










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRIA. Revista de investigaciones agropecuarias
On-line version ISSN 1669-2314
RIA. Rev. investig. agropecu. vol.44 no.2 Ciudad Autónoma de Buenos Aires Aug. 2018
ARTÍCULOS
Desarrollo morfológico de Digitaria eriantha Steud. ssp. eriantha y Panicum coloratum L. var. Coloratum
González, F.A.1; Ventimiglia, P.1; Jouve, V.V.1; Ferri, C.M.1
1 Facultad de Agronomía, Universidad Nacional de La Pampa (UNLPam), Ruta Nac. Nº 35, km 334, 6300 Santa Rosa, La Pampa. Correo electrónico: varijouve@agro.unlpam.edu.ar
Recibido 29 de junio de 2016
Aceptado 01 de marzo de 2018
Publicado online 15 de agosto de 2018
RESUMEN
La cuantificación del desarrollo morfológico de una pastura resulta de interés a los fines de la aplicación de prácticas de manejo. El objetivo fue cuantificar el estado medio de desarrollo morfológico, analizar su variación a través del tiempo térmico y establecer la relación existente entre el estado medio de desarrollo y la proporción de lámina viva en pasturas de digitaria (Digitaria eriantha Steud.) y mijo perenne (Panicum coloratum L.). A principios de octubre, sobre pasturas de digitaria y mijo perenne se marcaron tres parcelas de 3,0 x 6,0 m en las que se cosecharon a ras del suelo 30 macollos por parcela, tomados al azar, cada siete días y durante 16 semanas. Se cuantificó el estado de desarrollo de cada macollo cortado a través de índices numéricos que relacionan el peso o la cantidad de macollos en cada estado con el peso (EMP) o la cantidad (EMC) total de macollos cortados, y se determinó la proporción de lámina viva. Los datos se sometieron a análisis de regresión lineal y modelos lineales mixtos para mediciones repetidas. Los índices EMP y EMC, en digitaria y mijo perenne, se relacionaron en forma estrecha (p<0,01). La duración del periodo vegetativo fue aproximadamente 250 grados días (base 10 ºC) más prolongado en digitaria que en mijo perenne, aunque alcanzaron el estado reproductivo al mismo tiempo. EMP fue un mejor predictor de la proporción de lámina viva que EMC, con un mayor R2 (0,89 vs. 0,83) y una menor raíz cuadrada del cuadrado medio del error. Los índices por peso y por conteo permitieron describir el estado de desarrollo morfológico medio y predecir la proporción de lámina viva de digitaria y mijo perenne.
Palabras clave: Gramíneas perennes de crecimiento estival; Madurez; Morfología; Proporción de lámina.
ABSTRACT
The measurement of pasture morphological development is critical to the purpose of implementing management practices. The objective was to quantify the mean stage of morphological development, to analyze its variation through thermal time and to establish the relationship between mean stage of development and green leaf blade proportion of digitgrass (Digitaria eriantha Steud.) and kleingrass (Panicum coloratum L.). In early October, three plots of 3.0 × 6.0 m were marked on a pasture of digitgrass, and another one of kleingrass, for ground-level cutting of 30 tillers per plot selected at random, every seven days during a period of 16 weeks of canopy growing season. Mean stage by weight (MSW), mean stage by count (MSC) and green leaf blade proportion were determined. Data were analyzed using linear regression and mixed linear models for repeated measurements. The MSW and MSC indexes, in digitaria and kleingrass, were highly related (p<0.01). The duration of the vegetative period was about 250 degree days (base 10 ºC) longer in digitgrass than in kleingrass, although they attained the reproductive stage at the same time. The MSW was a better predictor of green leaf blade proportion than MSC, with a higher R2 (0.89 vs. 0.83) and a lower root mean square error. Both MSW and MSC indexes allowed describing the mean morphological developmental stage and predicting green leaf blade proportion of digitgrass and kleingrass.
Keywords: Leaf blade proportion; Maturity; Morphology; Warm-season grasses.
INTRODUCCIÓN
En la región semiárida central, parte de la superficie original del caldenal fue reemplazada, en forma progresiva, por sistemas de producción tanto ganaderos como mixtos o agrícolas (Viglizzo et al., 2010). El sistema de producción predominante en las áreas donde se mantiene el bosque de caldén (Prosopis caldenia Burk.) es el de cría bovina. En general, estas áreas se encuentran sometidas a condiciones de sobrepastoreo que determinaron la presencia de superficies extensas con distinto grado de degradación florística, a partir de la invasión de especies indeseables para la ganadería, que afectan la capacidad de carga animal (Roberto et al., 2005; SAyDS, 2006). Todo ello determinó la necesidad de mantener e incrementar la receptividad animal y, para ello, se trabajó con éxito en la implantación de gramíneas C4 perennes exóticas como complemento forrajero del pastizal en áreas de desmonte, planicies y médanos (Ferri, 2014). Dentro de las gramíneas C4 estudiadas que se adaptan a este ambiente se destacan digitaria (Digitaria eriantha Steud. ssp eriantha) cv Irene y mijo perenne (Panicum coloratum L. var coloratum) cv Verde, dado sus niveles elevados de acumulación de materia seca y valor nutritivo (Stritzler et al., 1996; Petruzzi et al., 1996). Estas especies se caracterizan por ser insensibles a la longitud del día para iniciar la fase reproductiva (Nada, 1980; Loch et al., 2004), observándose que luego de iniciada, esta se extiende durante toda la estación de crecimiento. Ello determina que las pasturas de estas especies presenten macollos con diferente estado de madurez en forma simultánea (Moore y Moser, 1995) afectando su valor nutritivo (Coleman et al., 2004). Por ello, el conocimiento del estado medio de desarrollo de pasturas de digitaria y mijo perenne podría ser de utilidad a los fines de la investigación y para la toma de decisiones de manejo (por ej., inicio del pastoreo, cosecha para heno, pastoreo de los rebrotes, producción de semillas, aplicación de agroquímicos).
Varios autores han elaborado sistemas para cuantificar el estado de desarrollo morfológico de una pastura (Moore et al., 1991; Sanderson, 1992), tal que considere su variabilidad. De este modo se pretende superar la cuantificación visual, la cual tiende a sobreestimar el estado de madurez de la pastura debido a que los macollos en elongación y reproductivos son visualmente dominantes en la población (Mitchell et al., 1997). El desarrollo ontogénico de los macollos de gramíneas perennes se puede dividir, según Moore et al. (1991), en cuatro estados (vegetativo, elongación de tallos, reproductivo y madurez de las semillas) que a su vez se conforman por subestados, que se corresponden con eventos morfológicos específicos. Esta clasificación permitió, al asignar un número a cada estado y subestado, la elaboración de escalas numéricas que pueden ser usadas con fines cuantitativos, como la construcción de índices. Así, el estado morfológico medio de una pastura puede ser calculado a través de índices numéricos que surgen de relacionar el peso (EMP, estado medio por peso) o la cantidad (EMC, estado medio por conteo) de macollos en cada estado con el peso o la cantidad total de macollos de una muestra, respectivamente (Moore et al., 1991). El sistema propuesto por Moore et al. (1991) resulta de fácil aplicación y fue diseñado para ser utilizado en gramíneas forrajeras perennes, tanto del pastizal como cultivadas. A partir de este sistema, Sanderson (1992) elaboró una propuesta que también considera los estados parciales de desarrollo foliar, otorgando más precisión a la cuantificación.
A su vez, estos índices pueden ser utilizados para establecer posibles relaciones entre la madurez del forraje y variables de calidad o producción (Fick y Janson, 1990; Kalu y Fick, 1981; Smart et al., 2001). Por un lado, esto es de importancia dado que el estado de madurez de las pasturas, al momento del pastoreo o corte, es una de las variables con mayor efecto sobre la calidad del forraje (Nelson y Moser, 1994), facilidad de aprehensión, palatabilidad y el potencial del rebrote de las especies forrajeras luego de la defoliación (Briske, 1991). Por otro lado, las decisiones de manejo sustentadas en el estado madurez se consideran fiables, dado que los efectos acumulados del ambiente, edad de las plantas y genéticos se integran y expresan en el desarrollo morfológico de una especie (Buxton y Marten, 1989). El presente trabajo se realizó en pasturas de digitaria y mijo perenne con los siguientes objetivos: 1) cuantificar y comparar el estado medio de desarrollo morfológico, obtenido mediante índices cuantitativos de estado medio por peso (EMP) y por conteo (EMC), de ambas pasturas; 2) comparar entre estas especies la variación de los índices cuantitativos de desarrollo morfológico a través del tiempo térmico; 3) determinar la relación existente entre los índices EMP y EMC con la proporción de lámina viva.
MATERIALES Y MÉTODOS
El trabajo se efectuó en el campo experimental de la Facultad de Agronomía (UNLPam) (36°46’ lat. sur; 64°16’ long. oeste, 210 m s.n.m.), sobre pasturas monofíticas de digitaria (Digitaria eriantha Steud. ssp. eriantha) cv Irene y mijo perenne (Panicum coloratum L. var. coloratum) cv Verde, implantadas en 1996 sobre un suelo Haplustol entico (MO: 1,7%; P: 37,0 ppm; pH: 7,1). Ambas pasturas fueron utilizadas mediante pastoreo rotativo y con similar carga animal instantánea, con 3 a 4 periodos de pastoreo durante las estaciones de crecimiento anteriores a las mediciones. En cada pastura, previo al inicio del crecimiento de primavera, el material muerto proveniente de la estación de crecimiento anterior fue cortado a 5 cm de altura y removido. Luego, para cuantificar el desarrollo morfológico medio de la pastura se marcaron y cercaron tres parcelas de 3,0 x 6,0 m.
A partir del día 7 de octubre de 2014, cada siete días y durante 16 semanas, se tomaron al azar y cosecharon al ras del suelo 30 macollos por parcela (Sanderson, 1992). Para la selección de los macollos cosechados se utilizó un dispositivo denominado marco de puntos (Goodall, 1952), el cual se apoyó sobre la superficie del suelo cada 50 cm, siguiendo una dirección establecida al azar. Dicho dispositivo consta de un bastidor con cinco varillas que se desplazan hacia el suelo en un ángulo de 45°, así se seleccionaron los macollos interceptados por las varillas de cada extremo y central. Luego, se determinó el estado de desarrollo, de cada macollo cortado, mediante el uso de descriptores de características macromorfológicas externas (visibles o palpables) de acuerdo a la escala propuesta por Sanderson (1992), se agruparon según el estado en que se encontraban y se contaron. Posteriormente, se secaron en estufa (48 h a 55 °C), se pesaron y separaron en sus fracciones morfológicas lámina, tallo (incluyendo vaina e inflorescencia) y material muerto, para determinar la proporción de lámina viva. La escala de estados numéricos (tabla 1) utilizada describe cuatro eventos (estados) fenológicos sucesivos y subestados, incluyendo desde la emergencia de plántula o rebrote en la primavera (0,5) hasta el desprendimiento de semillas (35).
Tabla 1. Escala de estados numéricos para gramíneas perennes de crecimiento estival basada en descriptores morfológicos (Sanderson, 1992).
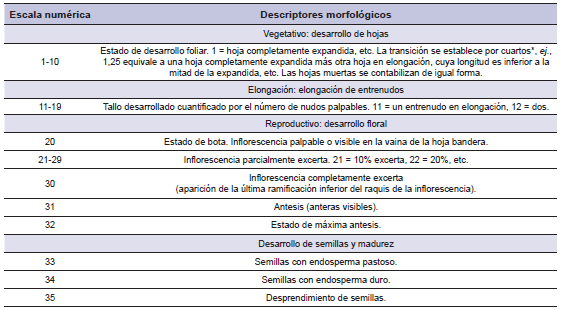
*En el caso que la longitud de la parte visible de la hoja en expansión excediera la longitud de la hoja expandida se estimó de la siguiente forma: [(n-1) + 0,9] donde n es el número de hojas en expansión y expandidas (Gustavsson, 2011).
Para determinar el estado medio de la pastura se utilizaron los índices por peso (EMP) y por conteo (EMC) de acuerdo a los modelos propuestos por Kalu y Fick (1981):
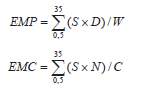
donde: S = número de estado (de 0,5 a 35 de acuerdo a escala; tabla 1),
D = peso seco de los macollos en el estado S,
W = peso seco total de la muestra de los 30 macollos,
N = número de macollos en el estado S,
C = número total de macollos en la muestra (n = 30).
Se determinó el desvío estándar de EMC (SEMC) para cada especie y en cada fecha de corte, a los efectos de estimar la variación alrededor de EMC dentro de la población de macollos siguiendo la ecuación propuesta por Moore et al. (1991):
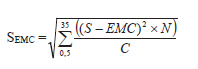
En tanto que, el desvío de EMP no se estableció dado que es función del producto de dos variables (S y W) que son dependientes entre sí (Moore et al., 1991). La temperatura del aire se registró cada 30 minutos, en abrigo meteorológico a 1,50 m de altura, mediante un data logger. Para determinar el tiempo térmico se utilizó la sumatoria de los grados días (GD) acumulados durante el periodo de evaluación de las pasturas comenzando el 01 octubre de la siguiente forma:
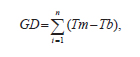
donde: Tm = temperatura media diaria,
Tb = temperatura base (10 °C; Sanderson y Moore, 1999; Mitchell et al., 2001),
n = número de días desde el inicio de las mediciones.
La temperatura media y la lluvia total del mes de octubre superó en 1,6 °C y triplicó, respectivamente, a las medias históricas (1975-2013). En tanto que las temperaturas medias mensuales de noviembre a febrero fueron cercanas a la media histórica, con diferencias inferiores al grado centígrado. En cuanto a la lluvia acumulada, durante noviembre y diciembre fue 2,5 veces inferior a la histórica; sin embargo, el elevado nivel de lluvias registrado al inicio de la primavera pudo haber atenuado, en parte, el posible efecto de la deficiencia posterior. A su vez, la lluvia total del periodo de evaluación fue superior al total histórico (484 vs. 426 mm; figura 1). Esta variabilidad intra e interanual en las lluvias es una característica de la región pampeana (Yahdjian y Sala, 2008), que se acrecentó durante los últimos años (Magrin et al., 2005). Las relaciones entre ambos índices de desarrollo y entre la proporción de lámina viva y los índices se establecieron mediante regresiones lineales. Se probaron los supuestos (ej., normalidad, homocedasticidad e independencia de los residuos) involucrados en la construcción de los modelos. Las pendientes e interceptos de las regresiones lineales fueron comparados mediante el uso de variables dummy. Para estimar el error asociado con la predicción se utilizó el error estándar de la regresión (ej., la raíz cuadrada del cuadrado medio del error (RCME)). La variación de los índices cuantitativos de desarrollo morfológico, a través del tiempo térmico, de ambas especies se analizó mediante la aplicación de modelos lineales mixtos para mediciones repetidas en el tiempo. Todos los análisis fueron realizados con el programa estadístico InfoStat (Di Rienzo et al., 2015).
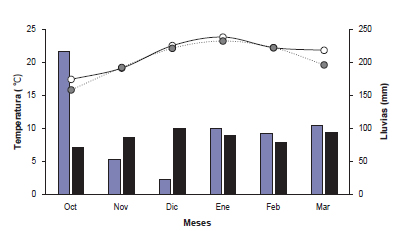
Figura 1. Temperatura media mensual ( ), temperatura media mensual histórica (1975-2013; ), lluvia mensual ( ) y lluvia mensual histórica ( ) desde octubre a febrero.
RESULTADOS Y DISCUSIÓN
La relación existente entre EMP y EMC fue estrecha y positiva (figura 2), lo cual indica que el cambio en uno de ellos se corresponde con un cambio proporcional en magnitud y en la misma dirección en el otro. Estos resultados acuerdan con antecedentes que prueban que EMP puede ser predicho a partir de EMC (Mueller y Fick, 1989; Mitchell et al., 1997). Ello es de utilidad dado que la aplicación del índice EMC elimina la necesidad del secado de los macollos como parte del proceso de determinación del estado medio de desarrollo. Además, se observó que en la medida que incrementó EMC, el incremento en EMP fue menor (p<0,05) en digitaria que en mijo perenne (figura 2). Esto sugiere que los macollos elongados de digitaria fueron más livianos que los de mijo perenne incidiendo sobre EMP y no en EMC dado que, en este último, todos los macollos tienen igual incidencia sin importar su peso. En la figura 3 se puede observar que el desarrollo aparente de las plantas estimado mediante el índice EMP fue más rápido o, en otros términos, el rango de valores obtenidos en función de los GD acumulados fue más amplio para EMP que para EMC. Este resultado es consistente con evaluaciones realizadas en alfalfa (Kalu y Fick, 1981; Kratchunov y Naydenov, 1995) y gramíneas C4 perennes (Hendrickson et al., 1998; Mitchell et al., 1997). En consecuencia, el uso de EMP facilitaría la detección de diferencias entre los estados de desarrollo en mayor medida que EMC (Kalu y Fick, 1981).
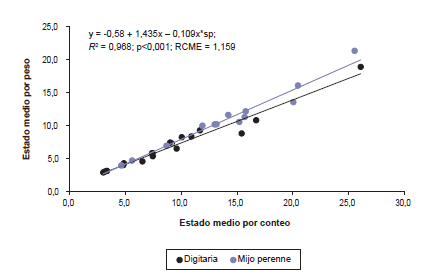
Figura 2. Relaciones entre el estado medio por peso (EMP) y el estado medio por conteo (EMC) para digitaria y mijo perenne (sp: digitaria = 0; sp: mijo perenne = 1). Cada observación representa el promedio de tres repeticiones. RCME, raíz cuadrada del cuadrado medio del error.
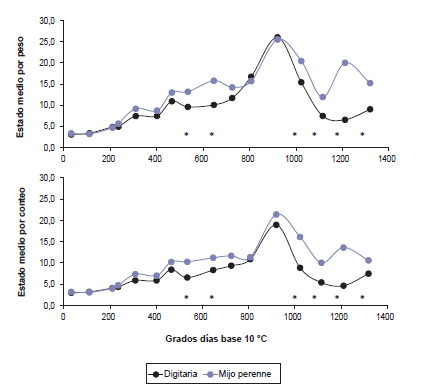
Figura 3. Relaciones entre los estados medio por peso (EMP) y por conteo (EMC) y los grados días base 10 °C acumulados desde el 01 de octubre para (a) digitaria y (b) mijo perenne
Las especies evaluadas presentaron un desarrollo morfológico medio similar (p>0,05), estimado con ambos índices, hasta aproximadamente los 530 GD en que mijo perenne alcanzó el estado medio de elongación de entrenudos (≥11; figura 3). A partir de dicho momento y hasta los 650 GD, ambos índices fueron menores (p<0,05) en digitaria, con respecto a mijo perenne, lo que indicaría que el periodo en que la primera especie permaneció en estado vegetativo fue más prolongado, alrededor de 250 GD (EMP y EMC < 11; digitaria: 646 y 807 GD; mijo perenne: 401 y 534 GD, respectivamente; figura 3). A los 900 GD, ambas especies, se aproximaron o alcanzaron el estado medio reproductivo. Las diferencias observadas en la tasa de desarrollo de las especies durante la fase de elongación de entrenudos sugieren una respuesta diferencial a la temperatura, que no se manifiesta en la expresión del desarrollo reproductivo, dado que ocurrió a un tiempo térmico similar. Esto último podría indicar la presencia de otros factores abióticos involucrados en definir la iniciación floral como la disponibilidad de agua edáfica (Link et al., 1990), el fotoperiodo (Pritchard y De Lacy, 1974) o la edad de los macollos (Loch et al., 2004). Posteriormente, luego de superados los 900 y hasta 1200 GD en digitaria y 1100 GD en mijo perenne, el estado medio numérico decreció (figura 3), lo cual se explicaría por la generación de nuevos macollos (Kalu y Fick, 1981), que se refleja en el incremento en la proporción de macollos vegetativos de 0,44 a 0,83 en digitaria y de 0,04 a 0,28 en mijo perenne. Este incremento, que surge con posterioridad al desarrollo floral, podría ser explicado por la recuperación en la humedad edáfica luego de un periodo de sequía (figura 1), lo que habría determinado un incremento en la tasa de macollaje (Horst y Nelson, 1979; Korte y Chu, 1983). A su vez, el decrecimiento mencionado en los valores de ambos índices fue mayor (p<0,05) en digitaria. Esto se corresponde con la mayor proporción de macollos en estado vegetativo para dicha especie en relación con mijo perenne. Por último, digitaria se mantuvo en estado de desarrollo vegetativo y mijo reinició la elongación de entrenudos, reiterándose la presencia de un periodo en estado medio vegetativo más prolongado en la primera especie.
La elongación anticipada de entrenudos en mijo perenne tendría implicancias respecto del manejo de la defoliación, dado que la elevación de los meristemas apicales aumentaría la probabilidad de su remoción, con efectos negativos sobre el rebrote posterior. Además, se podría esperar una disminución más rápida en la calidad nutritiva de mijo perenne, al alcanzar antes el estado de elongación de entrenudos. En este sentido, Sanderson et al. (1999) observaron, en un estudio de dos años en dos tipos de suelo, que digitaria presentó porcentajes de fibra en detergente neutro inferiores a mijo perenne.
En los primeros seis cortes del presente trabajo los valores del desvío estándar del EMC fueron pequeños en términos relativos, y similares entre especies (figura 4). Ello es indicio de que la población de macollos estaba integrada por individuos con un estado de desarrollo similar; en este caso, en ambas especies solo se registraron macollos en estado vegetativo. Luego, a medida que progresó el desarrollo morfológico y hasta alcanzar los 900 GD el desvío estándar fue en incremento, en respuesta a una diversidad creciente de estados y subestados. Se puede observar que dicho incremento fue menor en digitaria en comparación con mijo perenne (figura 4), lo cual se podría relacionar con una duración más prolongada del periodo vegetativo en la primera especie. Sin embargo, al alcanzar el estado medio reproductivo el desvío estándar de digitaria fue marcadamente superior, indicando una variación mayor en el estado de madurez de los macollos que mijo perenne. Lo anterior pone de manifiesto que, aun cuando dos poblaciones de macollos alcancen a un mismo momento un similar EMC, su composición morfológica puede ser muy diferente. Superado los 900 GD el valor del desvío estándar disminuyó, hasta alcanzar un mínimo a los 1200 y 1100 GD para digitaria y mijo perenne, respectivamente (figura 4). Esto sería consecuencia de, por un lado, la presencia de una gran proporción de macollos reproductivos en un estado de desarrollo avanzado y similar (ej., estado de desarrollo de semillas y madurez: S > 32) y, por el otro, la aparición de nuevos macollos en la población con similar estado de desarrollo. Lo expuesto explicaría la menor heterogeneidad en el estado de desarrollo medio de la población de macollos en ambas especies. Posteriormente, el desvío estándar incrementó en la medida que progresó el desarrollo morfológico de los macollos.
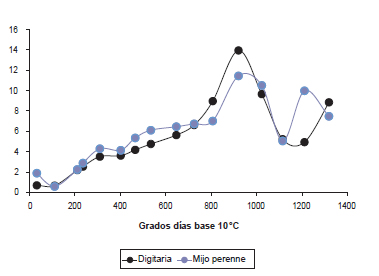
Figura 4. Desvío estándar del estado medio por conteo en función de los grados días base 10 °C acumulados desde el 01 de octubre para digitaria y mijo perenne.
Además, se analizó la dinámica de la partición de la materia seca (ej., proporción de lámina viva) asociada al estado de desarrollo de las dos especies, dado su efecto sobre el valor nutritivo del forraje, al incrementar el contenido de fibra en detergente neutro y disminuir su digestibilidad (Sanderson y Wedin, 1989; Temu et al., 2014). La variación en los índices de desarrollo explicó entre el 83 y 89% de la variación en la proporción de lámina viva, con una dispersión baja a moderada de los datos (RCME entre 4,8 y 5,8 g de lámina cada 100 g de materia seca). A su vez, las ecuaciones de regresión generadas a partir de EMP, en ambas especies, predijeron en forma ligeramente más ajustada la proporción de lámina viva, con relación a EMC (figura 5). Por una parte, la diferencia en la capacidad de predicción de los índices se debería a que en la estimación del índice EMP se considera el peso de los macollos individuales (Kalu y Fick, 1981), mientras que EMC es un promedio de todos los macollos presentes, teniendo los macollos jóvenes y maduros igual ponderación. El índice EMP incorpora el efecto ejercido por los tallos maduros, más pesados que los jóvenes, determinando así una mayor sensibilidad para captar cambios en el desarrollo morfológico, lo que permitió predecir la proporción de lámina en forma más ajustada. En el mismo sentido, varios autores encontraron una mejor capacidad de predicción de las variaciones en el valor nutricional del forraje mediante EMP (Kalu y Fick, 1981; Mitchell et al., 2001). En tanto que, bajo condiciones de pastoreo, EMC fue un pobre predictor de la relación lámina:tallo con el avance en la estación de crecimiento (Smart et al., 2001).
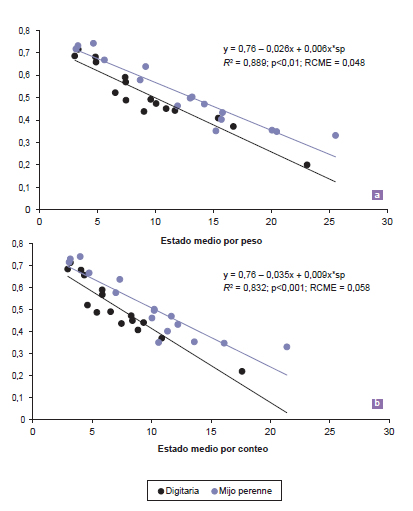
Figura 5. Relaciones entre la proporción de lámina y (a) el estado medio por peso (EMP) y (b) estado medio por conteo (EMC) para digitaria y mijo perenne (sp: digitaria = 0; sp: mijo perenne = 1). RCME, raíz cuadrada del cuadrado medio del error.
Por otra parte, la tasa de disminución en la proporción de lámina viva en función de ambos índices fue mayor (p<0,05) en digitaria con respecto a mijo perenne (figura 5), lo que se explicaría por el mayor flujo de senescencia en la primera (20,4 vs. 7,1 mg de material senescente por macollo; p<0,001; EE: 0,36) a través de todos los cortes (interacción especie x corte; p>0,227). Esto sugiere la importancia del flujo de senescencia como determinante de la menor proporción de lámina viva en digitaria, aun cuando el inicio de la fase de elongación de entrenudos en esta especie fue más tardía que para mijo perenne. Los resultados del presente trabajo sugieren que sería posible predecir la proporción de lámina mediante estos índices. Sin embargo, se requeriría ampliar las evaluaciones para proveer evidencia acerca de la posibilidad de extrapolar esta información a otras condiciones ambientales. Además, sería necesario evaluar posibles diferencias estacionales en la proporción de lámina, en los distintos estados de madurez de la pastura. Si bien a medida que las plantas forrajeras maduran la materia seca acumulada aumenta y su valor nutricional disminuye (Mitchell et al., 2001; Ziehr et al., 2014), estas tendencias podrían ser modificadas por las condiciones ambientales. Es decir, la disminución observada en la proporción de lámina y en la concentración proteica (Ferri, 2011), con el avance de la madurez, es afectada por las condiciones ambientales dado que estas determinan la cantidad de materia seca acumulada hasta alcanzar cada estado de desarrollo. De todos modos, aun cuando a través del desarrollo morfológico se pueda predecir la calidad nutricional del forraje bajo condiciones ambientales variables, la principal utilidad de los índices (EMP y EMC) radicaría en lograr caracterizaciones precisas y cuantitativas, del estado de desarrollo morfológico de especies C4 perennes.
CONCLUSIONES
Los índices cuantitativos por peso y por conteo permitieron describir el estado de desarrollo morfológico medio de digitaria y mijo perenne, aunque, el primero fue más sensible para detectar cambios morfológicos. A través de la aplicación de ambos índices se encontraron diferencias entre especies en cuanto a la duración del estado vegetativo; a su vez, ambas especies alcanzaron el estado reproductivo en forma simultánea. Además, dichos índices podrían proveer una estimación de la proporción de lámina viva de la población de macollos de ambas especies. Sin embargo, se requiere probar la consistencia de estas relaciones ante variaciones en las condiciones ambientales y a través de la estación de crecimiento.
BIBLIOGRAFÍA
1. BRISKE, D.D. 1991. Developmental morphology and physiology of grasses. Timber Press, Portland, OR. En: HEITSCHMIDT, R.K.; STUTH, J.W. (Eds.). Grazing management: An ecological perspective, pp. 85-108. [ Links ]
2. BUXTON, D.R.; MARTEN, G.C. 1989. Forage quality of plant parts of perennial grasses and relationship to phenology. Crop Sci. 29, 429-435. [ Links ]
3. COLEMAN, S.W.; MOORE, J.E.; WILSON, J.R. 2004. Quality and utilization. En: MOSER, L.E; BURSON, B.L.; SOLLENBERGER, L.E. (Eds.). Warm-season (C4) grasses. Agronomy Series 45, ASA-CSSA-SSSA, Madison, WI, EUA, pp. 267-308. [ Links ]
4. DI RIENZO, J.A.; CASANOVES, F.; BALZARINI, M.G., GONZÁLEZ, L., TABLADA, M.; ROBLEDO, C.W. 2015. InfoStat, versión 2015, Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. [ Links ]
5. FERRI, C.M. 2011. Relaciones entre composición química y estructura en Panicum coloratum L. Rev. Arg. Prod. Anim. 31, 504. [ Links ]
6. FERRI, C.M. 2014. Gramíneas forrajeras perennes de crecimiento estival (C4) para la región Pampeana semiárida, en el contexto de la intensificación ganadera y del cambio climático. Resultados finales. Proyectos de investigación científica y tecnológica orientados al desarrollo productivo provincial. Universidad Nacional de La Pampa, EdUNLPam, Santa Rosa, pp. 92-145. [ Links ]
7. FICK, G.; JANSON, C.G. 1990. Testing mean stage as a predictor of alfalfa forage quality with growth chamber trials. Crop. Sci. 30, 678-682. [ Links ]
8. GOODALL, D.W. 1952. Some considerations in the use of point quadrats for the analysis of vegetation. iv Patterns and minimal area. Aust. J. Sci. Res. 5, 1-41. [ Links ]
9. GUSTAVSSON, A.M. 2011. A developmental scale for perennial forage grasses based on the decimal code framework. Grass Forage Sci. 66, 93-108. [ Links ]
10. HENDRICKSON, J.R.; MOSER, L.E.; MOORE, K.J.; WALLER, S.S. 1998. Morphological development of 2 warm-season grasses the Nebraska Sandhills. J. Range Manage. 51, 456-462. [ Links ]
11. HORST, G.L.; NELSON, C.J. 1979. Compensatory growth of tall fescue following drought. Agron. J. 71, 559-563. [ Links ]
12. KALU, B.A.; FICK, G.W. 1981. Quantifying Morphological stage of maturity as a predictor of alfalfa herbage quality. Crop Sci. 23, 267-271. [ Links ]
13. KORTE, C.J.; CHU, A.C.P. 1983. Some effects of drought on perennial ryegrass swards. Proc. N.Z. Grassl. Ass. 44, 211-216. [ Links ]
14. KRATCHUNOV, I.; NAYDENOV, T. 1995. Estimation of Lucerne forage quality by means of morphological and meteorological data. Eur. J. Agron. 4, 263-267. [ Links ]
15. LINK, S.O.; GEE, G.W.; DOWNS, J.L. 1990. The effect of water stress on phenological and ecophysiological characteristics of cheatgrass and Sandberg`s bluegrass. J. Range Manage. 43, 506-513. [ Links ]
16. LOCH, D.S.; ADKINS, S.W.; HESLEHURST, M.R.; PATERSON, M.E.; BELLAIRS, S.M. 2004. Seed formation, development, and germination. En: MOSER, L.E; BURSON, B.L.; SOLLENBERGER, L.E. (Eds.). Warm-season (C4) grasses Agronomy Series 45, ASACSSA- SSSA, Madison, WI, EUA., pp. 95-143. [ Links ]
17. MAGRIN, G.O.; TAVASSO, M.I.; RODRÍGUEZ, G.R. 2005. Changes in climate and crop production during the 20th century in Argentina. Clim. Chang. 72, 229-249. [ Links ]
18. MITCHEL, R.B.; MOSER, K.P.; MOORE, K.J. 1997. Relationships of visual and quantitative methods of grass sward development. Proc. 18th Int. Grassl. Congr., Winnipeg, Manitoba, Canadá, p. 7-8. [ Links ]
19. MITCHELL, R.; FRITZ, J.; MOORE, K.; MOSER, L.; VOGEL, K.; REDFEARN, D.; WESTER, D. 2001. Predicting forage quality in switchgrass and big bluestem. Agron. J. 93, 118-124. [ Links ]
20. MOORE, K.J.; MOSER, L.E.; VOGEL, K.P.; WALLIER, S.S.; JONSON, B.E.; PEDERSEN, J.F. 1991. Describing and quantifying growth stages of perennial for age grasses. Agron. J. 83, 1073-1077. [ Links ]
21. MOORE, K.J.; MOSER, K.P. 1995. Quantifying developmental morphology of perennial grasses. Crop Sci. 35, 37-43. [ Links ]
22. MUELLER, S.C.; FICK, G.W. 1989. Converting alfalfa development measurements from mean stage by count to mean stage by weight. Crop Sci. 29, 821-823. [ Links ]
23. NADA, Y. 1980. Photoperiodic responses in flowering of main tropical pastures grasses. J. Jpn. Soc. Grassl. Sci. 26, 157-164. [ Links ]
24. NELSON, C.J.; MOSER, L.E. 1994. Plant factor affecting forage quality. En: Fahey, G.C. (Ed.). Forage quality, evaluation and utilization. ASA, CSSA and SSSA. Madison, WI, USA, pp. 409-420. [ Links ]
25. PETRUZZI, H.J.; JOUVE, V.V.; FERRI C.M.; STRITZLER, N.P.; PAGELLA, J.H. 1996. Tasa de crecimiento y valor nutritivo de siete gramíneas estivales en la región Pampeana semiárida. Rev. Arg. Prod. Anim. 16, 175-176. [ Links ]
26. PRITCHARD, A.J.; DE LACY, I.H. 1974. The cytology, breeding system and flowering behavior of Panicum coloratum. Aust. J. Bot. 22, 57-66. [ Links ]
27. ROBERTO, Z.; ADEMA, E.; RUCCI, T. 2005. Relevamiento fisonómico de la vegetación en el área del caldenal. Publicación Técnico N.º 60, EEA Anguil, INTA, p 24. [ Links ]
28. SANDERSON, M.A. 1992. Morphological development of Switchgrass and Kleingrass. Agron. J. 84, 415-419. [ Links ]
29. SANDERSON, M.A.; MOORE, K.J. 1999. Switchgrass morphological development predicted from day of the year or degree day models. Agron. J. 91, 732-734. [ Links ]
30. SANDERSON, M.A.; VOIGT, P.; JONES, R.M. 1999. Yield and quality of warm-season grasses in central Texas. J. Range Manage. 52, 145-150. [ Links ]
31. SANDERSON, M.; WEDIN, W. 1989. Phenological stage and herbage quality relationship in temperature grasses and legumes. Agron. J. 81, 864-869. [ Links ]
32. SAyDS (Secretaria de Ambiente y Desarrollo Sustentable). 2006. Primer Inventario Nacional de Bosques Nativos. Inventario de campo de la región del espinal distritos Caldén y Ñandubay. Anexo 1. Estado de conservación del distrito Caldén, p. 77. [ Links ]
33. SMART, A.J.; SCHACHT, W.H.; MOSER, L.E. 2001. Predicting leaf/stem ratio and nutritive value in grazed and nongrazed bigbluestem. Agron. J. 93, 1243-1249. [ Links ]
34. STRITZLER, N.P.; PAGELLA, J.H., JOUVE, V.V.; FERRI, C.M. 1996. Semi-arid warm-season grass yield and nutritive value in Argentina. J. Range Manage. 49, 121−125.
35. TEMU, V.W.; RUDE, B.J.; BALDWIN, B.S. 2014. Nutritive value response of native warm-season forage grasses to harvest intervals and durations in mixed stands. Plants 3, 266-283. [ Links ]
36. VIGLIZZO, E.F.; CARREÑO, L.V; PEREYRA, H.; RICARD, F.; CLATT, J.; PINCÉN, D. 2010. Dinámica de la frontera agropecuaria y cambio tecnológico. En: VIGLIZZO, E.F.; JOBBÁGY, E.C. (Eds.). Expansión de la frontera agropecuaria en Argentina y su impacto ecológico-ambiental. INTA, Buenos Aires, pp. 9-16. [ Links ]
37. YAHDJIAN, L.; SALAS, O.E. 2008. Climate change impacts on South American Rangelands. Rangelands 30, 34-39. [ Links ]
38. ZIEHR, R.D.; REA, G.L.; DOUGLAS, J.L.; SPAETH, K.E.; PEACOCK, G.L.; MUIR, J.P. 2014. Ontogenesis and nutritive value of warm-season perennial bunch grasses. Trop. Grassl. 2, 188-196. [ Links ]

