











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkAn important component in the evaluation of avian population dynamics is the estimation of fle-dgling productivity (Newton 1998). Mist-netting has been widely used as a method to estimate this de-mographic parameter (e.g., Peach et al. 1998, Kaiser and Berthold 2004, Mazerolle et al. 2005, DeSante et al. 2019, Saracco et al. 2019), although it may have several biases (Dunn and Ralph 2004). For instance, capture probability may vary by age class (Ballard et al. 2004, Nur et al. 2004), affecting the estimation of young pro-portion in the population (e.g., Hussell 2004, Kornegay et al. 2018). Several studies have reported a parallel relationship between the number of young captured and fledgling productivity, but not for all species (du Feu and McMeeking 1991, 2004, Nur and Geupel 1993). These results may be consequence of different post-fledgling dispersal processes among species (e.g., Chase et al. 1997, Anders et al. 1998, Vega Rivera et al. 1998, Daly et al. 2019). Thus, mist-net capture of young represents local productivity in some species and regional productivity in others (Dunn and Ralph 2004). Therefore, it is necessary to evaluate whether produc-tivity indices from mist-netting accurately reflect local productivity in studies of bird population dynamics.
The aforementioned studies about the relations-hip between young capture and fledgling productivity were conducted on species from Europe and North America, but to our knowledge this relationship has not been studied for South American bird species. However, the number of young captured has been considered a surrogate of productivity in some demo-graphic studies in the Neotropics (e.g., Sagario 2010, Presti et al. 2018). The aim of our research was to eva-luate if there is an association between young capture rate and the number of fledglings found during inten-sive nest monitoring of White-crested Elaenia (Elae-nia albiceps chilensis), a long-distance migrant (Bravo et al. 2017) and the most abundant bird species in the Andean-Patagonian Forest (Ippi et al. 2009, Cueto and Gorosito 2018), where it has an important role in forest regeneration (Cavallero et al. 2013, Bravo et al. 2015).
METHODS
Study site
We conducted our study at Cañadón Florido Ranch (42°55’S, 71°21’W), Chubut Province, Argentina. The vegetation of the area corresponds to the Val-divian Forest Province of the Andean Region (Morro-ne 2001). The forest at the study site is dominated by Maitén (Maytenus boaria) and Laura (Schinus patagoni-cus) trees. The understory is mainly dominated by Calafate (Berberis microphylla) shrubs. The forest canopy is low, averaging a height of 5 m. The climate is cha-racterized by cold and wet winters and mild but dry summers. Most precipitation falls as rain and snow during fall and winter (April-September). The annual mean precipitation in the region is 704 mm. Summer and winter mean temperatures are 13.4° and 1.8 °C, respectively. All climate data are from the "Río Per-cey” meteorological station (for the 1998-2017 pe-riod, Hidroeléctrica Futaleufú S.A.).
Young surveys
We captured White-crested Elaenias with mist-nets within a 20-ha plot during three breeding sea-sons, between October 2015 and March 2018. Nets were 12 m long with a 38 mm mesh size and were opened during the first 4-5 h after sunrise. We set two groups of 10 nets each within the plot. Each group was set in different zones into the forest, 200 m apart. Nets within each group were set 50 to 70 m apart and opened at least twice per month. The order of net group sampling was set randomly, with 10 or 15 days between sampling events, depending on wea-ther conditions (i.e., avoiding rainy or windy days). We banded all birds with one numbered aluminum band and with unique combinations of three colored bands. The sampling effort during the period Octo-ber-March of each breeding season were: 2015-2016: 18 net-days with 853 net-hours; 2016-2017: 16 net-days with 586 net-hours and 2017-2018: 18 net-days with 743 net-hours. We identified all young (hatched during each breeding season of the study) based on the absence of the white coronal patch and the bei-ge color of wing bars (Jaramillo 2003, Pearman and Areta 2020).
Nest monitoring
We searched for nests from late October to late February of the 2015-2018 breeding seasons in the same 20-ha plot where we did the mist-netting sur-vey, following the method proposed by Martin and Geupel (1993). We monitored nests following stan-dardized protocols (Ralph et al. 1993) and visited them daily until nesting was successful (i.e., at least one nestling left the nest) or failed. We classified causes of nest failure as destruction of the nest (e.g., by falling branches or cattle activity), nestling starva-tion, unfertilized eggs and predation. We conside-red that a nest was preyed if an active nest was seen empty before the estimated fledgling date or when we found all eggs broken inside or around the nest. We banded all nestlings before they fledged (at the age of 11 days after hatching).
Data analysis
We estimated abundance of young using capture rate as the number of captures per 100 net-hours (Karr 1981). Annual young abundance was estimated as the average capture rate of young considering all sample events during each breeding season. We estimated the annual production of fledglings as the total number of fledglings divided by the total number of active nests found during each breeding season of the study. Inferences from statistical analysis and demographic interpretation must be cautious, as po-tential biases may arise from the small sample size. Thus, we used exploratory data analysis to evaluate the association between estimated annual young abundance and annual production of fledglings. Since both variables were measured on different units, the comparison between them can change depending on the y-axis scale. To avoid this inconvenience, we standardizad both variables using a Z standard score (Zar 2010) to present a graph with both variables and a unique y-axis, allowing a safe comparison between them. Moreover, we ran a Pearson Correlation Analy-sis using Z standard score to evaluate the association between annual young abundance and annual pro-duction of fledglings.
RESULTS AND DISCUSSION
During the study we captured 87 young Whi-te-crested Elaenias and found 84 nests, from which 40 nestlings fledged. Of these nests, 33.3% were suc-cessful and 63.1% failed due to predation. The avera-ge annual young capture rate (± SD) was 13.57 ± 1.85 individuals per 100 net-hours and the average annual production of fledglings (± SD) was 0.48 ± 0.09 indivi-duals per nest. Annual young capture rate was highest during the breeding season of 2015-2016 and lowest in the 2016-2017 breeding season (Table 1). Annual production of fledglings showed a similar pattern of chan-ges among breeding seasons (Table 1). We observed a closed parallel relationship between Z standard scores of young capture rate and fledgling production among years (Fig. 1, r = 0.998, P = 0.0386, n = 3). This coincides with that reported for species in the Northern He-misphere (du Feu and McMeeking 1991, 2004, Nur and Geupel 1993), but to our knowledge the result presen-ted here is the first for a Neotropical passerine.
After White-crested Elaenias fledged, they were fed by the parents and formed family groups that moved across their breeding territories for almost the entire time between leaving the nest and the moment they dispersed from the territory they hatched in (Gorosito CA, personal observation). At the end of the summer (March), young White-crested Elaenias began their fall migration to the winter quarters (Gorosito 2020). Based on these evidences, we are confident that annual young abundance reflects local fledgling productivity.
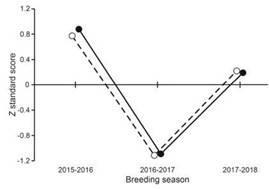
Figure 1: Z standard score of annual young abundance (black dots) and total fledgling productivity (white dots) of White-crested Elaenia (Elaenia albiceps chilensis) during three breeding seasons in the forest-steppe eco-tone at Cañadón Florido Ranch, Chubut Province, Argentina
Some studies indicated that climate change could affect migratory bird populations due to an advance in the phenology of food resources on breeding grounds and, consequently, food abundance would be scarce for successful reproduction when migratory birds arrive (Schoech and Hahn 2007, Both et al. 2010). The detection of this mismatch requires long-term studies that monitor the variation in fledgling pro-ductivity. However, nest monitoring involves intense field work and may be costly to support during mul-tiple years. Conversely, monitoring bird populations using captures could be more cost-effective, especia-lly in remote areas or for studies with little financial support, as is common in the Neotropic.
The use of capture rates to estimate current pro-ductivity is useful only if it reflects what actually happens in the wild. There are several biases when young capture rate is evaluated. Capture probabili-ty may vary by age class affecting the estimation of young proportion in the population (Kornegay et al. 2018, Daly et al. 2019). Missed visits during a banding season schedule in a locality could also affect such estimation (Miles et al. 2007, Cave et al. 2008). Although there are no studies for White-crested Elaenia that evaluate the probability of capture depending on age, we have not found a differential capture rate among adults of different ages (Gorosito et al. 2020). Howe-ver, it is an issue that should be studied, since it would be of great importance to establish the usefulness of mist net sampling to determine the proportion of young in the population, which is essential for the as-sociation described in our research to be valid.
We simultaneously monitored nests and captured young White-crested Elaenias during three breeding seasons, thus the small sample size implies being careful when considering the use of young capture rates as a surrogate of fledgling productivity. Long-term studies on bird populations are scarce in the Neotropic (e.g., Cueto et al. 2008, Rozzi and Jiménez 2013), and particularly there are no studies about the relationship analyzed in our research. Hence, more studies evaluating the relationship between young capture rate and fledgling productivity for other Neo-tropical bird species are necessary to generalize the results obtained here for White-crested Elaenia.
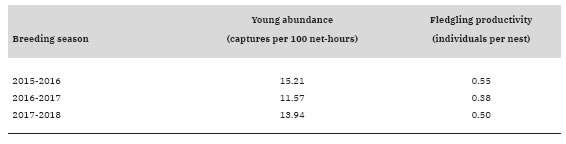
Table 1: Annual young abundance and total fledgling productivity of White-crested Elaenia (Elaenia albiceps chilensis) during three breeding seasons in the forest-steppe ecotone at Cañadón Florido Ranch, Chubut Province, Argentina.
ACKNOWLEDGEMENTS
We thank the Roberts family for allowing us to work at Cañadón Florido. For constructive reviews of the manuscript, we thank Taty Sagario and Alex Jahn. Research was founded by the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET, Argentina), National Geographic Society (USA), the Bergstrom Award of the Association of Field Ornitho-logists (USA) and the Frangois Vuilleumier Fund of the Neotropical Ornithological Society (USA). Birds were captured with permission of Dirección de Fauna y Flora Silvestre, Provincia del Chubut (Argentina).
Recibido: 27 de Septiembre de 2021
Aceptado: 18 de Febrero de 2022

