Scaramuzzino et al., 2019), su crecimiento espontáneo y capacidad de invasión son limitados. En este caso, probablemente se trate de ejemplares plantados, ya que actualmente poseen un patrón poblacional remanente, dada la ausencia de plántulas registrada durante los muestreos. Por su lado, Arundo donax, la especie que domina la comunidad de Cañaveral de caña de Castilla, también es una especie exótica naturalizada en la costa rioplatense que forma parches densos cerca de arroyos, caminos y vías férreas (Söyrinki, 1991; Passarelli et al., 2014). El hecho de que esta comunidad haya sido clasificada dentro de la unidad de Bosque responde a su ubicación transicional entre esta unidad y la de Pastizal con arboledas, en donde algunas ramas de ligustro, ligustrina y paraíso se registraron dentro de las parcelas de cañaveral. El distanciamiento de estas parcelas respecto a las otras (números 68 y 85 en la figura 2), indican una composición considerablemente diferente a ambas unidades.
En la unidad de vegetación de Pastizal con arboledas predominaron comunidades fisonómicamente similares a la estepa graminosa que cubría la mayor parte de la región (Cabrera & Zardini, 1993; Paruelo et al., 2007). Sin embargo, su composición fue notablemente diferente, dominando especies de gramíneas no nativas (Burkart et al., 2011; Andrade et al., 2018), por lo que pueden considerarse neoecosistemas, de forma similar a lo discutido para las comunidades de Bosque. Las comunidades se diferenciaron en función de sus especies acompañantes: Ricinus communis, Dipsacus fullonum y Trifolium repens. Se trata de especies exóticas invasoras que crecen en terrenos productivos, artificiales o recientemente disturbados (Söyrinki, 1991; Sirolli & Kalesnik, 2011; Busso et al., 2013). La Arboleda de tipa tuvo presencia de gramíneas en su sotobosque, por lo que quedó clasificada junto a los pastizales aunque su composición se aproximó también a la unidad de Bosque (comunidad 7, Fig. 2). Su especie dominante, Tipuana tipu, es nativa del noroeste de Argentina y constituye una especie común en el arbolado de la ciudad, crece en forma subespontánea en la región y es considerada invasora en otros lugares donde no es nativa (Valla et al., 1999; Sampaio & Schmidt, 2014; Shabani et al., 2020). Fuera de las parcelas de muestreo también se observaron arboledas dominadas por otras especies ya mencionadas para la unidad de Bosque, como Eucalyptus sp. Estas arboledas se corresponden con lo tratado por la FAO como árboles fuera del bosque (de Foresta et al., 2013).
En conclusión, a pesar de tratarse de un relevamiento invernal, los resultados permitieron registrar el 59% del total de especies registradas históricamente para el área (Sirolli et al., 2018) y distinguir que las unidades de vegetación características de la RECU-CN se distribuyen entre Humedal, Bosque y Pastizal con arboledas. Las superficies hormigonadas son considerables y también pudieron ser detectadas mediante el muestreo. Con mayor detalle, pudieron registrarse y caracterizarse algunas de las comunidades que pueden observarse en campo, encontrándose composiciones similares a las descritas en la bibliografía de la región sólo para la unidad de vegetación de Humedal, mientras que las comunidades de Bosque y de Pastizal con arboledas presentaron características propias de los neoecosistemas. Por lo analizado en el presente trabajo, es relevante considerar en el manejo del área los regímenes de inundación y de disturbios antrópicos, inclusive teniendo en cuenta que la ocurrencia de algunos de estos últimos puede determinar la permanencia de la unidad de Pastizal con arboledas. Por lo tanto, se destaca como principales valores de conservación la coexistencia de unidades de vegetación fisonómicamente diversas y relativamente equilibradas en extensión, así como también, la presencia de comunidades y especies típicas de la región dentro de la unidad de Humedal de la RECU-CN. Por último, el enriquecimiento con especies nativas en la península de relleno aumentará considerablemente dicho valor de conservación (Dearborn & Kark, 2010; Holl, 2020).

