










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRIA. Revista de investigaciones agropecuarias
On-line version ISSN 1669-2314
RIA. Rev. investig. agropecu. vol.38 no.3 Ciudad Autónoma de Buenos Aires Dec. 2012
ARTÍCULOS
Determinación del requerimiento de frío y de calor en duraznero [Prunus persica (L.) Batsch.] mediante un modelo de correlación
Chaar, J1.; Astorga, D1.
1Instituto Nacional de Tecnología Agropecuaria. Estación Experimental Agropecuaria Junín, Mendoza Isidoro Busquets s/n (5572). La Colonia, Junín, Mendoza Correo electrónico: jchaar@correo.inta.gov.ar
Recibido 07 de julio de 2012
Aceptado 05 de septiembre de 2012
Publicado online 06 de noviembre de 2012
RESUMEN
Los Árboles frutales de hoja no persistente poseen un período de reposo entre los ciclos vegetativos. Para salir de reposo invernal, las yemas deben acumular primero frío (endodormancia) y luego calor (ecodormancia). El objetivo de este trabajo fue determinar el requerimiento de frío y de calor en cinco cultivares de duraznero mediante un método de correlación, con datos de fecha de plena floración y temperatura del aire de siete años, en el este de Mendoza. El requerimiento de frío varió entre 914 y 1262 unidades de frío, mientras que el de calor fluctuó entre 2177 y 6490 horas grado de crecimiento. Excepto en cv. "Prima", la variabilidad entre años del requerimiento de frío (coeficiente de variación, CV: 18,3 a 18,9%) fue mayor que la correspondiente al requerimiento de calor, (CV: 6,9 a 13,9%). Los requerimientos de calor disminuyeron en años de mayor acumulación de frío e indicaron una compensación. El modelo utilizado, permite disponer de información mÁs precisa sobre los requerimientos térmicos de los cultivares, y así poder realizar una zonificación que racionalice el cultivo y maximice su aprovechamiento, en las Áreas agroclimÁticamente adecuadas.
Palabras clave: Dormancia; Fenología; Heladas; Quiescencia; Temperatura.
ABSTRACT
Deciduous fruit trees have a rest period between vegetative cycles. For winter dormancy release, buds need to accumulate chill (endodormancy) and then heat (ecodormancy). The aim of this study was to determine chilling and heat requirements of five peach cultivars by means of a correlation model, with full bloom and temperature data collected during seven years, in eastern Mendoza. Chilling requirement ranged from 914 to 1262 chill units, whereas heat requirement fluctuated between 2177 and 6490 growing degree hours. Except in cv. "Prima", chill requirement variability between years (CV: 18.3 to 18.9%) was greater than that of heat requirement (CV: 6.9 to 13.9%). Heat requirement diminished in years of high chilling accumulation. The model proposed in this study provides more accurate information about thermal requirements of cultivars in order to rationalize crop distribution in the environment.
Keywords: Dormancy; Phenology; Freeze; Quiescence; Temperature.
INTRODUCCIÓN
Las plantas perennes se distinguen de otras en su capacidad de suspender y retomar el crecimiento recurrentemente en respuesta a condiciones ambientales, a menudo estacionales. La posibilidad de formar yemas y experimentar ciclos de crecimiento y dormancia ha sido una adaptación evolutiva de suma importancia (Rohde y Bhalerao, 2007). Dentro de las plantas perennes, los Árboles frutales de hoja no persistente poseen un período de reposo que coincide en nuestras latitudes, en la provincia de Mendoza, con parte del otoño, el invierno y comienzos de la primavera, según la especie y cultivar. Para salir de dicho reposo, las yemas (florales y vegetativas) deben primero estar expuestas a temperaturas bajas, en el período considerado de acumulación de frío, y luego a temperaturas moderadas, en el período de acumulación de calor. Estas dos etapas diferenciadas, son conocidas como endodormancia y ecodormancia, respectivamente (Lang et al., 1987).
La endodormancia, es liberada mediante la exposición a bajas temperaturas a largo plazo. La acumulación de frío restablece la capacidad de crecer pero no promueve el crecimiento (Rohde y Bhalerao, 2007). Posteriormente, un vez que la yema acumuló la cantidad de calor necesaria (propia del cultivar), se desencadenan los procesos de floración o de brotación vegetativa, según el tipo de yema. La disponibilidad de agua y nutrientes también regulan la salida de la ecodormancia (Lang et al., 1987). Para florecer, es necesario que la yema cubra los requerimientos cuantitativos en cada una de las dos etapas. Mientras que en zonas geogrÁficas con climas de invierno poco riguroso ciertos cultivares pueden no cubrir su requerimiento de frío, el requerimiento de calor generalmente se satisface, variando la fecha de floración en función de la velocidad de acumulación. Por ello, los cultivares mÁs plÁsticos, en cuanto a posibilidad de lograr cosechas aceptables en zonas geogrÁficas diversas, son aquellos que poseen bajo requerimiento de frío y elevado requerimiento de calor. Mientras que el bajo requerimiento de frío les permite poder satisfacerlo aún en zonas con inviernos poco fríos, el elevado requerimiento de calor se manifiesta en la floración tardía, lo cual posibilita evadir heladas tardías.
Existen diversos métodos para estimar las necesidades térmicas para la salida del reposo invernal en especies caducifolias, con antecedentes tanto a campo (Anderson et al., 1986; Alonso et al., 2005), como en laboratorio (Citadin et al., 2001; Naor et al., 2003; Gariglio et al., 2006; Alburquerque et al., 2008; Aslamarz et al., 2009; Rahemi y Pakkish, 2009; Harrington et al., 2010).
Debido a la falta de conocimiento de los mecanismos bioquímicos y fisiológicos que controlan el requerimiento de frío, casi todos los modelos han sido desarrollados empírica o estadísticamente para ajustar las respuestas (principalmente las fechas de floración) de especies arbóreas a condiciones climÁticas locales (Fan et al., 2010). En cada cultivar, la determinación de la fecha de comienzo de la ecodormancia, y mediante ésta la estimación de los requerimientos de frío y de calor, puede realizarse a través de la relación entre la temperatura media promedio de cierto período (dentro del período de reposo) y la fecha de plena floración, considerando una serie de años. Si bien no hay un límite abrupto entre los períodos de acumulación de frío y de acumulación de calor, puede considerarse que el comienzo de la ecodormancia ocurre cuando las bajas temperaturas retrasan la fecha de floración (Alonso et al., 2005). Excepto para cerezo (Prunus avium L.), (Tersoglio y Naranjo, 2009; Naranjo y Tersoglio, 2010) no existe a nivel local información sobre las necesidades térmicas otoño-invernales de los frutales cultivados, entre ellos el duraznero. Por lo tanto, el objetivo de este trabajo fue determinar el requerimiento de frío y de calor en durazneros conserveros mediante un método de correlación, en el este de Mendoza.
MATERIALES Y MÉTODOS
Sitio y material experimental
El estudio se llevó a cabo en la Estación Experimental Agropecuaria Junín INTA, ubicada en La Colonia, Junín, Mendoza (33° 6' 57,5" Sur; 68° 29' 4" Oeste; 653 msnm), en una colección de cultivares de duraznero para conserva, durante el período 1998-2007 (excepto los años 2001 y 2005).
Los cultivares evaluados fueron: "Dee-Six", "Maria Serena", "Ouro", "Prima" y "Ross".
El marco de plantación era de 5 m x 5 m, siendo el portainjerto "Nemaguard" (Prunus persica L. Batsch.) y las plantas conducidas en vaso.
Determinación de la fecha de plena floración (PF)
En cada uno de los años de estudio, para cada cultivar se promedió información de cuatro plantas, determinÁndose la fecha de plena floración mediante la estimación visual a nivel de planta entera del 50% de flores abiertas, en el estadio F según Baggiolini (1952).
Registro de temperatura del aire
Para cada uno de los años de estudio, los datos de temperatura horaria de abril a septiembre se obtuvieron a partir de una estación meteorológica ubicada a 300 m de la parcela experimental, perteneciente a la Dirección de Agricultura y Contingencias ClimÁticas del Gobierno de Mendoza.
Estimación de la fecha de transición entre endodormancia y ecodormancia
Se utilizó el método propuesto por Alonso et al. (2005), con los datos de la fecha de plena floración y de la temperatura media del aire de los años 1998, 1999, 2000, 2002, 2003, 2004 y 2006. El fundamento del método se basa en el efecto diferente de las temperaturas frías y templadas sobre la fecha de plena floración, según su incidencia sea durante la endodormancia o la ecodormancia. Se considera iniciada la ecodormancia cuando el coeficiente de correlación de Pearson entre la fecha de plena floración y el promedio de temperaturas medias del período considerado (5, 10, 15, 20, 25 o 30 días) es significativo, luego de una serie continua de coeficientes negativos. Los coeficientes de correlación menores de -0,754 son significativos (a = 0,05), debido a que las correlaciones se han obtenido de un conjunto de datos de siete años (Hoshmand, 1988).
CÁlculo de los requerimientos de frío y de calor
A partir de las temperaturas horarias se calcularon las unidades de frío (UF) y las horas grado de crecimiento (GDH ºC), mediante el modelo de Richardson et al. (1974). El primer día de acumulación de frío se consideró el día siguiente a la última acumulación diaria negativa de UF (la fecha media de inicio de la endodormancia fue el 6 de mayo), mientras que el último día de acumulación fue el estimado por el modelo de correlación. El promedio de las UF de los siete años se consideró como la necesidad de frío de cada cultivar.
Las GDH ºC se calcularon desde la fecha de transición entre endo-ecodormancia y la fecha de plena floración, promediando los valores de siete años para determinar los requerimientos de calor de cada cultivar. Una GDH ºC, se define como una hora a una temperatura 1 ºC por arriba de la temperatura base de 4,5 ºC. Las temperaturas por encima de 25 ºC se asumen iguales a 25 ºC, y es la mayor acumulación posible por cada hora de 20,5 GDH ºC (Alonso et al., 2005).
Validación del modelo
Se utilizaron datos correspondientes al año 2007, los cuales no se incluyeron para la elaboración del modelo.
AnÁlisis estadístico
El anÁlisis estadístico de los datos se realizó con el programa InfoStat/Profesional (Universidad Nacional de Córdoba, 2006), mediante las herramientas de anÁlisis de correlación de Pearson y anÁlisis de regresión lineal.
RESULTADOS Y DISCUSIÓN
La distribución de los coeficientes de correlación de Pearson entre la fecha de plena floración y el promedio de temperaturas medias varió según la longitud del período considerado. En la figura 1, se observa dicha variación para el cultivar "Maria Serena". Para la estimación de los requerimientos térmicos se utilizó el período de 20 días, donde todos los cultivares evaluados presentaron coeficientes de correlación negativos significativos.
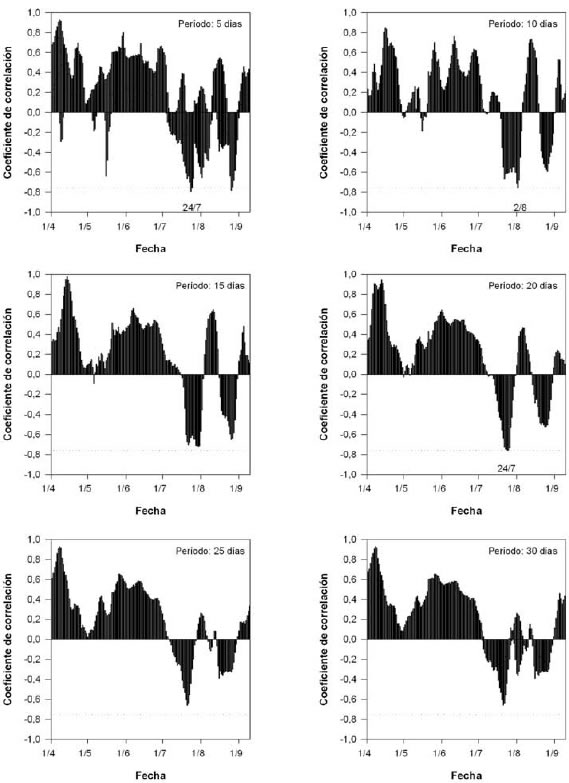
Figura 1. Distribución de los coeficientes de correlación de Pearson entre la fecha de plena floración y el promedio de temperaturas medias, según la amplitud del período considerado. Duraznero cv. "Maria Serena". La línea discontinua corresponde al mínimo valor significativo (r = -0,754).
Mediante la observación de los coeficientes de correlación entre la temperatura media promedio y la fecha de plena floración, se pudo determinar el momento de transición entre endodormancia y ecodormancia (figura 2). En los cultivares evaluados se observó un mismo patrón de variación del coeficiente de correlación. En la etapa "I", las yemas se encontraron en paradormancia y la inhibición del crecimiento se originó a partir de una señal bioquímica proveniente de otra parte de la planta (Lang et al., 1987). El coeficiente de correlación positivo significa que cuanto menor es la temperatura, antes ocurre la floración en la siguiente primavera, debido al adelantamiento de la caída natural de hojas y consecuentemente la salida de la paradormancia (entrada prematura a la endodormancia). La etapa "II", coincidió temporalmente con la caída natural de hojas, evento que representa la transición entre para y endodormancia (Cesaraccio et al., 2004). En la etapa "III", los coeficientes de correlación positivos indicaron un atraso de la siguiente floración cuando las temperaturas fueron elevadas y señalaron que las yemas se encontraban en endodormancia. La etapa "IV", es transicional, mientras que en "V" ocurría la ecodormancia, donde temperaturas elevadas promueven el proceso de apertura floral, en la etapa"VI" (figura 2a).
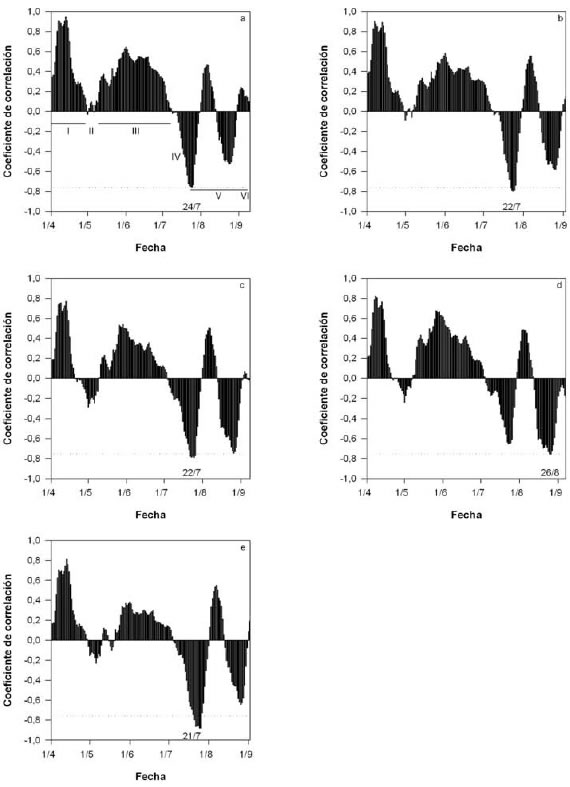
Figura 2. Distribución de los coeficientes de correlación de Pearson entre la fecha de plena floración y el promedio de temperaturas medias de 20 días. a) cv. "Maria Serena"; b) cv. "Dee-Six"; c) cv. "Ouro"; d) cv. "Prima"; e) cv. "Ross". La línea discontinua corresponde al mínimo valor significativo (r = -0,754).
Si bien en el cv. "Prima" el coeficiente de correlación negativo fue significativo recién el 26 de agosto (un mes después que los demÁs cultivares evaluados), a fines de julio presentó un pico de correlación negativa, de manera similar a los otros materiales (figura 2d). Por lo tanto, la fecha de transición entre la endodormancia y la ecodormancia fue similar en los cinco cultivares, ocurrido en el último tercio del mes de julio.
En la figura 3, se puede observar el significado del coeficiente de correlación negativo y significativo entre la temperatura media del período considerado y la fecha de plena floración para el cv. "Ross". A medida que aumenta la temperatura media del período, se adelanta la floración, lo que indica que la yema ya superó la etapa de acumulación de frío y se encuentra en la ecodormancia. Sin embargo, y de manera similar a lo encontrado por Alonso et al. (2005), la correlación entre las variables evaluadas vuelve a fluctuar luego del inicio de la ecodormancia (etapa "V", figura 2a). La disminución en la magnitud del coeficiente de correlación puede deberse a que el efecto de adelantamiento de la floración, provocado por temperaturas elevadas a la salida del invierno, es proporcionalmente menor que el de temperaturas moderadas. En efecto, según Richardson et al. (1974), las temperaturas superiores a 25 ºC sólo acumulan 20,5 GDH ºC, estando a 25 ºC la óptima acumulación (mayor cociente GDH ºC / ºC). Luego, el coeficiente de correlación comienza a ser positivo (aunque no significativo) y, posiblemente, durante este período de fluctuación ocurran otros procesos no contemplados por los autores, en donde la relación entre el momento de floración y los períodos de temperatura media esté influida por otros factores ambientales o intrínsecos de la planta. Al respecto, Cesaraccio et al. (2004), mencionan que ninguno de los modelos estudiados para Populus tremula L. y Salix chrysocoma Dode predijeron bien la ruptura de las yemas, por lo cual es probable que algún otro factor distinto a la temperatura esté involucrado en la salida de la dormancia.
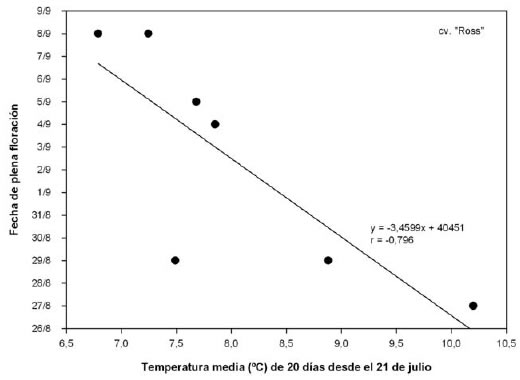
Figura 3. Relación entre la temperatura media promedio de 20 días desde el inicio de la ecodormancia y la fecha de plena floración, en duraznero cv. "Ross".
Excepto en cv. "Prima", la variabilidad entre años del requerimiento de frío (coeficiente de variación, CV: 18,3 a 18,9%) fue mayor que la correspondiente al requerimiento de calor (CV: 6,9 a 13,9%). Según Fan et al. (2010), los requerimientos de frío y de calor de un determinado cultivar no son constantes, sino que dependen de la interacción genotipo x ambiente y varían con la evolución de las temperaturas de cada año y de cada lugar. Factores como luz, temperatura y nutrición pueden alterar los requerimientos de frío y calor entre años (Rahemi y Pakkish, 2009). Sin embargo, al promediar una serie de años es posible diferenciar cultivares en base a sus requerimientos térmicos medios. El requerimiento de frío medio en cuatro de los cultivares de duraznero evaluados varió entre 914 y 946 UF (tabla 1). El requerimiento de frío determina, en gran medida, los límites climÁticos para el adecuado desempeño del cultivo. AdemÁs de la variación geogrÁfica en cuanto a la oferta de frío otoño-invernal, la acumulación de frío podría verse afectada a futuro debido al aumento de las temperaturas mínimas (Cesaraccio et al., 2004; Altieri y Nicholls, 2009; Minetti, 2010). Alonso et al. (2005), mencionan requerimientos de 358 a 481 UF en almendro, cumpliéndose la endodormancia en apenas 32 a 40 días, según el cultivar. Por el contrario, la necesidad de calor en almendro fue muy superior a los valores observados en duranzero (tabla 1) variando entre 5345 y 9352 GDH ºC.
Tabla 1. Fecha media de transición entre endo y ecodormancia, requerimiento de frío y de calor, y fecha media de plena floración de los cultivares de duraznero evaluados, según el modelo de correlación (Alonso et al., 2005).
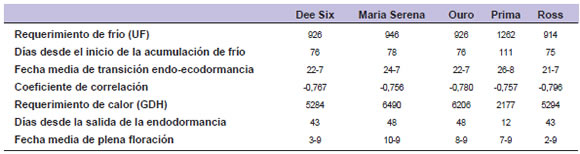
Alburquerque et al. (2008), encontraron en cerezo requerimientos térmicos de 397 a 1001,5 UF y de 7326 a 9450 GDH ºC. Rahemi y Pakkish (2009), observaron requerimientos de calor en pistacho (Pistacia vera L.) de 8852 a 15420 GDH ºC. La variabilidad del requerimiento de calor fue mayor que la de la necesidad de frío (CV: 34% y 15%, respectivamente), al comparar entre los distintos cultivares estudiados. A pesar de la similar fecha de transición de endo a ecodormancia entre los cultivares evaluados, la mayor variabilidad en el requerimiento de calor determina diferencias en la fecha de plena floración. Esta información es importante cuando se requiere utilizar cultivares de floración mÁs tardía, para evitar o disminuir el daño causado por heladas primaverales. Similares resultados fueron observados por Alonso et al. (2005).
Al validar el modelo con los datos climÁticos y de fecha de plena floración correspondientes al año 2007 (observaciones a campo, datos no incluidos en la elaboración del modelo), se encontró un estrecho ajuste (y = 2,1983 x - 49316; R2= 0,95). Sin embargo, la fecha de plena floración estimada mediante el modelo fue entre 6 y 14 días mÁs atrasada que la observada a campo, según el cultivar. Por lo tanto, el modelo debería modificarse en lo que respecta al criterio del comienzo de acumulación de frío. En la figura 4, se observa que el comienzo de acumulación de frío al utilizar el modelo de Richardson et al. (1974) es ampliamente variable entre años. En años en que hay acumulaciones negativas de frío ya entrado el otoño (4 de mayo de 2007; -2,5 UF), el modelo de correlación atrasa la fecha de plena floración estimada con respecto a lo observado a campo. Posiblemente, las plantas entren en la endodormancia previamente a la última acumulación negativa de UF, por lo que no es suficiente un pequeño período de temperaturas cÁlidas para anular el frío acumulado. Según el modelo dinÁmico (Fishman et al., 1987), que asume una base bioquímica de la ruptura de la endodormancia, un primer paso produce un intermediario reversible de la sustancia para la salida de la endodormancia, y el segundo paso fija el intermediario mediante una transición irreversible, por lo cual el frío acumulado previo a un período cÁlido puede ser efectivo. Otro criterio para establecer el inicio del reposo invernal es mencionado por Cesaraccio et al. (2004), mediante un modelo cuyo comienzo de acumulación de frío coincide con determinada etapa fenológica, como la caída natural de hojas en otoño.
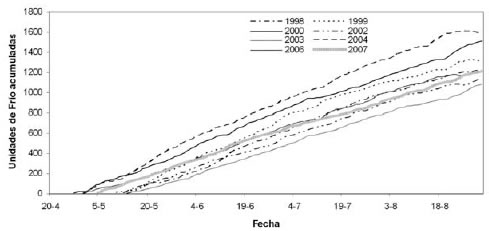
Figura 4. Evolución de la acumulación de Unidades de Frío (según Richardson et al., 1974) en los años estudiados. Junín, Mendoza.
En la figura 5, se observa que el cv. "Prima" posee una relación entre el calor y el frío acumulados distinta al resto de los materiales (según los datos obtenidos mediante el modelo de correlación), por lo cual no se incluyó en el siguiente anÁlisis. A pesar de la débil relación entre las UF y las GDH ºC acumuladas (R2=0,36), se observó una tendencia a disminuir los requerimientos de calor en años de mayor acumulación de frío. Similares resultados, fueron encontrados por Couvillon y Erez (1985). De manera similar a lo mencionado para almendro (Alonso et al., 2005), la fecha de plena floración en duraznero dependió principalmente del requerimiento de GDH ºC.
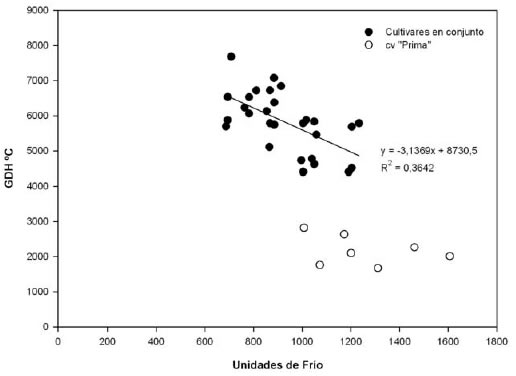
Figura 5. Relación entre el calor (GDH ºC) y el frío (UF) acumulados en los años estudiados, según cultivar de duraznero.
En la figura 6, se observa cómo la fecha de plena floración se atrasa al aumentar el requerimiento de calor en duraznero (excepto en el cv. "Dee-Six"), mientras que el bajo requerimiento de frío se relaciona con una floración temprana sólo cuando el requerimiento de calor también es bajo. Por lo tanto, cuando hubo una menor acumulación de frío y el requerimiento de calor fue mayor, se retrasó la fecha de plena floración. La menor duración de la endodormancia (debida al retraso en la salida de la paradormancia) podría ser beneficiosa para disminuir el riesgo de daño por heladas tardías.
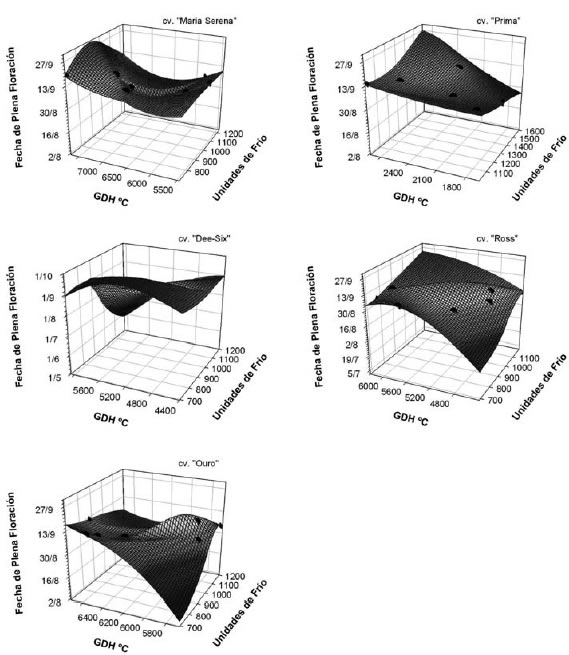
Figura 6. Requerimiento de frío, requerimiento de calor y fecha de plena floración de los cultivares de duraznero evaluados.
A partir de la variabilidad interanual encontrada en los requerimientos de frío y de calor de un mismo cultivar en un mismo sitio, se manifiesta la importancia de evaluar los materiales vegetales con respecto a la relación entre sus requerimientos térmicos (ya que no son constantes), o bien promediar los valores obtenidos de una serie extensa de años.
CONCLUSIONES
El modelo utilizado posibilita disponer de información mÁs precisa sobre los requerimientos térmicos de los cultivares, y así poder realizar una zonificación que racionalice el cultivo y maximice su aprovechamiento en las Áreas agroclimÁticamente adecuadas. Adicionalmente, la determinación de la variación interanual en los requerimientos de frío y de calor y su relación con las condiciones de temperatura media del aire, permitirían diseñar prÁcticas de manejo tendientes a retrasar la floración y, consecuentemente, a disminuir el riesgo de daño por heladas tardías.
AGRADECIMIENTOS
A la Dirección de Agricultura y Contingencias ClimÁticas del Gobierno de Mendoza, por el suministro de los datos climÁticos.
BIBLIOGRAFÍA
1. ALBURQUERQUE N.; GARCÍA-MONTIEL F.; CARRILLO A.; BURGOS L. 2008. Chilling and heat requirement of sweet cherry cultivars and the relationship between altitude and the probability of satisfying the chill requirements. Environ. Exp. Bot. 64, 162-170. [ Links ]
2. ALONSO J. M.; ANSÓN J. M.; ESPIAU M. T.; SOCIAS I COMPANY R. 2005. Determination of endodormancy break in almond flower buds by a correlation model using the average temperature of different day intervals and its application to the estimation of chill and heat requirements and blooming date. J. Amer. Soc. Hort. Sci. 130(3), 308-318 [ Links ]
3. ALTIERI M.; NICHOLLS C. 2009. Cambio climÁtico y agricultura campesina: impactos y respuestas adaptativas. LEISA 24(4), 5-8. [ Links ]
4. ANDERSON J.; RICHARDSON E.; KESNER C. 1986. Validation of chill unit and flower bud phenology models for 'Montmorency' sour cherry. Acta Hort. 184, 71-78. [ Links ]
5. ASLAMARZ A.; VAHDATI K.; RAHEMI M.; HASSANI D. 2009. Estimation of chilling and heat requirements of some Persian walnut cultivars and genotypes. HortSci. 44(3), 697-701. [ Links ]
6. BAGGIOLINI, M. 1952. Stade repères du pecher. Revue Romande d'Agriculture, Viticulture et Arboriculture 4, 29. [ Links ]
7. CESARACCIO C.; SPANO D.; SNYDER R.; DUCE P. 2004. Chilling and forcing model to predict bud-burst of crop and forest species. Agricultural and Forest Meteorology 126, 1-13. [ Links ]
8. CITADIN I.; RASEIRA M.; HERTER F.; BAPTISTA DA SILVA J. 2001. Heat requirement for blooming and leafing in peach. HortSci. 36(2), 305-307. [ Links ]
9. COUVILLON G.; EREZ A. 1985. Influence of prolonged exposure to chilling temperatures on bud break and heat requirement for bloom of several fruit species. J. Amer. Soc. Hort. Sci. 110(1), 47-50. [ Links ]
10. FAN S.; BIELENBERG D.; ZHEBENTYAYEVA T.; REIGHARD G.; OKIE W.; HOLLAND D.; ABBOTT A. 2010. Mapping quantitative trait loci associated with chilling requirement, heat requirement and bloom date in peach (Prunus persica). New Phytologist 185, 917-930. [ Links ]
11. FISHMAN S.; EREZ A.M.; COUVILLON G. 1987. The temperature dependence of dormancy breaking in plants: Mathematical analysis of a two-step model involving cooperative transition. Journal of Theoretical Biology 124(4), 473-483. [ Links ]
12. GARIGLIO N.; GONZÁLEZ ROSSIA D.; MENDOW M.; REIG C.; AGUSTI M. 2006. Effect of artificial chilling on the depth of endodormancy and vegetative and flower budbreak of peach and nectarine cultivars using excised shoots. Sci. Hort. 108, 371-377. [ Links ]
13. HARRINGTON C.;GOULD P.; ST.CLAIR J. 2010. Modeling the effects of winter environment on dormancy release of Douglas-fir. Forest Ecology and Management 259, 798-808. [ Links ]
14. HOSHMAND A. 1988. Statistical methods for agricultural sciences. Timber Press, Portland, Or. [ Links ]
15. LANG G.; EARLY J.; MARTIN G.; DARRELL R. 1987. Endo, para, and ecodormancy: Physiological terminology and classification for dormancy research. HortSci. 22, 371-377. [ Links ]
16. MINETTI J. Cambio climÁtico real y simulado en el centro-sur de Sudamérica. Impactos en la agricultura. V Jornadas de actualización en Riego y Fertirriego, Mendoza, 11 al 13 de agosto de 2010. [ Links ]
17. NAOR A.; FLAISHMAN M.; STERN R.; MOSHE A.; EREZ A. 2003. Temperature effects on dormancy completion of vegetative buds in apple. J. Amer. Soc. Hort. Sci. 128(5), 636-641. [ Links ]
18. NARANJO G.; TERSOGLIO E. 2010. Características de la disponibilidad de frío invernal para cerezos en la zona norte de la provincia de Mendoza, Argentina. Agriscientia 27, 27-34. [ Links ]
19. RAHEMI M.; PAKKISH Z. 2009. Determination of chilling and heat requirements of pistachio (Pistacia vera L.) cultivars. Agr. Sci. China 8(7), 803-807. [ Links ]
20. RICHARDSON, E. A.; SEELEY S. D. Y WALKER D. R. 1974. A model for estimating the completion of rest for 'Redhaven' and 'Elberta' peach trees. HortSci. 9, 331-332. [ Links ]
21. ROHDE A.; BHALERAO R. 2007. Plant dormancy in the perennial context. Trends in Plant Science 12(5), 217-223. [ Links ]
22. TERSOGLIO E.; NARANJO G. 2009. Identificación del inicio de la ecodormancia en cerezo variedad "Bing". ITEA 105(4), 272-281. [ Links ]

