











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Las sociedades cazadoras recolectoras que habitaron los litorales de ambientes acuáticos marinos establecieron distintas interacciones con el entorno. Los cuerpos de agua desempeñaron un importante papel dada la diversidad de sus componentes bióticos, y el litoral en sí mismo constituyó un ecotono con ambientes marinos y terrestres del interior. Además, estos paisajes conformaron vías de comunicación y transporte de grupos humanos, ideas y valoraciones simbólicas (Orquera & Gómez Otero, 2007; Zubimendi & Ambrústolo, 2016; Prates & Bonomo, 2017). El litoral atlántico de la Patagonia (LAP) ha estado ampliamente ocupado, desde hace al menos 8000 años AP en el continente y 5700 AP en la Isla Grande de Tierra del Fuego (fechados sobre valvas, no corregidos por efecto reservorio, Favier Dubois & Borrero, 2005; Gómez Otero et al., 2013; Zubimendi et al., 2015). En este proceso de exploración y ocupación del territorio las sociedades cazadoras recolectoras habitaron ambientes en continua transformación producto de procesos geomorfológicos y paleoclimáticos (Cruz et al., 2006). Los grupos humanos realizaron un uso sostenido de algunos sectores del LAP a lo largo del tiempo, así como para el Holoceno tardío una utilización intensiva de los recursos marinos, tal es el caso de la costa norte de Santa Cruz (CNSC) y la costa este y sur de Tierra del Fuego (Borrero, 1985; Borrero & Lanata, 1988; Borrero et al., 2006; Zubimendi et al., 2011; Vázquez et al., 2013).
Los estudios arqueobotánicos realizados hasta la fecha han revelado, mediante análisis antracológicos, carpológicos y polínicos, aspectos relevantes del uso de los recursos vegetales en el LAP (Marconetto, 1996, 2002; Piqué, 1999; Capparelli et al., 2009; Ortega & Marconetto, 2009, 2012; Berihuete, 2010; Ortega, 2012; Ciampagna, 2015, 2016; Ciampagna et al., 2016; Caruso Fermé et al., 2017, 2019; Franch et al., 2021). Estos análisis se han desarrollado, por un lado, en el litoral de la provincia de Río Negro y Chubut en áreas puntuales tales como el Golfo San Matías (Río Negro) y el Golfo Nuevo (Chubut). En el Golfo San Matías, se analizaron los restos leñosos carbonizados de cuatro sitios concheros de la costa norte y se compararon los resultados con los obtenidos en perfiles polínicos (Ortega & Marconetto, 2009, 2012; Marcos & Ortega, 2014). En el Golfo Nuevo (Península Valdés, Chubut) se desarrollaron análisis de carbones procedentes del sitio San Pablo 6 (Caruso et al., 2017). Por otro lado, se llevaron a cabo estudios antracológicos en el extremo sur de Patagonia, más precisamente en el área litoral de Santa Cruz y de Tierra del Fuego. En la primera, se han estudiado varios sitios arqueológicos costeros de los cuales se analizaron gran variedad de restos vegetales como son los carbones, frutos, hojas y microrrestos procedentes de artefactos de molienda (Ciampagna, 2015, 2016, 2018; Ciampagna et al., 2016, 2020, 2021). Para su interpretación se generó un modelo de uso de plantas por los grupos humanos sobre la base de un exhaustivo estudio de fuentes etnohistóricas de Patagonia (Ciampagna & Capparelli, 2012), mientras que para el litoral atlántico de Tierra del Fuego los estudios son más recientes, y se han realizado exclusivamente sobre restos antracológicos y carpológicos del sitio CSP2017 (Franch et al., 2021). Fuera del litoral atlántico, existen varios análisis arqueobotánicos sobre carbones, frutos y semillas, que permitieron caracterizar la utilización de distintos tipos de recursos vegetales en el archipiélago fueguino, tal como se describe a continuación. Para los sitios costeros del Canal de Beagle, los trabajos de Piqué (1999) evalúan las estrategias de gestión del combustible mediante la identificación de los recursos leñosos de cinco sitios concheros: Túnel VII, Lanashuaia, Alashawaia, Shamakush I y X. Caruso Fermé et al. (2017, 2019) estudian el uso de los recursos leñosos empleados como combustible de Imiwaia I y Heskaia 35. También Berihuete (2010) analiza en su Tesis Doctoral restos de semillas y frutos de diferentes sitios arqueológicos de la costa norte del canal Beagle, entre ellos los yacimientos de Lanashuaia y Túnel VII. A estos trabajos se suman varios estudios antracológicos de la zona litoral del Pacífico chileno (Solari, 1988, 1992, 2003, 2007; Camus & Solari, 2008; Solari & Lehnebach, 2010).
Los estudios arqueobotánicos de ambientes litorales mencionados muestran un amplio espectro de prácticas de uso y procesamiento de vegetales, que involucran la gestión de las estepas arbustivas y de los bosques de Nothofagus spp. según estén presentes en cada área. Entre estas prácticas se destaca el uso de combustibles -donde se detectan distintos tipos de fuegos-, el uso de maderas como artefactos, la obtención de alimentos a partir del consumo fresco de frutos, la cocción de órganos subterráneos y la molienda de frutos tanto de carácter alimenticio como aquellos utilizados potencialmente como medicinas. En este marco, la asociación del registro vegetal con otros materiales del conjunto arqueológico, tales como instrumental óseo, lítico y malacológico, puede contribuir a comprender aún más los comportamientos de los grupos cazadores recolectores en la interacción con el ambiente acuático y el ecotono con el ambiente terrestre (sensu Prates & Bonomo, 2017).
La madera carbonizada es uno de las macrorrestos vegetales más frecuentes en los sitios arqueológicos y permite estudiar las plantas leñosas utilizadas por los grupos humanos (Buxó & Piqué, 2008). Los carbones -entendidos aquí como derivados de material leñoso-, al igual que la mayoría de los especímenes arqueobotánicos, permiten un estudio dual, ya que son productos de la relación dialéctica entre las comunidades humanas y su entorno. Por lo tanto, son a la vez testigos directos del ambiente regional y restos materiales de la actividad antrópica realizada en los sitios de ocupación (Piqué, 1999). Aunque la recolección de leños ha estado condicionada por factores ecológicos (como la distribución espacial y disponibilidad, entre otros), es la acción humana la que determina sus estrategias de gestión y aprovisionamiento (Buxó & Piqué, 2008), las cuales dependen de los procesos de producción y reproducción de la comunidad (Piqué, 1999). La conformación y descripción de una exhaustiva colección de referencia de leños, entonces, es indispensable al momento de abordar el estudio de los restos materiales de este tipo en los sitios arqueológicos por varios motivos. Por un lado, brinda un amplio espectro taxonómico de la flora potencialmente presente en el pasado regional y las características morfoanatómicas que permiten su reconocimiento. Por otro lado, permite inferir el tipo de asociación entre las maderas y las prácticas de recolección y el manejo de arbustos y de árboles en la estepa y en el bosque en tiempos pasados (Andreoni, 2010).
Por todo lo expuesto, este trabajo tiene por objetivo, en primer lugar, presentar y caracterizar anatómicamente la colección de referencia de maderas potencialmente utilizadas por las sociedades cazadoras recolectoras del litoral atlántico de Patagonia sur (LAPS), donde se incluye territorio tanto continental (Santa Cruz), como insular (Tierra del Fuego). En segundo lugar, generar una clave dicotómica que facilite los estudios antracológicos en la región. Por último, esta investigación propone dos casos de aplicación de los resultados obtenidos, incluida la puesta a prueba de la clave dicotómica en la identificación de carbones arqueológicos provenientes de los sitios del Holoceno tardío final tales como Palo Alto (CNSC, Santa Cruz) y Cabo San Pablo 2017 (Isla Grande de Tierra del Fuego) (Fig. 1). El primer sitio brinda información original y el segundo se hace sobre la base de datos recientemente publicados (Franch et al., 2020). Además de la creación de un corpus de información a nivel regional que favorezca los análisis de carbones, el trabajo es particularmente relevante por el aporte original de descripciones anatómicas de especies vegetales de las que, por no poseer uso comercial en la actualidad, se conoce muy poco, especialmente en lo que atañe a los arbustos de porte pequeño.
Fitogeografía del LAPS y vegetación del Holoceno tardío con base en análisis polínicos Provincia de Santa Cruz
La zona litoral presenta dos unidades de vegetación de mayor extensión: una estepa arbustiva con arbustos de 3 m de altura o más, denominada “Estepa arbustiva del Golfo San Jorge”, que se extiende de norte a sur por una franja costera de 90 a 25 km de ancho hasta Punta Medanosa, y una estepa de arbustos enanos o rastreros, que se extiende sobre la meseta central de Santa Cruz denominada en forma homónima “Meseta central” (Fig. 1, Oliva et al., 2001). La Estepa arbustiva del Golfo San Jorge se extiende sobre un suelo ondulado surcado por cañadones y valles que desembocan en el mar (Oliva et al., 2001). Entre las especies dominantes se encuentran los arbustos de Retanilla patagonica (Speg.) Tortosa (malaespina), Colliguaja integerrima Gillies. & Hook. (duraznillo), Azorella prolifera (Cav.) G.M. Plunkett & A.N. Nicolas (=Mulinum spinosum (Cav.) Pers; neneo), Mulguraea ligustrina (Lag.) N. O´Leary & P. Peralta var. lorentzii (Niederl. ex Hieron.) N. O´Leary & P. Peralta (mata negra), Ephedra ochreata Miers. (solupe), Lycium chilense Miers. ex Bertero (yaoyín) y Anarthrophyllum rigidum (Gillies ex Hook. & Arn.) Hieron. (mata amarilla) (Oliva et al., 2001). Se relevó Lepidophyllum cupressiforme (Lam.) Cass. en el distrito del Golfo San Jorge (Obs. pers. de Ciampagna y Capparelli). Entre las gramíneas se encuentran Pappostipa humilis (Cav.) Romasch. var. humilis (coirón amargo) y Festuca argentina (Speg.) Parodi (huecú). También se registran plantas anuales tales como Lepidium sp., Erodium cicutarium (L.) L´Hér. ex Aiton (alfilerillo) y Vulpia sp. Es importante mencionar que, aunque la provincia fitogeográfica del Monte se extiende hasta unos kilómetros al sur del paralelo 44°, es decir hasta el centro-este de la Provincia de Chubut (Morello, 1958), la Estepa arbustiva del Golfo San Jorge facilita la distribución extrema austral de algunos elementos del monte, entre los que es posible citar: Nassella tenuis (Phill.) Barkworth, Prosopis denudans Benth., Prosopidastrum globosum (Gillies ex Hook. & Arn.) Burkart, Bounganvillea spinosa (Cav.) Heimerl, Schinus johnstonii F.A. Barkley, Larrea ameghinoi Speg. y Lycium ameghinoi Speg. (León et al., 1998). Se consideró también en este último caso a Erythrostemon gilliesi (Wall ex Hook) Klotzsch var. gilliesi y como parte de la estepa del Distrito occidental a Fabiana imbricata Ruiz & Pav. Retuer y Bertolami (2010) observan que en los matorrales de los cañadones costeros se desarrollan tres comunidades de vegetación y mencionan, además de las especies anteriores, a Chuquiraga avellanedae Lorentz, Adesmia salamancensis Burkart, Senecio filaginoides DC., Atriplex lampa (Moq.) D. Dietr. y Suaeda divaricata Moq. Por otra parte, el “Matorral de Mata Negra”, la “Estepa magellánica seca” y la “Estepa magellánica húmeda” son unidades de vegetación que ocupan menor extensión en la franja litoral. En la primera predominan especies leñosas tales como Mulguraea tridens (Lag.) O`Leary & P. Peralta, Nardophyllum obtusifolium Hook. & Arn., Berberis microphylla G. Forst., Nassauvia glomerulosa (Lag. ex Lindl.) D. Don, Nassauvia ulicina (Hook. F.) Macloskie, Clinopodium darwinii (Benth.) Kuntze y Ephedra frustillata Miers. En la segunda, se encuentran Nardophyllum bryoides (Lam.) Cabrera, Nassauvia ulicina, Nassauvia fuegiana (Speg.) Cabrera, Perezia recurvata (Vahl) Less., Ephedra frustillata, Berberis microphylla y Mulguraea tridens. Mientras que en la última unidad dominan Chiliotrichum diffusum (G. Forst.) Kuntze, Berberis microphylla, Empetrum rubrum Vahl ex Willd., Baccharis nivalis (Wedd.) Sch. Bip. ex Phill, Nassauvia fuegiana, Azorella fuegiana Speg., Nassauvia aculeata (Less.) Poepp. & Endl. y Perezia recurvata (Oliva et al., 2001). Estudios polínicos demuestran que la vegetación del área mencionada durante el Holoceno tardío sería análoga a la actual (De Porras, 2010).
Provincia de Tierra del Fuego
Particularmente se hará referencia a la Isla Grande de Tierra del Fuego, la cual presenta diversos ecosistemas que se desarrollan en cinco grandes unidades de vegetación: la “Estepa magellánica húmeda”, el “Ecotono bosque-estepa”, el “Bosque caducifolio”, el “Bosque mixto” y el “Bosque lluvioso o perennifolio” (Tuhkanen, 1992) (Fig. 1). La estepa se desarrolla al norte de la Isla Grande y se caracteriza por llanuras onduladas de herbáceas dominadas por Festuca gracillima Hook. f. Le sigue hacia el sur el Ecotono bosque-estepa, con comunidades intermedias entre la estepa y el bosque, donde los pastizales dan paso a formaciones arbustivas dominadas por Chiliotrichum diffusum (mata negra), mientras que en las depresiones se desarrollan praderas o vegas (Tuhkanen, 1992). La especie arbórea dominante de este ecotono es Nothofagus antarctica (G. Forst.) Oerst. (ñire), mientras que en los lugares más altos y cerrados se encuentra Nothofagus pumilio (Poepp. & Endl.) Krasser (lenga), ambas especies caducifolias (Moore, 1983; Tuhkanen, 1992).
El Bosque caducifolio se extiende principalmente al norte y este del lago Fagnano, está compuesto por comunidades de Nothofagus pumilio y de Nothofagus antarctica en formaciones monoespecíficas. Los límites entre ambas comunidades boscosas son netos y contrastantes, con una transición de escasos metros. Las formaciones arbustivas aparecen en los límites de las masas boscosas y están compuestas por matorrales de Chiliotrichum diffusum, Berberis microphylla (calafate) y Empetrum rubrum (murtilla) (Moore, 1983; Tuhkanen, 1992). El Bosque mixto se extiende desde la costa sur de la Isla Grande hasta la costa sur del lago Fagnano. Estos bosques están caracterizados por la dominancia de Nothofagus pumilio y de Nothofagus betuloides (Mirb.) Oerst. (guindo). Por debajo de las copas de Nothofagus se identifican Drimys winteri J.R. Forst. & G. Forst. (canelo) y, en los bosques secos, también Maytenus magellanica (Lam.) Hook. f. (leña dura). Las formaciones arbustivas están compuestas por matorrales de Chiliotrichum diffusum, Berberis microphylla, Berberis ilicifolia L. f. (michay), Gaultheria mucronata (L. f.) Hook. & Arn. (chaura) y Empetrum rubrum (Moore, 1983; Tuhkanen, 1992). El Bosque perennifolio es una formación que se encuentra en una pequeña área en el extremo sudeste de la Isla Grande, en tanto se desarrolla principalmente en el interior del archipiélago del Pacífico, Isla de los Estados y Península Mitre. Se trata de un bosque denso compuesto principalmente por Nothofagus betuloides, junto con Drimys winteri, Maytenus magellanica y Embothrium coccineum J.R. Forst. & G. Forst. (notro). El estrato arbustivo está representado por Escallonia serrata Sm., Berberis ilicifolia y Gaultheria mucronata (chaura) (Tuhkanen, 1992).
Hacia 3000 años AP ya se encontrarían establecidas las condiciones climáticas actuales. El principal cambio registrado en el polen a lo largo del Holoceno tardío es la disminución de Nothofagus sp. y un aumento de los pastizales después del 1000 cal. AP, que se intensifica entre los 680 y 300 cal. AP. Esto fue causado por condiciones frescas y húmedas que coincidieron con la Pequeña Edad de Hielo (LIA) en el hemisferio sur (Borromei et al., 2009; Coronato, 2014). Estos datos indican que, durante la ocupación de los sitios analizados en este trabajo, el bosque de Nothofagus estaba plenamente instalado.
Casos de Aplicación: sitios arqueológicos del LAPS
Provincia de Santa Cruz. El sitio conchero Palo Alto se ubica en la costa norte de la provincia (Fig. 1), en el primer escalón del flanco de meseta (Fig. 2A, B), a 60 m de la línea de marea y a 15 m sobre el nivel del mar, en la localidad arqueológica de Bahía Lángara sobre la costa central del Golfo San Jorge. El fechado sobre valvas de Nacella magellanica lo ubica en los 690 ± 90 años AP (LP-2280) (Zubimendi et al., 2010). Es un sitio de 50 x 25 m en el que se excavaron 4,25 m2 (Fig. 2C), siguiendo niveles artificiales de 5 cm en un primer sondeo y de 10 cm hasta los 27 cm de profundidad respecto del nivel 0.
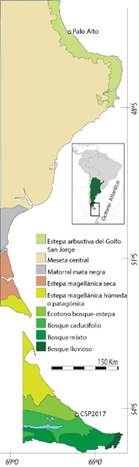
Fig. 1 Mapa con la ubicación de los sitios y la fitogeografía del sector litoral de las provincias de Santa Cruz y Tierra del Fuego (tomado de Oliva et al., 2001 y Tuhkanen, 1992). Figura en color en la versión en línea http://www.ojs.darwin.edu.ar/index.php/darwiniana/ article/view/1020/1254
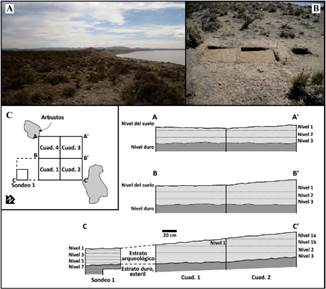
Fig. 2 A, estepa arbustiva del Golfo San Jorge en los alrededores del sitio Palo Alto. B, vista de la excavación. C, planos de excavación y estratigrafía del sitio Palo Alto (CNSC, tomado de Zubimendi et al., 2010). Figura en color en la versión en línea http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/1020/1254
Allí se observó un sedimento consolidado sin restos arqueológicos que corresponde a un estrato terciario del Patagoniano (Fig. 2). A excepción de esta última capa, el sedimento es eólico arenoso de color pardo, muy suelto y con gran cantidad de rodados pequeños. Las valvas de moluscos se disponen en forma azarosa sin estructura. En estratigrafía se recuperaron restos líticos, cerámicos, faunísticos marinos y terrestres, y antracológicos (carbones dispersos). Algunos rasgos negativos sugieren posibles huellas de poste. La fauna marina comprende pinnípedos, peces, gasterópodos y bivalvos, mientras que la fauna terrestre está constituida por guanacos y restos de huevos de ñandú. Algunos de estos últimos presentan rasgos de exposición al fuego. En el conjunto lítico predominan los desechos mientras que las formas base potenciales aparecen en muy baja proporción. Prevalece la calcedonia translúcida de disponibilidad local (Zubimendi et al., 2010). El estudio de este registro sugiere actividades de consumo de recursos marinos y terrestres en igual proporción, últimas etapas de talla de los artefactos líticos, y una ocupación del sitio durante la primavera verano (Zubimendi et al., 2010).

Fig. 3 A, bosque caducifolio en los alrededores de CSP2017. B, vista de excavación sitio CSP2017. C, D, detalle de los materiales capa B. (B-D tomado de Franch et al., 2020). Figura en color en la versión en línea http://www.ojs.darwin. edu.ar/index.php/darwiniana/article/view/1020/1254
Provincia de Tierra del Fuego
El sitio CSP2017 se encuentra a 1 km hacia el oeste del cabo San Pablo, en la Isla Grande de Tierra del Fuego (Fig. 1, Fig. 3A), y está fechado en 630 ± 40 años AP (LP-3536). En el sitio CSP2017, se excavaron 9 sectores de 1x1 m2 de un total de 4 cuadrículas (2x2 m2) (Fig. 3B). Presenta una estratigrafía dividida en 3 capas. La capa A, que corresponde al nivel superficial, es orgánica ya que presenta restos vegetales como raíces. La capa B es un sedimento oscuro de humus en la que se encuentran los restos arqueológicos; el material se presenta inicialmente con mayor densidad y luego se torna escaso a medida que se profundiza la excavación. Finalmente, la capa C, está compuesta por una arcilla grisácea totalmente estéril (para más detalle ver Franch et al., 2020) (Fig. 3C, D). Dicha capa presenta una gran diversidad de restos arqueológicos, entre los que se incluyen restos faunísticos de vertebrados e invertebrados, restos de material lítico y restos carpológicos y antracológicos. La fauna terrestre está representada por restos de aves, roedores y mamíferos; la fauna marina incluye pinnípedos y peces, y los invertebrados son en su totalidad moluscos, tales como gasterópodos y bivalvos. El conjunto lítico está compuesto por artefactos bifaciales y unifaciales, elaborados con materia prima local, principalmente riolita, lutita y cinerita, aunque también se halló un fragmento de madera petrificada (De Angelis et al., 2017; Franch et al., 2021).
Materiales y métodos
Colección de referencia
Se realizó la colecta de ejemplares para la confección de un herbario que permita la correcta identificación a nivel de género y/o especie de las plantas leñosas más representativas del área de estudio, dentro de la que se incluyó una muestra de madera de cada una. Se colectaron dieciocho taxones de leños de porte arbustivo del área litoral de la provincia de Santa Cruz (Adesmia boronioides Hook. f., Atriplex lampa, Baccharis darwinii Hook. & Arn., Berberis microphylla., Erythrostemon gilliesi var. gilliesii,Chuquiraga avellanedae, Fabiana imbricata, Lepidophyllum cupressiforme, Nassauvia sp., Schinus sp., Senecio sp., Colliguaja integerrima, Lycium sp., Mulguraea tridens, Nardophyllum bryoides, Prosopis denudans, Prosopidastrum globosum, Suaeda divaricata). C. gillesii var. gilliesii y S. divaricata no pudieron colectarse en el área de estudio, por lo que se utilizan especímenes colectados por Capparelli en la provincia fitogeográfica del Monte. Asimismo, se colectaron doce taxones de plantas de porte arbustivo y arbóreo, del área litoral de la Isla Grande de Tierra del Fuego (Berberis microphylla, Berberis empetrifolia Lam., Berberis ilicifolia, Chiliotrichum diffusum, Drimys winteri, Embothrium coccineum, Maytenus magellanica, Nothofagus antarctica, Nothofagus betuloides, Nothofagus pumilio, Gaultheria mucronata y Ribes magellanicum Poir.). Las muestras de madera se obtuvieron de ramas secundarias; de cada una se cortaron 6 cubos de 1,5 cm de lado, uno de los cuales se seleccionó para la realización de cortes histológicos, uno para el cálculo de la densidad y los otros cuatro tacos para la realización de una carbonización experimental (Capparelli et al., 2003; Andreoni, 2009, 2010, 2014; Ciampagna, 2015, 2016).
El cubo destinado a los cortes histológicos de cada ejemplar se sometió a hervor en una mezcla de agua con detergente industrial entre 2 a 8 horas dependiendo de la dureza de la madera. Posteriormente, se realizaron los cortes con micrótomo de deslizamiento en los tres planos de observación (transversal, longitudinal tangencial y longitudinal radial), de un grosor de 28µm a 30µm. Se decoloraron los cortes con hipoclorito de sodio al 50%, entre 5 y 10 minutos, para eliminar los contenidos celulares. Se lavaron los cortes en agua destilada durante 10 minutos para eliminar los restos de hipoclorito de sodio. A continuación, se deshidrataron y tiñeron los cortes con safranina en un batería de alcoholes durante 5 minutos (alcohol 50º con safranina al 1%, alcohol 70°, alcohol 80°, alcohol 96°, alcohol 100°, alcohol 100º y xilol en partes iguales y xilol 100%). Finalmente, se montaron los cortes de forma permanente con Bálsamo de Canadá (Ancibor & Pérez de Micou, 1995; Andreoni, 2009, 2014; Ciampagna, 2015). Estos preparados se observaron a través de un microscopio óptico de luz transmitida, se identificaron y describieron los caracteres cualitativos diagnósticos de la madera siguiendo los criterios de la IAWA List of Microscope Features for Hardware Identification (Wheeler et al., 1989; Richter et al., 2004; Capparelli et al., 2009) pero con énfasis en caracteres cualitativos más que cuantitativos, ya que se busca facilitar el análisis antracológico y estos últimos caracteres suelen alterarse con la carbonización (Rossen & Olson, 1985). En el caso de Baccharis darwinii que no se cuenta con material fresco, pero sí con material de referencia carbonizado, se reporta foto de su anatomía a través de su observación bajo luz incidente.
Los ejemplares de herbario y los cortes histológicos se encuentran depositados en la colección experimental del Laboratorio 129, Edificio Anexo I, Unidades de Investigación del Museo de Ciencias Naturales de La Plata y en el Laboratorio del GIATMA del CADIC-CONICET, Ushuaia, Argentina.
A partir de la descripción anatómica se confeccionó una clave dicotómica que facilite la identificación de material antracológico. Para la confección de dicha clave, además del material actual examinado en el presente trabajo, se consideraron taxones presentes en el área de estudio que no pudieron ser colectados en el campo, colectados previamente y descritos por Andreoni (2010, 2015a, b) y Andreoni y Capparelli (2012) para Cuyo: Azorella (= Mulinum) sp., Ephedra sp., Condalia sp., Grindelia sp., Prosopis alpataco Phil. f., Larrea sp., Ochetophylla sp.
Material actual examinado
Adesmia boronioides
ARGENTINA. Santa Cruz. Puerto Deseado, Bahía del Oso Marino, 23-I-2011, Ciampagna 654, colección de herbario Laboratorio 129, FCNyM.
Atriplex lampa
ARGENTINA. Santa Cruz. Puerto Deseado, Punta Buque, 1-II-2011, Ciampagna 664, colección de herbario Laboratorio 129, FCNyM.
Baccharis sp
ARGENTINA. Santa Cruz. Puerto Deseado, Cañadón Torcido,6-II-2012, Ciampagna 706, colección de herbario Laboratorio 129, FCNyM.
Berberis empetrifolia
ARGENTINA. Tierra del Fuego. Cabo San Pablo, 21-XI-2017, Franch 220, colección de herbario Laboratorio 129, FCNyM,
Berberis ilicifolia
ARGENTINA. Tierra del Fuego. Ushuaia, Playa Larga, 23-X-2017, Franch 116, colección de herbario Laboratorio 129, FCNyM.
Berberis microphylla
ARGENTINA. Santa Cruz. Caleta Olivia, Cañadón León, 11-X-2009, Ciampagna 604, colección de herbario Laboratorio 129, FCNyM. Tierra del Fuego. Ushuaia, Playa Larga, 23-X-2017, Franch 101, colección de herbario Laboratorio 129, FCNyM.
Chiliotrichum diffusum
ARGENTINA. Tierra del Fuego. Ushuaia, Playa Larga, 23-X-2017, Franch 109, colección de herbario Laboratorio 129, FCNyM.
Chuquiraga avellanedae
ARGENTINA. Santa Cruz. Caleta Olivia, Cañadón León, 11-X-2009, Ciampagna 601, colección de herbario Laboratorio 129, FCNyM.
Colliguaja integerrima
ARGENTINA. Santa Cruz. Caleta Olivia, Cañadón León, 11-X-2009, Ciampagna 602, colección de herbario Laboratorio 129, FCNyM.
Drimys winteri
ARGENTINA. Tierra del Fuego. Ushuaia, Playa Larga, 23-X-2017, Franch 114, colección de herbario Laboratorio 129, FCNyM.
Embothrium coccineum
ARGENTINA. Tierra del Fuego. Playa Larga, 2017, Franch 129, colección de herbario Laboratorio 129, FCNyM.
Erythrostemon gilliesi var. gilliesii
ARGENTINA. Catamarca. El Shincal, 7-IV-1992, Capparelli 60 (LP), Capparelli 65, colección de herbario Laboratorio 129, FCNyM.
Fabiana imbricata
ARGENTINA. Chubut. Trevelin. 10-II-2011, Ciampagna 674, colección de herbario Laboratorio 129, FCNyM.
Gaultheria mucronata
ARGENTINA. Tierra del Fuego. Ushuaia, Playa Larga, 23-X-2017, Franch 128, colección de herbario Laboratorio 129, FCNyM.
Lepidophyllum cupressiforme
ARGENTINA. Santa Cruz. Puerto Deseado, Cañadón del Puerto, 8-II-2012, Ciampagna 725, colección de herbario Laboratorio 129, FCNyM.
Lycium sp
ARGENTINA. Santa Cruz. Puerto Deseado, Peñón Azopardo, 19-I-2011, Ciampagna 637, colección de herbario Laboratorio 129, FCNyM.
Maytenus magellanica
ARGENTINA. Tierra del Fuego. Ushuaia, Playa Larga, 23-X-2017, Franch 125, colección de herbario Laboratorio 129, FCNyM.
Mulguraea tridens
ARGENTINA. Santa Cruz. Puerto Deseado, Cañadón Torcido, 17-XI-2012, Ciampagna 748, colección de herbario Laboratorio 129, FCNyM.
Nardophyllum bryoides
ARGENTINA. Santa Cruz. Puerto Deseado, Bahía del Oso Marino, 20-I-2011, Ciampagna 641, colección de herbario Laboratorio 129, FCNyM.
Nassauvia sp
ARGENTINA. Santa Cruz. Puerto Deseado, 17-XII-2012, Ciampagna 755, colección de herbario Laboratorio 129, FCNyM.
Nothofagus antárctica
ARGENTINA. Tierra del Fuego. Cabo San Pablo, 21-XI-2017, Franch 164, colección de herbario Laboratorio 129, FCNyM.
Nothofagus betuloides
ARGENTINA. Tierra del Fuego. Ushuaia, Playa Larga, 23-X-2017, Franch 113, colección de herbario Laboratorio 129, FCNyM.
Nothofagus pumilio
ARGENTINA. Tierra del Fuego. Laguna Bombilla, 21-XI-2017, Franch 192, colección de herbario Laboratorio 129, FCNyM.
Prosopis denudans
ARGENTINA, Santa Cruz. Caleta Olivia, Cañadón León, 11-X-2009, Ciampagna 610, colección de herbario Laboratorio 129, FCNyM.
Prosopidastrum globosum
ARGENTINA. Santa Cruz. Caleta Olivia, Cañadón León, 10-IX-2009, Ciampagna 589, colección de herbario Laboratorio 129, FCNyM.
Ribes magellanicum
ARGENTINA. Tierra del Fuego. Ushuaia, Playa Larga, 23-X-2017, Franch 115, colección de herbario Laboratorio 129, FCNyM.
Schinus sp
ARGENTINA. Santa Cruz. Caleta Olivia, Cañadón León, 11-X-2009, Ciampagna 609, colección de herbario Laboratorio 129, FCNyM.
Recuperación e identificación del material antracológico
Sitio Palo Alto
El sondeo inicial (Sondeo 1) fue de 5 x 5 cm hasta los 27 cm de profundidad máxima en un sector de media a alta densidad de restos materiales malacológicos en superficie. Se excavaron nueve niveles artificiales de 5 cm de potencia a excepción de identificar cambios sedimentológicos significativos. Luego se realizaron cuatro cuadrículas más, de 1 m2 cada una, al sudeste del primer sondeo. Los carbones dispersos estudiados provienen de los niveles uno a cinco del estrato fértil del sondeo 1, mientras que del nivel 6 hasta el nivel 9 no se hallaron carbones. El registro antracológico de las cuadrículas uno a cuatro se encuentra todavía en estudio. Estos carbones fueron obtenidos por medio de una zaranda en seco con malla de 2 mm y de los fondos de zaranda (Zubimendi et al., 2010). El total de sedimento tamizado fue de 78,75 l, que corresponde a 11,25 l por nivel. El denominado fondo de zaranda se analizó en el laboratorio donde se registró la presencia/ausencia de otros restos arqueobotánicos (Ciampagna, 2015).
Sitio CSP2017
Para el sitio CSP2017 la estrategia empleada fue la recogida sistemática de 4l de sedimento de cada uno de los 9 m2 excavados y por cada nivel arqueológico. Esta recogida sistemática se combinó con una recogida más intensiva de sedimentos, la totalidad o cantidad representativa de contextos puntuales como estructuras de combustión, áreas de descarte, entre otras. Estas muestras de sedimento se sometieron a flotación en el laboratorio de antropología del CADIC (Ushuaia, Tierra del Fuego, Argentina). El resto de sedimento extraído del sitio arqueológico fue cribado o tamizado en agua durante el transcurso de la excavación, mediante cribas con luz de malla de entre 5 y 2 mm, permitiendo una rápida limpieza y selección del material arqueobotánico en el que se incluyen los restos leñosos carbonizados (para más detalle ver Mansur & De Angelis, 2013; Franch et al., 2020).
Identificación taxonómica de los carbones arqueológicos
La identificación de las especies vegetales que forman el registro antracológico, se realizó mediante la observación de cada fragmento a través de un microscopio óptico equipado con luz incidente y polarización. En ambos sitios el tamaño de la submuestra fue delimitado a través de la realización de una curva de riqueza específica (Chabal, 1989; Perez de Micou, 1991; Badal, 1992; Marconetto, 2005). Los carbones se fracturaron manualmente y se describieron los caracteres anatómicos diagnósticos según los planos de observación (corte transversal, longitudinales tangencial y radial), así como presencia o ausencia de hifas y de alteraciones ocurridas por la carbonización. Se compararon con el material de referencia y se pasaron por la clave dicotómica realizada a partir del mismo. Se utilizó también material bibliográfico específico para el área y otros textos de anatomía de la madera acotados a determinados taxones (Wheeler et al., 1989; Solari, 1992; Ancibor & Pérez de Micou, 1995, 2002; Garibotti, 1998; Piqué, 1999; Marconetto, 2002; Richter et al., 2004; Roig & Videla, 2006-2010; Capparelli et al., 2009; Ortega & Marconetto, 2009; Tortorelli, 2009; Andreoni, 2010; Andreoni & Capparelli, 2012; Caruso, 2012; Guerra et al., 2012; Rodríguez, 2013; Andreoni, 2014, 2015a, b; Ciampagna, 2015, 2016; Cueto & Andreoni, 2016; Ciampagna et al., 2016). Se calculó la frecuencia relativa de cada taxón por nivel, la ubicuidad en términos del porcentaje de muestras en las cuales un taxón estuvo presente y los porcentajes de alteraciones de los carbones por nivel; para más detalle ver Ciampagna (2015) y Franch et al. (2020).
Resultados
Colección de referencia: Descripción de los caracteres diagnósticos de la estepa arbustiva del Golfo San Jorge (Figs. 4 a 7).
Familia Amarantaceae
Atriplex lampa (Material bibliográfico de referencia consultado: Marconetto, 2002; Roig & Videla, 2006-2010; Ortega & Marconetto, 2009; Andreoni, 2014, 2015a; Cueto & Andreoni, 2016).
Plano transversal: Anillos de crecimiento demarcado por hileras de parénquima en bandas de dos a tres células. Porosidad semicircular. Cámbium anómalo, floema incluso tipo difuso, dentro del parénquima paratraqueal y en cada conjunto de vasos del leño tardío. Vasos en patrón diagonal y radial. Vasos exclusivamente solitarios. Fibras de pared muy gruesa. Madera sin radios. (Fig. 4A).
Plano longitudinal tangencial: Placas de perforación simples y oblicuas. Punteaduras intervasculares alternas con rebordes conspicuos (areoladas). Engrosamientos helicoidales a lo largo de los elementos de vaso de espiras anchas. Células parenquimáticas fusiformes con dos células por serie parenquimática. Parénquima axial y/o elementos de vaso parcialmente estratificados (Fig. 4B).
Plano longitudinal radial: no se realizó, dado que la madera carece de radios.
Suaeda divaricata (Material bibliográfico de referencia consultado: Capparelli 2004).
Plano transversal: Anillos de crecimiento no demarcados o ausentes. Cámbium anómalo, floema incluso tipo difuso concéntrico, dentro de bandas de parénquima de más de cuatro células de ancho. Madera sin radios. Vasos solitarios, y en menor proporción múltiples cortos de dos a tres. Fibras de paredes delgadas a gruesas (Fig. 4C).
Plano longitudinal tangencial: Placas de perforación simple. Engrosamientos espiralados en vasos angostos y fibras. Células parenquimáticas fusiformes con dos células por serie parenquimática. Grupos de células correspondientes al floema incluso. Estratificación parcial de vasos, parénquima axial y fibras (Fig. 4D).
Plano longitudinal radial: no se realizó, dado que la madera carece de radios.
Familia Asteraceae
Baccharis darwinii (Material bibliográfico de referencia consultado: Garibotti, 1998; Rodríguez 2008; Andreoni, 2010; Guerra et al., 2012).
Plano transversal: Anillos de crecimiento demarcados por parénquima marginal y vasos en banda tangencial. Porosidad semicircular a difusa, con vasos en disposición radial, diagonal y dendrítica. Vasos múltiples radiales cortos, agrupados y solitarios. Parénquima axial paratraqueal confluente. Parénquima axial apotraqueal difuso en agregados. Fibras de paredes delgadas a gruesas (Fig. 4E).
Plano longitudinal tangencial: Placas de perforación simple, oblicuas. Punteaduras intervasculares alternas. Engrosamientos helicoidales en vasos. Radios de uni a triseriados así como de cuatro a diez seriados. Algunos radios agregados. Tipo de radios homogéneos de células elongadas. Estratificación parcial (solo en fibras) (Fig. 4F).
Plano longitudinal radial: Radios de células erectas y cuadradas mezcladas (Fig. 4G).
Chuquiraga avellanedae (Material bibliográfico de referencia consultado: Ortega & Marconetto, 2009; Andreoni, 2010; Andreoni, 2014).
Plano transversal: Anillos de crecimiento demarcados por la porosidad semicircular, el achatamiento de las fibras y una hilera de vasos dispuesta tangencialmente en el leño temprano. Vasos exclusivamente solitarios. Fibras de paredes delgadas a gruesas. Parénquima axial ausente o extremadamente escaso (Fig. 4H).
Plano longitudinal tangencial: Placas de perforación simple, oblicuas. Punteaduras intervasculares alternas poligonales. Engrosamiento helicoidal tenue en los elementos de vaso, siendo los elementos de vasos muy finos. Fibras con punteaduras con rebordes conspicuos (fibrotraqueidas). Engrosamiento helicoidal en fibras del tejido básico. Radios uni a tri-seriados y grandes de cuatro a diez-seriados (prevalecen los de siete células de ancho). Son radios homogéneos de células elongadas. Células radiales perforadas (Fig. 4I).
Plano longitudinal radial: Placas de perforación simple, oblicuas. Punteaduras vaso-radiales con rebordes visibles, similares a punteaduras intervasculares en tamaño y forma. Radios con células procumbentes, cuadradas y erectas mezcladas (Fig. 4J).
Lepidophyllum cupressiforme (Material bibliográfico de referencia consultado: Caruso, 2012).
Plano transversal: Anillos de crecimiento demarcados por fibras achatadas en el leño tardío, por porosidad semicircular y por una hilera de vasos en banda tangencial en el leño temprano. Algunos vasos en patrón diagonal en el leño tardío. Vasos exclusivamente solitarios. Fibras de pared delgada a gruesa. Parénquima axial apotraqueal difuso y difuso en agregados (Fig. 4K).

Fig. 4 Lámina de cortes histológicos de la estepa arbustiva del Golfo San Jorge; corte transversal (izq), longitudinal tangencial (centro) y radial (der): A-B, Atriplex lampa. C-D, Suaeda divaricata. E-G, Baccharis darwinii. H-J, Chuquiraga avellanedae. K-M, Lepidophyllum cupressiforme. Figura en color en la versión en línea http://www.ojs. darwin.edu.ar/index.php/darwiniana/article/view/1020/1254
Plano longitudinal tangencial: Placas de perforación simple, oblicuas. Punteaduras intervasculares alternas. Engrosamientos helicoidales presentes en los elementos de vaso. Fibrotraqueidas vasculares vasicéntricas presentes. Fibras con punteaduras simples a punteaduras con rebordes diminutos y fibras con reborde conspicuo (fibrotraqueidas). Engrosamientos helicoidales en fibras del tejido básico. Radios uni a triseriados (generalmente bi y triseriados) y cuatro a diez-seriados (generalmente cuatro a ocho), homogéneos de células alargadas. Se observó una baja densidad de radios (10 radios/mm). Células del radio perforadas (Fig. 4L).
Plano longitudinal radial: Todas las células del radio erectas y/o cuadradas. Punteaduras vaso radiales con rebordes visibles, similares a punteaduras intervasculares en tamaño y forma a lo largo de las células radiales (Fig. 4M).
Nardophyllum bryoides
Plano transversal: Anillos de crecimiento demarcados por fibras achatadas en el leño temprano. Porosidad difusa. Vasos en patrón diagonal y radial. Vasos solitarios y en menor medida múltiples cortos de dos, todos de contorno angular. Fibras de paredes delgadas a gruesas. Parénquima axial ausente o extremadamente escaso (Fig. 5A).
Plano longitudinal tangencial: Predominantemente placas de perforación simples, aunque también escalariformes de doce barras. Elementos de vaso con apéndice en los extremos. Punteaduras intervasculares alternas. Fibras con punteaduras con reborde conspicuo (fibrotraqueidas). Radios uni a tri-seriados, con células alargadas, homogéneos (Fig. 5B).
Plano longitudinal radial: Todas las células del radio erectas y/o cuadradas (Fig. 5C).
Nassauvia sp.
Plano transversal: Anillos de crecimiento demarcados por porosidad semicircular y por una hilera de vasos en disposición tangencial en el leño temprano. Vasos en disposición radial en el leño tardío. Vasos exclusivamente solitarios. Fibras de paredes delgadas a gruesas en el leño tardío (fibrotraqueidas) y muy gruesas en el leño temprano (Fig. 5D).
Plano longitudinal tangencial: Placas de perforación simple, oblicuas. Vasos con y sin apéndices cortos. Engrosamientos helicoidales en los elementos de vaso. Traqueidas vasculares vasicéntricas presentes, fibras con punteaduras con reborde conspicuo (fibrotraqueidas). No se observaron radios (Fig. 5E).
Plano longitudinal radial: no se realizó, dado que la madera carece de radios.
Senecio sp. (Material bibliográfico de referencia consultado: Roig & Videla, 2006-2010; Andreoni, 2014)
Plano transversal: Anillos de crecimiento demarcados por hileras de fibras achatadas. Porosidad difusa. Vasos en patrón diagonal y radial, solitarios y radiales múltiples de dos, tres y cuatro. Vasos de contorno angular. Fibras de pared muy delgada. Parénquima axial apotraqueal difuso en agregados (Fig. 5F).
Plano longitudinal tangencial: Placas de perforación simple, oblicuas. Vasos con apéndices. Punteaduras intervasculares alternas y opuestas. Traqueidas vasculares vasicéntricas presentes. Fibras con punteaduras con reborde conspicuo (fibrotraqueidas). Radios uni a tri-seriados y grandes hasta diez-seriados, homogéneos de células elongadas (Fig. 5G).
Plano longitudinal radial: Punteaduras vaso-radiales con rebordes visibles, similares a punteaduras intervasculares en tamaño y forma. Radios de células procumbentes, erectas y cuadradas mezcladas. Radios de células perforadas (Fig. 5H).
Familia Euphorbiaceae
Colliguaja integerrima (Material bibliográfico de referencia consultado: Ancibor & Pérez de Micou, 1995; Marconetto, 2002; Andreoni & Capparelli, 2012; Cueto & Andreoni, 2016).
Plano transversal: Anillos de crecimiento demarcados por hileras de fibras achatadas y por porosidad semicircular. Vasos en bandas tangenciales en el leño temprano, en patrón diagonal y radial en leño tardío. Vasos solitarios, y radiales múltiples de cuatro o más. Vasos de contorno angular. Fibras de pared muy delgada. Parénquima axial ausente o extremadamente escaso (Fig. 5I).
Plano longitudinal tangencial: Placas de perforación simple, oblicuas. Punteaduras intervasculares alternas.
Engrosamientos helicoidales en los elementos de vaso. Fibras con punteaduras con rebordes conspicuos (fibrotraqueidas). Engrosamientos helicoidales en fibras del tejido básico. Radios mayormente uniseriados, escasamente bi y parcialmente triseriados, heterogéneos III (según Tortorelli 2009). Células radiales perforadas (Fig. 5J).
Plano longitudinal radial: Punteaduras vaso-radiales con reborde visibles, similares a punteaduras intervasculares en tamaño y forma a lo largo de las células radiales. Todas las células del radio erectas y/o cuadradas (Fig. 5K).

Fig. 5 Lámina de cortes histológicos de la estepa arbustiva del Golfo San Jorge (cont.); corte transversal (izq), longitudinal tangencial (centro) y radial (der): A-C, Nardophyllum bryoides. D-E, Nassauvia sp. F-H, Senecio sp. I-K, Colliguaja integerrima. L-N, Adesmia boronioides. Figura en color en la versión en línea http://www.ojs.darwin. edu.ar/index.php/darwiniana/article/view/1020/1254
Familia Fabaceae
Adesmia boronioides (Material bibliográfico de referencia consultado: Garibotti, 1998; Roig & Videla, 2006-2010; Andreoni, 2010; Andreoni & Capparelli, 2012).
Plano transversal: Anillos de crecimiento demarcados por vasos en bandas tangenciales en leño temprano, porosidad semicircular y vasos en patrón diagonal a dendrítico en leño tardío. Predominan vasos solitarios angulares. Fibras de pared muy gruesa. Parénquima axial apotraqueal difuso. Parénquima axial paratraqueal confluente acompañando en el leño tardío a los vasos en disposición dendrítica y diagonal. Parénquima axial en bandas marginales de cinco células de espesor en el leño temprano asociado a los vasos (Fig. 5L).
Plano longitudinal tangencial: Placas de perforación simples, horizontales y oblicuas. Punteaduras intervasculares alternas y opuestas. Engrosamientos helicoidales a lo largo de los vasos dispuestos en espiras apretadas y horizontales. Fibras con punteaduras simples a fibras con punteaduras de rebordes diminutos. Fibras no septadas presentes. Células parenquimáticas fusiformes, una célula por serie parenquimática. Radios uni a tri-seriados y radios grandes de cuatro a diez-seriados (comúnmente siete seriados) de tipo Heterogéneos II de acuerdo con Tortorelli (2009). Células envolventes en los radios. Estratificación parcial en parénquima (Fig. 5M).
Plano longitudinal radial: Punteaduras vaso-radiales con rebordes visibles, similares a punteaduras intervasculares en tamaño y forma a lo largo de las células radiales. Radios con células procumbentes, cuadradas y erectas mezcladas (Fig. 5N).
Prosopis denudans (Material bibliográfico de referencia consultado: Castro 1994).
Plano transversal: Anillos de crecimiento demarcados por porosidad semicircular. Vasos solitarios y múltiples cortos de dos, tres y cuatro, con contenido, vasos agrupados en el leño tardío. Fibras de pared muy gruesa. Parénquima axial paratraqueal confluente y en bandas (Fig. 6A).
Plano longitudinal tangencial: Placas de perforación simple, oblicuas y horizontales. Vasos con apéndices cortos y sin apéndices. Punteaduras intervasculares alternas, ornadas. Fibras con punteaduras simples con rebordes diminutos. Células parenquimáticas fusiformes de dos células por serie parenquimática. Radios uni a tri-seriados (en gran proporción uni-seriados y parcialmente bi-seriados y en menor medida tri-seriados). Radios de tipo heterogéneo homocelular. Cristales prismáticos en fibras y radios (Fig. 6B).
Plano longitudinal radial: Punteaduras vaso-radiales con rebordes muy reducidos hasta con apariencia de simples. Todas las células del radio procumbentes con una hilera de células erectas y/o cuadradas (Fig. 6C).
Prosopidastrum globosum (Material bibliográfico de referencia consultado: Roig & Videla, 2006-2010; Andreoni, 2014).
Plano transversal. Anillos de crecimiento demarcados por hileras células de parénquima marginal de disposición achatada, por porosidad semicircular y por vasos en disposición tangencial en leño temprano. Vasos en disposición diagonal a dendrítica en leño tardío. Vasos solitarios y vasos múltiples radiales cortos. Vasos agrupados frecuentes. Fibras de pared muy gruesa. Parénquima axial en bandas angostas terminales, y paratraqueal confluente y en bandas mayor de tres células de ancho y angostas de hasta tres células de ancho (Fig. 6D).
Plano longitudinal tangencial: Placas de perforación simple, oblicuas. Punteaduras intervasculares alternas. Engrosamientos helicoidales y reticulados presente a lo largo del elemento de vaso. Fibras septadas presentes. Células parenquimáticas fusiformes, dos a tres células por serie parenquimática. Radios uni a triseriados de tipo homogéneos II de acuerdo con Tortorelli (2009) (Fig. 6E).
Plano longitudinal radial: Todas las células radiales procumbentes. Cristales prismáticos presentes en fibras. Andreoni (2014) observa células procumbentes y cuadradas mezcladas (Fig. 6F).
Familia Caesalpinoideae
Erythrostemon gilliesi var. gilliesii (Material bibliográfico de referencia consultado Capparelli 2004; Andreoni, 2015a)
Plano transversal: Anillos de crecimiento demarcados por pocas hileras de células de parénquima axial marginal y por porosidad semicircular a difusa. Vasos solitarios y múltiples radiales de dos a cuatro. Vasos en bandas tangenciales en el leño temprano. Fibras de pared muy gruesa. Parénquima axial paratraqueal vasicéntrico, confluente y en bandas de hasta tres células y de más de tres células de ancho. Escaso parénquima axial apotraqueal difuso (Fig. 6G).
Plano longitudinal tangencial: Placas de perforación simple, horizontales a oblicuas. Punteaduras alternas, ornadas. Células parenquimáticas fusiformes y de 2 a 3 células por serie parenquimática. Radios unia tri-seriados y radios grandes 4-5 (parcialmente seis) seriados. Estos últimos pueden tener más de 1 mm de alto. Radios agregados presentes. Células envolventes en radios. Engrosamientos helicoidales en vasos angostos del leño tardío (Fig. 6H).
Plano longitudinal radial. Células del cuerpo del radio procumbentes y una hilera marginal de células erectas o cuadradas. Cristales prismáticos en cadenas cortas (seis a dieciocho) en células axiales septadas (Fig. 6I).

Fig. 6 Lámina de cortes histológicos de la estepa arbustiva del Golfo San Jorge (cont.); corte transversal (izq), longitudinal tangencial (centro) y radial (der): A-C, Prosopis denudans. D-F, Prosopidastrum globosum. G-I, Erythrostemon gilliesii var. gilliesii. J-L, Mulguraea tridens. M-Ñ, Berberis microphylla. Figura en color en la versión en línea http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/1020/1254
Familia Verbenaceae
Mulguraea tridens (Material bibliográfico de referencia consultado: Andreoni & Capparelli, 2012; Caruso, 2012).
Plano transversal: Anillos de crecimiento demarcados por porosidad semicircular. Disposición de los vasos en bandas tangenciales en leño temprano y en patrón diagonal y dendrítico en leño tardío. Vasos exclusivamente solitarios y de contorno angular. Fibras de paredes delgadas a gruesas (Fig. 6J).
Plano longitudinal tangencial: Placas de perforación simple, oblicuas. Punteaduras intervasculares alternas, poligonales. Engrosamientos helicoidales a lo largo de los elementos de vaso. Fibras con punteaduras con rebordes conspicuos (fibrotraqueidas). Engrosamientos espiralados en fibras del tejido básico (firbrotraqueidas). Fibras no septadas presentes. Radios uni a triseriados (principalmente uni y bi-seriados) de tipo heterogéneo Iib (según Tortorelli, 2009) (Fig. 6K).
Plano longitudinal radial: Punteaduras vaso-radiales con rebordes visibles, similares a punteaduras intervasculares en tamaño y forma a lo largo de las células radiales. Radios con células procumbentes, cuadradas y erectas mezcladas (Fig. 6L).
Familia Berberidaceae
Berberis microphylla (Santa Cruz; Material bibliográfico consultado: Ancibor & Pérez de Micou, 1995; Piqué, 1999; Capparelli et al. 2009; Andreoni, 2010; Caruso, 2012)
Plano transversal: Anillos de crecimiento demarcados, porosidad semicircular. Vasos en bandas tangenciales con disposición ulmoide (o festoneada) en el leño temprano y diagonal y/o dendrítica en el leño tardío. Vasos exclusivamente solitarios, de contorno angular, algunos con contenido. Fibras de pared muy gruesa. Parénquima axial ausente o extremadamente escaso (Fig. 6M).
Plano longitudinal tangencial: Placas de perforación simple, oblicuas. Punteaduras intervasculares, principalmente alternas poligonales, raramente opuestas. Engrosamiento helicoidal a lo largo de los elementos de vasos. Fibras con punteaduras con rebordes conspicuos (fibrotraqueidas). Fibras con punteaduras en las paredes radiales y tangenciales. Fibras del tejido básico con espesamiento espiralado. Radios de dos tamaños distintos. Radios uni a tri-seriados (comúnmente bi-seriados) y radios grandes cuatro a diez-seriados (comúnmente ocho seriados), homogéneos. Radios de altura mayor a 1 mm. Células envolventes en radios. Cristales prismáticos presentes (Fig. 6N).
Plano longitudinal radial: Punteaduras vaso-radiales con rebordes visibles, similares a las punteaduras intervasculares en tamaño y forma a lo largo de la célula radial. Radios con células cuadradas, erectas y procumbentes mezcladas. Cristales prismáticos en células radiales procumbentes (Fig. 6Ñ).
Familia Anacardiaceae
Schinus sp. (Material bibliográfico de referencia consultado: Ancibor & Pérez de Micou, 1995; Garibotti, 1998; Ancibor & Pérez de Micou, 2002; Marconetto, 2002; Capparelli, 2004; Roig & Videla, 2006 -2010; Ortega & Marconetto, 2009; Andreoni, 2010; Andreoni & Capparelli, 2012; Andreoni, 2014).
Plano transversal: Anillos de crecimiento demarcados por la porosidad semicircular y una hilera de fibras achatadas. Vasos en bandas tangenciales en el leño temprano, en patrón diagonal y radial (múltiples de 4, 5 y 6) en el leño tardío. Vasos solitarios y agrupados frecuentes en el leño tardío. Fibras de pared delgada a gruesa y parénquima axial ausente o extremadamente escaso (Fig. 7A).
Plano longitudinal tangencial: Placas de perforación simple, oblicuas. Punteaduras intervasculares alternas poligonales. Engrosamiento reticulado a lo largo del elemento de vaso. Fibras con punteaduras simples a punteadoras con rebordes diminutos. Engrosamientos espiralados en fibras del tejido básico, fibras septadas presentes. Radios uni a tri-seriados heterogéneos tipo I con extremos uniseriados de hasta cuatro células (Tortorelli, 2009, Guerra et al., 2012). Presencia de canales radiales. Cristales prismáticos presentes (Fig. 7B).
Plano longitudinal radial: Punteaduras vaso-radiales con rebordes visibles, similar a punteaduras intervasculares en tamaño y forma a lo largo de la célula radial. Células del radio procumbentes con una hilera de células erectas y/o cuadradas en los márgenes, también se registraron radios con células cuadradas, erectas y procumbentes mezcladas. Se observaron cristales prismáticos en células radiales, cuadradas y erectas (Fig. 7C).
Familia Solanaceae
Fabiana imbricata (Material bibliográfico de referencia consultado: Rodríguez, 2004 -Fabiana bryoides Phil.-, 2008).
Plano transversal: Anillos de crecimiento demarcados por hileras de fibras achatadas. Porosidad semicircular a difusa. Vasos en una banda tangencial en leño temprano y patrón diagonal y/o radial en el leño tardío. Vasos mayormente solitarios, escasamente múltiples radiales cortos (dos y tres) y agrupados. Fibras de pared delgada a gruesa. Parénquima axial apotraqueal difuso (Fig. 7D).
Plano longitudinal tangencial: Placas de perforación simple, oblicuas. Punteaduras intervasculares alternas. Fibras con punteaduras con rebordes conspicuos (fibrotraqueidas). Radios exclusivamente uniseriados homogéneos; Rodríguez (2004) observa radios heterogéneos, heterocelulares. Células parenquimáticas fusiformes y de 3 a 4 células por serie (Fig. 7E).
Plano longitudinal radial: Células del cuerpo del radio procumbentes con una hilera de células marginales erectas y/o cuadradas. Radios con células procumbentes, cuadradas y erectas mezcladas. Cristales prismáticos en células radiales cuadradas y erectas (Fig. 7F).
Lycium sp. (Material bibliográfico de referencia consultado: Capparelli 2004; Roig & Videla, 2006-2010; Ortega & Marconetto, 2009; Andreoni & Capparelli, 2012).
Plano transversal: Anillos de crecimiento demarcados por porosidad semicircular. Vasos en bandas tangenciales en leño temprano, y patrón diagonal a dendrítico en el tardío. Vasos exclusivamente solitarios. Fibras de pared delgada a gruesa. Parénquima axial apotraqueal difuso en agregados y paratraqueal escaso. Parénquima axial marginal o aparentemente en bandas marginales de una hilera de células (Fig. 7G).
Plano longitudinal tangencial: Placas de perforación simple, oblicuas. Punteaduras intervasculares alternas. Engrosamiento helicoidal presente en elementos de vaso. Fibras con punteaduras simples a punteaduras con rebordes diminutos. Fibras con punteaduras con rebordes conspicuos (fibrotraqueidas). Engrosamiento helicoidal en fibras del tejido básico. Fibras no septadas presentes. Serie parenquimática de dos, tres y cuatro células. Radios exclusivamente uniseriados, de tipo heterogéneos III (según Tortorelli, 2009; Fig. 7H).
Plano longitudinal radial: Punteaduras vaso-radiales con rebordes visibles, similares a punteaduras intervasculares en tamaño y forma a lo largo de las células radiales. Células procumbentes, cuadradas y erectas mezcladas (Fig. 7I).
Colección de referencia: Descripción de los caracteres diagnósticos de Tierra del Fuego (Figs. 8 y 9).
Familia Asteraceae
Chiliotrichum diffusum (Material bibliográfico de referencia consultado: Solari, 1992; Piqué, 1999; Caruso 2012).

Fig. 7 Lámina de cortes histológicos de la estepa arbustiva del Golfo San Jorge (cont.); corte transversal (izq), longitudinal tangencial (centro) y radial (der): A-C, Schinus sp. D-F, Fabiana imbricata. G-I, Lycium sp. Figura en color en la versión en línea http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/1020/125
Plano transversal: Anillos de crecimiento demarcados por hileras de fibras achatadas en el leño tardío y una hilera de vasos dispuestos tangencialmente en el leño temprano. Porosidad difusa. Vasos casi exclusivamente solitarios (Fig. 8A).
Plano longitudinal tangencial: Radios uni, bi-triseriados. Punteaduras intervasculares alternas. Fibras con punteaduras con rebordes conspicuos (fibrotraqueidas). Estratificación parcial en fibrotraqueidas y vasos (Fig. 8B).
Plano longitudinal radial: Placa de perforación simple, escalariforme y transicionales entre ambas. Engrosamientos helicoidales en los elementos del vaso. Punteaduras vaso-radiales con rebordes visibles, similaresa las punteaduras intervasculares (Fig. 8C).
Familia Winteraceae
Drimys winteri (Material bibliográfico de referencia consultado: Solari, 1992; Piqué, 1999; Tortorelli, 2009).
Plano transversal: Anillos no demarcados, madera sin vasos, exclusivamente fibrotraqueidas de paredes delgadas. Parénquima apotraqueal difuso o ausente (Fig. 8D).
Plano longitudinal tangencial: Radios de dos tamaños distintos. Radios uni (bi) seriados. Radios multiseriados de 4 a 10 series. Radios con porciones uniseriadas tan largas como las biseriadas, heterogéneos. Radios mayores a 1 mm de largo. Radios multiseriados agregados. (Fig. 8E).
Plano longitudinal radial: Placas de perforación escalariformes. Fibrascon punteaduras con rebordes conspicuos (de forma areolada) (fibrotraqueidas), opuestas y alternas. Punteaduras fibrotraqueida-radio con bordes conspicuos similares a las de las fibrotraqueidas entre sí. Radios con células erectas (Fig. 8F).
Familia Celastraceae
Maytenus magellanica (Material bibliográfico de referencia consultado: Wheeler et al., 1989; Solari, 1992; Piqué, 1999; Caruso, 2012).
Plano transversal: Anillos de crecimiento demarcados por una hilera de fibras achatadas. Porosidad difusa. Vasos exclusivamente solitarios, de contorno angular. Vasos en una banda tangencial en el leño temprano y disposición radial (en menor medida diagonal) en el leño tardío. Fibras de paredes delgadas a gruesas. Parénquima axial apotraqueal difuso (Fig. 8G).
Plano longitudinal tangencial: Radios uni, bi-triseriados. Serie parenquimática de tres-cuatro y cinco-ocho células. Fibras con punteaduras con rebordes conspicuos (fibrotraqueidas). Punteaduras intervasculares alternas de forma redondeada. De acuerdo con la IAWA (Wheeler et al., 1989) las punteaduras intervasculares son escalariformes (Fig. 8H).
Plano longitudinal radial: Placa de perforación simple y engrosamientos helicoidales en los elementos del vaso. Punteaduras vaso-radiales con rebordes visibles, similar a punteaduras intervasculares en tamaño y forma a lo largo de la célula radial. Radios con células erectas marginales (Fig. 8I).
Familia Ericaceae
Gaultheria mucronata (Material bibliográfico de referencia consultado: Solari, 1992).
Plano transversal: Anillos de crecimiento no demarcados o ausentes, porosidad difusa. Vasos exclusivamente solitarios. Fibras de paredes delgadas a gruesas. Vasos solitarios, de contorno angular, también múltiples radiales de dos a cuatro y escasos agrupados. Fibras de paredes delgadas a gruesas. Parénquima paratraqueal escaso (Fig. 8J).
Plano longitudinal tangencial: Radios comúnmente uniseriados, también bi-tri y parcialmente tetraseriados. Fibras con punteaduras con rebordes conspicuos (fibrotraqueidas). Punteaduras intervasculares de escalariformes a opuestas. Engrosamientos helicoidales en vasos (Fig. 8K).
Plano longitudinal radial: Placas de perforación escalariformes, de diez a veinte barras. Engrosamientos helicoidales a lo largo del cuerpo del elemento del vaso. Punteaduras vaso-radiales con rebordes visibles; similares a punteaduras intervasculares en tamaño y forma a lo largo de la célula radial. Radios con el cuerpo de células procumbentes, erectas y cuadradas mezcladas y una a tres hileras de células marginales erectas (Fig. 8L).
Familia Proteaceae
Embothrium coccineum (Material bibliográfico de referencia consultado: Piqué, 1999; Caruso, 2012).
Plano transversal: Anillos de crecimiento demarcados por porosidad semicircular e hileras de fibras achatadas. Vasos formando bandas tangenciales de disposición ulmoide en el leño temprano y diagonal a dendrítica en el leño tardío. Vasos agrupados. Fibras de paredes delgadas a gruesas. Parénquima axial escaso o ausente (Fig. 8M).
Plano longitudinal tangencial: Radios de dos tamaños distintos, uni y multi-seriados (de cuatro a diez series). Puntaedauras intervasculares alternas. Fibras con punteaduras con rebordes conspicuos (fibrotraqueidas) (Fig. 8N).
Plano longitudinal radial: Placas de perforación simples, horizontales a oblicuas. Punteaduras vaso-radiales con rebordes visibles, similar a punteaduras intervasculares en tamaño y forma a lo largo de la célula radial. Radios con células procumbentes (Fig. 8Ñ).
Familia Grossulariaceae
Ribes magellanicum (Material bibliográfico de referencia consultado: Solari, 1992; Piqué, 1999; Caruso, 2012).

Fig. 8 Lámina de cortes histológicos de Tierra del Fuego; corte transversal (izq), longitudinal tangencial (centro) y radial (der): A-C, Chiliotrichum diffusum. D-F, Drimys winteri. G-I, Maytenus magellanica. J-L, Gaultheria mucronata. M-Ñ, Embothrium coccineum. O-Q, Ribes magellanicum. Figura en color en la versión en línea http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/1020/1254
Plano transversal: Anillos de crecimiento demarcados por hileras de fibras achatadas. Porosidad difusa. Vasos dispuestos en bandas tangenciales de disposición ulmoide. Vasos principalmente solitarios, escasamente múltiples cortos de dos, tres y agrupados. Fibras de pared delgada a gruesa. Parénquima axial apotraqueal difuso (Fig. 8O).
Plano longitudinal tangencial: Radios multiseriados de cuatro a diez series. Células envolventes en radios. Punteaduras intervasculares opuestas a escalariformes. Fibras con punteaduras con rebordes conspicuos (fibrotraqueidas). Placa de perforación escalariforme de entre diez a veinte barras. De acuerdo con Piqué (1999) también placa de perforación simple y transicionales entre ambas (Fig. 8P).
Plano longitudinal radial: Punteaduras vaso-radiales con rebordes visibles, similar a punteaduras intervasculares en tamaño y forma a lo largo de la célula radial. Radios con células procumbentes, cuadradas y erectas mezcladas (Fig. 8Q).
Fagaceae
Nothofagus antarctica (Material bibliográfico de referencia consultado: Solari, 1992; Piqué, 1999; Caruso, 2012).
Plano transversal: Anillos de crecimiento demarcados por una o más hileras de fibras achatadas, porosidad difusa. Vasos en disposición diagonal y/o radial. Vasos solitarios (principalmente en leño temprano), múltiples radiales (de dos a seis) y agrupados. Múltiples radiales de más de cuatro vasos comunes. Fibras de pared delgada a gruesa. Parénquima axial apotraqueal difuso (Fig. 9A).
Plano longitudinal tangencial: Radios exclusivamente uniseriados. Punteaduras intervasculares opuestas y escalariformes, de forma oval y areoladas. Tortorelli (2009) registra fibras libriformes tabicadas (Fig. 9B).
Plano longitudinal radial: Placas de perforación simples, oblicuas, con un apéndice terminal. Radios con un cuerpo de células procumbentes (y/o cuadradas, y cuatro o más hileras de células erectas). Punteaduras vaso-radiales con rebordes muy reducidos hasta con apariencia de simples: horizontales (escalariformes, muy estrechas) a verticales (en empalizada). Piqué (1999) observa, además de las simples, placas de perforación a escalariformes y de transición entre ambas (Fig. 9C).
Nothofagus betuloides (Material bibliográfico de referencia consultado: Solari, 1992; Piqué, 1999; Caruso, 2012).
Plano transversal: Anillos de crecimiento demarcados por una o más hileras de fibras achatadas, porosidad semicircular a difusa. Vasos con tendencia a formar una banda tangencial en leño temprano, y con disposición diagonal y/o radial en leño tardío. Vasos solitarios, múltiples radiales y agrupados. Múltiples radiales de más de cuatro vasos menos comunes que en el caso anterior. Fibras de pared delgada a gruesa. Parénquima axial apotraqueal difuso (Fig. 9D).
Plano longitudinal tangencial: Radios exclusivamente uniseriados. Punteaduras intervasculares opuestas y escalariformes de forma oval alargada y areolada. Fibras con punteaduras con rebordes conspicuos (fibrotraqueidas). De acuerdo a la IAWA (Wheeler et al., 1989) se registran engrosamientos helicoidales en los elementos del vaso (Fig. 9E).
Plano longitudinal radial: Placas de perforación simples, oblicuas, con un apéndice terminal. Radios con células procumbentes, cuadradas y erectas mezcladas. Punteaduras vaso-radiales con rebordes muy reducidos hasta con apariencia de simples: horizontales (escalariformes, muy estrechas) a verticales (en empalizada). Según Tortorelli (2009) presenta radios cortos. Piqué (1999) observa también placas de perforación transicionales entre escalariformes y simples (Fig. 9F).
Nothofagus pumilio (Material bibliográfico de referencia consultado: Solari, 1992; Piqué, 1999; Caruso, 2012).
Plano transversal: Anillos de crecimiento demarcados por una o más hileras de fibras achatadas, porosidad semicircular a difusa. Vasos con tendencia a formar una banda tangencial en leño temprano y con disposición diagonal y/o radial en el leño tardío. Vasos solitarios angulares, múltiples radiales y agrupados. Múltiples radiales de más de cuatro vasos menos comunes que en N. antartica. Fibras de pared delgada a gruesa. Parénquima axial apotraqueal difuso (Fig. 9G).
Plano longitudinal tangencial: Radios exclusivamente uniseriados. Punteaduras intervasculares opuestas y escalariformes.Fibras con punteaduras con rebordes conspicuos (fibrotraqueidas). De acuerdo con Tortorelli (2009) presenta fibras libriformes (Fig. 9H).
Plano longitudinal radial: Placas de perforación simples, muy oblicuas, con un apéndice terminal. Engrosamientos helicoidales en los vasos. Radios con hileras de células procumbentes e hileras de células erectas (y cuadradas). Punteaduras vaso-radiales con rebordes muy reducidos hasta con apariencia de simples: redondeadas o angulares. Piqué (1999) observa, además de las simples, placas de perforación escalariformes y de transición entre ambas (Fig. 9I).
Familia Berberidaceae
Berberis empetrifolia (Material bibliográfico de referencia consultado: Piqué, 1999; Andreoni, 2010, 2015).
Plano transversal: Anillos de crecimiento demarcados. Vasos en bandas tangenciales con disposición ulmoide en el leño temprano, y diagonal y/o radial a dendrítica en el leño tardío. Vasos principalmente solitarios, de contorno angular, algunos vasos agrupados. Fibras de paredes gruesas. Parénquima axial ausente o extremadamente escaso (Fig. 9J).
Plano longitudinal tangencial: Placa de perforación simple, oblicua. Punteaduras intervasculares alternas, poligonales. Fibras con punteaduras con rebordes conspicuos (fibrotraqueidas). Fibras con punteaduras en las paredes radiales y tangenciales. Radios de dos tamaños distintos. Radios uni a tri-seriados (comúnmente bi-seriados) y radios grandes cuatro a diez-seriados, homogéneos. Radios largos de altura mayor a 1 mm. Andreoni (2015) observa también radios agregados. Cristales prismáticos presentes (Fig. 9K).
Plano longitudinal radial: Placas de perforación simples. Engrosamientos helicoidales en los elementos del vaso. Radios con células procumbentes y erectas. Punteaduras vaso-radiales con reborde visible, similares a las punteaduras intervasculares en tamaño y forma a lo largo de la célula radial. Cristales prismáticos presentes (Fig. 9L).
Berberis ilicifolia (Material bibliográfico de referencia consultado: Solari, 1992; Piqué, 1999).
Plano transversal: Anillos de crecimiento demarcados, porosidad semicircular. Vasos en bandas tangenciales con disposición ulmoide en el leño temprano, y disposición diagonal, radial y/o dendrítica en el leño tardío. Vasos solitarios, de contorno angular, y vasos agrupados. Fibras de paredes gruesas. Parénquima axial ausente o extremadamente escaso (Fig. 9M).
Plano longitudinal tangencial: Placas de perforación simples, horizontales a oblicuas. Fibras con punteaduras con rebordes conspicuos (fibrotraqueidas). Punteaduras intervasculares alternas poligonales y opuestas. Radios largos, de altura mayor a 1 mm. Radios grandes cuatro a diez-seriados, homogéneos. Células envolventes en radios. Radios agregados (Fig. 9N).
Plano longitudinal radial: Engrosamientos helicoidales en los elementos del vaso. Radios con células procumbentes y erectas. Punteaduras vaso-radiales con rebordes visibles, similares a las punteaduras intervasculares en tamaño y forma a lo largo de la célula radial (Fig. 9Ñ).
Berberis microphylla (Tierra del Fuego; Material bibliográfico de referencia consultado: Ancibor & Pérez de Micou, 1995, 2002; Piqué, 1999; Capparelli et al. 2009; Andreoni, 2010; Caruso, 2012; Ciampagna, 2015).
Plano transversal: Anillos de crecimiento demarcados, porosidad semicircular. Vasos en bandas tangenciales con disposición ulmoide en el leño temprano, y diagonal, radial y/o dendrítica en el leño tardío. Vasos principalmente solitarios, aunque también agrupados. Fibras de paredes gruesas. Parénquima axial ausente o extremadamente escaso (Fig. 9O).
Plano longitudinal tangencial: Radios grandes multiseriados (de 4 a 10 series), homogéneos. Radios largos hasta 1 mm. Punteaduras intervasculares alternas y opuestas. Fibras con punteaduras con rebordes conspicuos (fibrotraqueidas). Fibras con punteaduras en las paredes radiales y tangenciales (Fig. 9P).
Plano longitudinal radial: Placas de perforación simples, oblicuas. Punteaduras intervasculares principalmente alternas poligonales, más raramente opuestas. Engrosamiento helicoidal a lo largo de los elementos del vaso. Radios con células procumbentes y erectas (Fig. 9Q).

Fig. 9 Lámina de cortes histológicos de Tierra del Fuego; corte transversal (izq), longitudinal tangencial (centro) y radial (der): A-C, Nothofagus antarctica. D-F, Nothofagus betuloides. G-I, Nothofagus pumilio. J-L, Berberis empetrifolia. M-Ñ, Berberis ilicifolia. O-Q, Berberis microphylla. Figura en color en la versión en línea http://www. ojs.darwin.edu.ar/index.php/darwiniana/article/view/1020/1254
Clave dicotómica. Taxones de la estepa arbustiva Golfo San Jorge
1. CT: Madera con variaciones cambiales. CLTg: vasos con engrosamiento espiralado .............................................. 2
1. CT: Madera sin variantes cambiales .................................................................................................................. 4
2(1). CT: Maderas con crecimiento secundario anómalo con paquetes separados de xilema-floema (distribuidos en una matriz de fibras o parénquima) anillos no demarcados, vasos exclusivamente solitarios; CLTg: placas de perforación y punteaduras escalariformes, maderas sin radios (descripción Andreoni, 2010) ................................................ Mulinum
2. CT: Maderas con floema incluso ............................................................................................................................... 3
3(2). CT: anillos demarcados por parénquima en bandas de 2 a 3 células. Floema incluso tipo difuso, dentro del parénquima paratraqueal y en forma centrípeta a cada conjunto de vasos ............................................... Atriplex
3. CT: anillos no demarcados, floema incluso tipo difuso concéntrico dentro de bandas de parénquima de más de 4 células de ancho ......................................................................................................................... Suaeda
4(1). CT: anillos demarcados de transición abrupta, traqueidas de contorno poligonal, la mayoría solitaria, fibrotraqueidas. CLTg: traqueidas con placas de perforación foraminada, fibrotraqueidas, campos de cruzamiento cupresoide, radios de 4 a 5 células de ancho (madera de Gimnosperma, descripción Andreoni, 2010) .......... Ephedra
4. Otras combinaciones de caracteres (maderas de Angiospermas, sin tráqueas) ......................................................... 5
5(4). CT: disposición de los vasos radial/diagonal al menos en leño tardío (a veces algo dendrítico). CLTg: Fibras con punteaduras conspicuas y punteaduras vaso-radio con rebordes conspicuos en vista tangencial, se observan punteaduras en todos los tipos de células (incluso en el CLR). Cuerpo del radio predominantemente con células elongadas dispuestas en varias direcciones, aunque también puede haber algunas células redondeadas ........................................................ 6
5. Otras combinaciones de caracteres. CT: disposición de los vasos por lo general tangencial en leño temprano y diagonal/radial a dendrítica en leño tardío. CLTg: no se presentan células elongadas en distintas direcciones en los cuerpos de los radios multiseriados. Si bien puede haber fibrotraqueidas o fibras con punteaduras visibles, las punteaduras en CLTg y CLrd no son tan abundantes como se aprecia en Asteraceae, ni se dan en todos los tipos de células ............................................................................................................................ 12
6(5). Maderas con anillos no demarcados ..................................................................................................................... 7
6. Maderas con anillos demarcados ............................................................................................................................... 9
7(6). Radios comúnmente 1-3 seriados, a veces parcialmente 4 seriados (descripción Andreoni 2015b) ....... Grindelia
7. Radios con más de 4 células de ancho frecuentes ..................................................................................................... 8
8(7). Parénquima axial confluente .................................................................................................................... Baccharis
8. Parénquima axial difuso ........................................................................................................................ Senecio
9(6). Radios extremadamente escasos o no visibles ..................................................................................................... 10
9. Radios conspicuos o visibles, parénquima axial ausente o escaso, radios agregados .............................. Chuquiraga
10(9). Parénquima axial difuso y difuso en agregados, poros solitarios .................................................. Lepidophyllum
10. Parénquima axial ausente o escaso ......................................................................................................................... 11
11(10). Placas escalariformes y simples, poros múltiples cortos .............................................................. Nardophyllum
11. Placas simples, poros solitarios .................................................................................................................. Nassauvia
12(5). Parénquima en bandas y/o parénquima paratraqueal presente ........................................................................... 13
12. Sin parénquima en bandas ni parénquima paratraqueal. El parénquima es ausente, escaso o apotraqueal difuso/difuso en agregados ........................................................................................................................................................................ 20
13(12). Radios exclusivamente uniseriados ......................................................................................................... Lycium
13. Radios no exclusivamente uniseriados ................................................................................................................... 14
14(13). Radios mayores a 1 mm presentes, radios 1-3 seriados y 4-10 seriados .......................................................... 15
14. Radios mayores a 1 mm ausentes, radios 1 a 4 seriados ........................................................................................ 16
15(14). Células por serie parenquimática fusiformes y 2 por serie .................................................................... Adesmia
15. Células por serie parenquimática: fusiformes, 3 a 4, 5 a 8 ................................................................. Erythrostemon
16(14). Radios heterogéneos con colas uniseriadas y cuerpo multiseriado presentes .................................... Mulguraea
16. Todos los radios homogéneos de células redondeadas ........................................................................................... 17
17(16). Sin parénquima en bandas ................................................................................................................................ 18
17. Con parénquima en bandas ..................................................................................................................................... 19
18(17). Parénquima aliforme (descripción Andreoni y Capparelli 2012) ........................................... Prosopis alpataco
18. Parénquima vasicéntrico (descripción Andreoni 2015a) ............................................................................. Condalia
19(17). Con engrosamiento helicoidal en vasos ..................................................................................... Prosopidastrum
19. Sin engrosamiento helicoidal en vasos ........................................................................................ Prosopis denudans
20(12). Con estructura estratificada en vasos, parénquima y fibras (descripción de Andreoni 2015a) ................ Larrea
20. Sin estructura estratificada ...................................................................................................................................... 21
21(20). Radios exclusivamente uniseriados .................................................................................................................. 22
21. Radios no exclusivamente uniseriados ................................................................................................................... 23
22(21). Radios uniseriados heterogéneos, vasos con engrosamiento espiralado ............................................. Colliguaja
22. Radios uniseriados homogéneos, vasos sin engrosamiento espiralado ................................................... Fabiana
23(21). Radios mayores a 1mm presentes, radios de 2 tamaños, 1-3 seriados y 4-10 seriados .................................... 24
23. Radios mayores a 1 mm ausentes, radios de 1 tamaño, 1-3 seriados ............................................................. Schinus
24(23). Con patrón ulmoide ................................................................................................................................ Berberis
24. Sin patrón ulmoide (descripción Andreoni & Capparelli, 2012) .......................................................... Ochetophylla
Clave dicotómica para los taxones de Tierra del Fuego
1. CT: Anillos de crecimiento demarcados .................................................................................................................... 2
1. CT: Anillos de crecimiento no demarcados o ausentes .............................................................................................. 7
2(1). CTg: Radios exclusivamente uniseriados, punteaduras intervasculares opuestas y escalariformes ..................... 3
2. CTg: Radios no exclusivamente uniseriados ............................................................................................................. 4
3(2). CR: Ausencia de engrosamientos helicoidales ................................................... Nothofagus antarctica/betuloides
3. CR: Presencia de engrosamientos helicoidales ............................................................................. Nothofagus pumilio
4(2). CTg: Presencia de fibrotraqueidas ........................................................................................................................ 5
4. CTg: Ausencia de fibrotraqueidas ............................................................................................................................. 6
5(4). CTg: Radios uni-bi-triseriados. CR: perforaciones simples, escalariformes y transición entre ambas ........................................................................................................................................................................ Chiliotrichum diffusum
5. CTg: Radios multiseriados (de 4 a 10 series). CR: Perforaciones simples ..................................................... Berberis
6(4). CR: Perforaciones simples .................................................................................................................................... 7
6. CR: Perforaciones escalariformes de 10 a 20 barras, punteaduras vaso-radiales con rebordes visibles ................................................................................................................................................................................ Ribes magellanicum
7(6). CT: Parénquima apotraqueal difuso. CTg:Radios uni, bi-triseriados ................................... Maytenus magellanica
7. CT: Parénquima paratraqueal confluente. CTg: Radios uniseriados y multiseriados (de 4 a 10 series) .......................................................................................................................................................................... Embothrium coccineum
8(1). CT: Madera sin vasos, exclusivamente traqueidas de paredes delgadas. CTg: Radios multiseriados (de 4 a 10 series) .......................................................................................................................................... Drimys winteri
8. CT: Madera de porosidad difusa. CTg: Radios comúnmente uniseriados ................................ Gaultheria mucronata
Casos de Aplicación: sitios arqueológicos del LAPS
Análisis antracológico del sitio Palo Alto.
En la submuestra analizada del sitio Palo Alto, que proviene del sondeo 1, se identificaron ocho taxones (Fig. 10A, C, Fig. 11A-H). Entre ellos prevalece, tanto en abundancia como ubicuidad, Lycium seguido por Atriplex/Suaeda, Schinus, Senecio/Baccharis, Discaria, Chuquiraga, Prosopis y Berberis (Fig. 10A). La presencia de carbones no determinables se debe principalmente a que se encontraban dentro de la categoría de tamaño más pequeña (2-4,9 mm), y en algunos casos también se vieron afectados por alteraciones como la vitrificación.
En cuanto al porcentaje de la frecuencia de estos taxones por nivel, se observa que Lycium predomina en todos los niveles a excepción del nivel cuatro, en el cual Atriplex/Suaeda lo supera. Este último se encuentra en frecuencias altas en todos los niveles (Fig. 10C). El primero de los géneros está presente en todos los niveles mientras que Atriplex/Suaeda, Senecio/Baccharis y Schinus están presentes en cuatro de los niveles. La densidad calculada de los carbones respecto al volumen de sedimento de cada nivel permite observar un incremento a mayor profundidad, especialmente a partir del nivel 2. La riqueza de taxones también aumenta a partir del segundo nivel y se mantiene constante en los niveles 4 y 5 (Tabla 1).
Del total de los fragmentos de carbón analizados el 57,29 % (N=106) presentan alguna o varias alteraciones. Éstas se concentran principalmente en Atriplex/Suaeda y, en menor porcentaje, en Lycium (Fig. 10C). Las grietas y roturas del parénquima en Atriplex/Suaeda son las alteraciones predominantes, seguido por la vitrificación en ambos taxones.
Análisis antracológico del sitio Cabo San Pablo 2017 (CSP2017).
El análisis taxonómico del sitio CSP2017 ha permitido identificar 3 taxones (Franch et al., 2020). En cuanto a la composición del conjunto, éste se encuentra marcadamente dominado por Nothofagus antartica/betuloides, y de forma secundaria encontramos en el registro antracológico Nothofagus pumilio. Ribes magellanicum aparece de forma esporádica como un único fragmento en capa C. Los no determinables están relacionados a la presencia de alteraciones (Tabla 2).
Los restos leñosos carbonizados se distribuyen en todas las capas. El porcentaje de la frecuencia de taxones por capa, indica que la presencia de N. antarctica/betuloides destaca en la capa B (Fig. 10B, C, Fig. 11I-K).
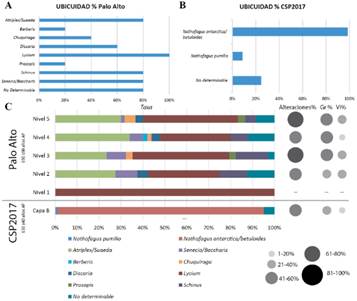
Fig. 10 A, ubicuidad (%) por taxón en Sitio Palo Alto. B, ubicuidad (%) por taxón en Capa B de CSP2017 (tomado de Franch et al., 2020). C, frecuencia relativa (%) por taxón y alteraciones (%) de Palo Alto y CSP2017 por nivel o capa (CSP2017 tomado de Franch et al., 2020). En Alteraciones% se incluyen los porcentajes de grietas y roturas del parénquima (Gr%), vitrificación (Vi%) y otras alteraciones secundarias o esporádicas (ataque de insectos xilófagos, fractura laminar y presencia de hifas o micelios). Figura en color en la versión en línea http://www.ojs.darwin.edu.ar/ index.php/darwiniana/article/view/1020/1254
Del total de fragmentos de carbón analizados el 41,78% (N=742) presentan alguna o varias alteraciones. Éstas se concentran principalmente en Nothofagus antarctica/betuloides y, en menor porcentaje, en Nothofagus pumilio (Fig. 10C).
Las grietas radiales de contracción son las alteraciones predominantes, seguido por la vitrificación, el ataque de insectos xilófagos y la fractura laminar. Finalmente, la presencia de hifas o micelios aparece de forma esporádica.

Fig. 11 Imágenes de los planos anatómicos de los carbones arqueológicos; transversal (izq), longitudinal tangencial (centro) y radial (der): A-C, Lycium sp. del sitio Palo Alto. D-E, Atriplex/Suaeda del sitio Palo Alto. F-H, Schinus sp. del sitio Palo Alto. I-K, Nothofagus antarctica/betuloides del sitio CSP2017. L-N, Nothofagus pumilio del sitio CSP2017. Figura en color en la versión en línea http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/1020/1254
Discusión
En este trabajo se presentó la descripción de leños del LAPS, correspondientes a 18 taxones de la estepa arbustiva del Golfo San Jorge y doce del ecotono bosque-estepa, el bosque caducifolio y el bosque mixto de Tierra del Fuego. Para aquellas especies descritas por otros autores en otras ubicaciones geográficas, se reconocen algunas diferencias posiblemente debido a las características ambientales de tipo de suelo y clima. Es el caso, por ejemplo, de Atriplex lampa donde aquí se reconocieron anillos de crecimiento demarcado por hileras de parénquima en bandas de 2 a 3 células mientras que para la especie colectada en Mendoza por Andreoni (2015a) no se observó demarcación de anillos. En Baccharis darwinii, en este estudio, se describe porosidad difusa y parénquima axial paratraqueal confluente. En cambio, Guerra et al. (2012) describen los cortes histológicos de B. linearis, B. obovata y B. salicifolia del ecotono bosque estepa de Chubut e identifican porosidad circular a semicircular en las dos primeras y semicircular en la última. Además de parénquima paratraqueal confluente observan parénquima vasicéntrico. En este sentido, la colecta de ejemplares para el área de estudio y el desarrollo de una clave dicotómica es relevante antes de realizar un estudio antracológico.
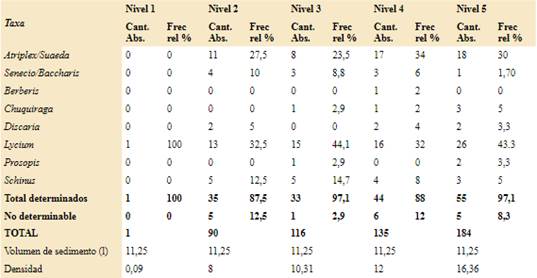
Tabla 1 Conteo absoluto y frecuencia relativa porcentual de carbones para los litros de sedimento analizados por nivel en el sitio Palo Alto.
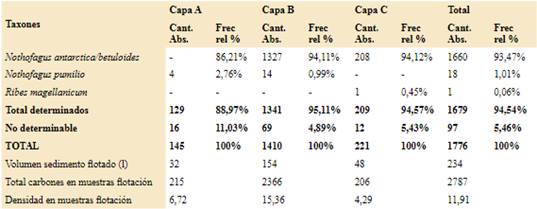
Tabla 2 Conteo absoluto y frecuencia relativa porcentual de carbones para los litros de sedimento analizados por capa en el sitio CSP2017.
La clave dicotómica, entonces, resultó una herramienta de gran importancia para la identificación de los carbones de dos sitios arqueológicos de la costa norte de Santa Cruz y la faja central de Tierra del Fuego. En ambos sitios se identificaron taxonómicamente 11 taxones, 10 de ellos representados en la clave dicotómica y uno de ellos consultado a través de bibliografía específica (Andreoni, 2014, 2015b).
En el sitio Palo Alto, las especies identificadas taxonómicamente se encuentran, en la actualidad, próximas al sitio. Sobre el litoral se encuentran Atriplex/Suaeda, Senecio/Baccharis, Schinus, Chuquiraga y Lycium, y más distante, hacia el flanco de meseta y cañadones, Prosopis denudans. Los estudios polínicos realizados en la meseta central podrían sugerir que para el Holoceno tardío final la vegetación sería análoga a la actual (De Porras, 2010).
La ubicuidad indica que Lycium se registró en todos los niveles del sondeo 1, seguidos por Atriplex/Suaeda y Senecio/Baccharis. El conjunto de fragmentos de carbón se encontró disperso y en el registro arqueológico no se observaron concentraciones de carbones o estructuras de fogón. Sin embargo, los restos de cáscara de huevo de ñandú y algunos restos óseos recuperados presentaron evidencias de exposición al fuego. Las maderas de los taxones identificados, de acuerdo con sus propiedades biofísicas, representarían una combinación de maderas de rápido encendido (por ejemplo Lycium, Senecio/Baccharis) con otras que producirían mayor cantidad de brasa y duración (por ej. Schinus, Prosopis) (Ciampagna, 2015). Entre las alteraciones que se observaron en los carbones, no se identificaron aquellos rasgos diagnósticos de temperaturas por encima de los 400 °C (por ej. contracciones radiales y en anillo al mismo tiempo en Lycium, Ciampagna, 2015). La ausencia de hongos y marcas causadas por insectos xilófagos podría indicar que se utilizó madera en buen estado y que no habría pasado un largo período antes de su uso luego de la muerte.
La baja densidad de restos antracológicos y la inexistencia de rasgos de fogones podrían explicarse a priori por procesos tafonómicos y de remoción como los que se observan en el conjunto óseo, altamente meteorizado. Zubimendi y colaboradores (2010), sugieren que los restos arqueológicos habrían quedado expuestos a la intemperie durante un periodo prolongado de tiempo antes de quedar enterrados, y luego podrían haber sido sometidos intensamente a agentes de alteración postdepositacional, como animales fosoriales, y recientemente también a procesos antrópicos (por ej. presencia de caminos, explotación de canteras de arena, entre otras). También este contexto podría explicarse por la limpieza de los fogones, o los fuertes vientos que podrían haber dispersado los carbones antes de su entierro.
Por otra parte, el análisis taxonómico de la madera carbonizada del sitio CSP2017 permitió identificar taxones arbóreos del bosque (Nothofagus antarctica/betuloides, Nothofagus pumilio) y otro de porte arbustivo presente en sotobosque y/o espacios abiertos (Ribes magellanicum). La ubicuidad nos indica que el género Nothofagus estuvo asociado a la mayor parte de los contextos de los pisos de ocupación del sitio, mientras que Ribes magellanicum aparece con un uso espacialmente más restringido. La concordancia de los taxones identificados con las descripciones de la vegetación actual y los registros polínicos del Holoceno tardío indican que las especies leñosas utilizadas como combustible estaban disponibles en el entorno de los sitios (Franch et al., 2020).
Los restos leñosos carbonizados de CSP2017 presentan un alto porcentaje de fragmentos con alteraciones, causadas principalmente por grietas de contracción que se concentran en Nothofagus sp. Las carbonizaciones experimentales de Nothofagus antarctica de Tierra del Fuego sugieren que la abertura en anillos y radios se da en fragmentos con humedad (8 % y el 12 %) o sin humedad, a los 700 ºC y no a los 400 ºC, por lo que estas alteraciones se habrían producido en carbonización a temperaturas mayores de 400 ºC (Franch et al., 2020). La presencia de hongos y marcas causadas por insectos xilófagos es baja, lo que podría indicar que en ambos casos se empleaba madera en buen estado y que no habría pasado un largo periodo antes de su uso luego de la muerte.
En ambos sitios, los carbones dispersos dan cuenta de eventos de combustión asociados a contextos arqueológicos con presencia de recursos faunísticos marinos y terrestres en ocupaciones de cazadores recolectores para el Holoceno tardío final. Estos grupos seleccionaron maderas de porte arbustivo en Palo Alto, y de porte arbóreo y arbustivo proveniente del bosque en CSP2017, ambos ambientes próximos a los sitios. En este sentido, la existencia de este ecotono marino terrestre cobra sentido en la selección, procesamiento y consumo de distintos recursos que están disponibles el litoral atlántico e insular como en la franja interior inmediata. Este trabajo contribuye al estudio de las maderas utilizadas como combustibles en sitios arqueológicos donde predominan acumulaciones de moluscos y explotación de otros recursos costeros y se suma a otros realizados por Ortega & Marconetto (2009, 2012), Caruso Fermé & Gómez Otero (2016), Hammond & Ciampagna (2019) y Franch et al. (2020).
En síntesis, la realización de cortes histológicos y el estudio de las maderas es un paso necesario y valioso en el armado de la colección de referencia. El aumento de la riqueza en especies en la formulación de claves dicotómicas permitirá mayores niveles de resolución en los estudios antracológicos de sociedades cazadoras recolectoras de la Patagonia.
Conclusiones
La creación de una colección de referencia que representara la mayor cantidad de especies leñosas disponibles en el entorno de los sitios arqueológicos, para la estepa arbustiva del Golfo San Jorge y para las diferentes regiones de la Isla Grande de Tierra del Fuego (ecotono bosque-estepa, el bosque caducifolio y el bosque mixto) permitió describir las características anatómicas de cada madera y realizar distintas claves dicotómicas según el área trabajada. Este estudio agilizó la determinación taxonómica de los carbones arqueológicos, favoreció a realizar una correcta interpretación de este registro y permitió realizar interpretaciones e inferencias sobre las sociedades humanas y su relación con el entorno vegetal. Sin duda, constituirá una herramienta que facilitará nuevos estudios antracológicos en sitios arqueológicos de las regiones estudiadas.

