











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkINTRODUCTION
Keywords: alfalfa, high cutting frequency, establishment year, contrasting dormancy level.
The establishment phase of lucerne (Medicago sativa L.), after sowing, refers to a period in which biomass is mainly partitioned to the perennial organs which consist of the taproot plus crown. This phase ends, regardless of ontogeny, when the pasture reaches ∼4-5 t DM/ha of perennial biomass (Sim et al., 2015). During the establishment phase, the perennial organs are a strong sink for the carbon fixed by the plant (Khaiti and Lemaire, 1992). Moreover, until perennial biomass do not exceed 3000-4000 kg DM/ha, the partitioning of photosynthate to these organs is high and hence the aerial growth is below their potential (Sim et al., 2015). Consequently, management practices that favor the rapid formation of perennial organs lead to rapidly obtaining a highly productive lucerne pasture. Therefore, the current recommendation is to delay first defoliation until open flower buds are observed during the first year after sowing (Moot et al., 2003). This long period of uninterrupted growth allows maximum opportunity to establish reserves of carbon and nitrogen in the perennial organs (Justes et al., 2002; Thiebeau et al., 2011).
The previous recommendation contrasts with the new grazing management proposal which advises that the start of grazing in spring should be when the standing biomass (above 5 cm) of the first paddock is close to 1,000-1,500 kg DM/ha (15-20 cm of canopy height) and then paddocks should be rotated every 15-25 days, with a 30-40 cm of pre-grazing canopy height, which optimizes forage production and forage quality (Berone et al., 2021). Such a recommendation results in 30% higher beef live weight production (Berone et al., 2020), compared with the traditional criteria of starting grazing at 10% flowering and rotation every 30-35 days (Basigalup and Ustarroz, 2007).
A high frequency and severity of defoliation can be very detrimental to the persistence of lucerne plants during their establishment phase, as it leads to an increased reliance on C and N reserves stored in perennial organs for regrowth (Volenec et al., 1996), which are minimal during this stage (Sim et al., 2015). Previous work shows that plants less severely defoliated, with higher residual green leaf area, have less dependence of root C and N reserves to regrowth after grazing (Simon et al., 2004; Meuriot et al., 2005). Therefore, frequent but less severe defoliation would be a viable option for grazing during the first spring after sowing.
The implication of reconciling a rapid formation of a vigorous perennial system with an early and intensive grazing of lucerne is not a minor issue. In fact, Argentine production systems lucerne pastures in the establishment phase (i.e. one year after sowing) represent a high proportion of the area sowed with perennial pastures (Gastaldi et al., 2020; Jauregui et al., 2022). Consequently, management practices allowing grazing lucerne pastures as early as possible, without affecting their productive persistence, should have a great impact at the system level.
The aim of the present work was to evaluate, during the first year after sowing, the impact of cutting severity on aerial growth and perennial biomass, in two lucerne cultivars contrasting in their degree of winter dormancy and defoliated at a frequency higher than that usually recommended (i.e. defoliations earlier than the 10% flowering stage).
MATERIALS AND METHODS
Site, materials tested and experimental design
The experiment was carried out in a glasshouse and outdoors in Balcarce, Argentina (37° 45’ S, 58° 18’ W), under natural photoperiod from July 2014 to September 2015. The semi-dormant WL611 (G6) and the non-dormant WL2058 (FD10) cultivars and two cutting heights (4 and 10 cm) were studied.
On 22 July 2014, two hundred and sixty-four polyvinyl chloride (PVC) containers (132 for G6 and 132 for G10) 78 cm in height and 16 cm in diameter, lined with polystyrene bag perforated at the bottom were filled with a uniform mixture of topsoil from a typical Argiudol (6% OM; 33 ppm P Bray 1) and river sand (1:1 v/v). Prior to sowing, all containers were irrigated to saturation and afterwards (during all the experiment) were watered every two days with tap water in order to maintain soil water content close to field capacity. Sowing was carried out in a greenhouse on 24 July 2014. Ten seeds were placed in the centerline of the containers and a first thinning was carried out when the seedlings reached the stage of a fully unfolded true leaf, leaving only three plants per container.
On September 24, 2014, the plants were defoliated at 4 cm in height and the containers were placed outdoors. For this, a distance of approximately 20 cm was maintained between the center of one tube and the center of another tube. The 132 containers of each cultivar were placed in two (2.0 m x 1.5 m) separated by a corridor (1.0 m wide and 2.0 m long). Once installed, the cutting height treatments were assigned in a completely randomized design with three replicates (i.e. three containers). In total, the entire experiment occupied an area of 8 m2 (4.0 m x 2.0 m). On October 3,2014, a second thinning of plants was carried out, leaving a final density of two plants per container. One hundred and eight containers were used for data collection in destructive sampling while the remaining were used as a borders. The arrangement of the containers described above allowed simulating a pasture condition with a density of 500,000 plants/ha (Brown et al., 2006). To avoid nitrogen (N) deficiencies, urea fertilization was applied weekly at a rate of 46 kg N/ha (100 kg urea/ha). To avoid phosphorus (P) deficiencies, triple superphosphate was applied after each cutting at a dose of 20 kg P/ha. The experiment was kept free of weeds and pests.
Meteorological record
Data on incident radiation and mean daily temperature were obtained from the Agrometeorological Station of the EEA INTA Balcarce, located less than 500 m from the experiment. The accumulated rain during the outdoor period (October 2014-September 2015) was quite similar (915 mm) to that observed in the 2004-2014 cycle (879 mm). The monthly mean daily incident radiation was similar (14.0 Mj) to that observed in the 2004-2014 cycle, and fluctuated between 5.9-6.4 Mj for July-August 2015 and 22.9-22.3 Mj for December 2014 to January 2015. The monthly mean daily temperature was higher than that observed in the 2004-2014 cycle (on average +0.7 °C), and fluctuated between 9-11 °C for the coldest months (July-August 2015) and 21-22 °C for the warmest months (December 2014 to February 2015).
Measurements
From November 5, 2014 to September 2, 2015, ten harvests were made with an average frequency of 370 cumulative growing degree days (GDD), considering a base temperature of 5 °C (Thiébeau et al., 2011; table 1). Destructively sampled containers were sequentially removed from the southern side. After each sampling date, the southern border was moved northwards after each destructive sample to minimize edge effects and to preserve the integrity of the experiment. On February 25, 2015, after the corresponding measurements, all plants were cut at 4 cm, to remove the accumulated dead material, which was very high, particularly in the plants cut at 10 cm (data not shown). The following harvests were carried out at the corresponding cutting heights. On April 10, 2015 it was necessary to make an unplanned cut, due to a widespread pigeon attack that was controlled late. The cutting height of treatments was conserved, but no determinations were made.
After the cut all the aerial biomass was separated from the crown. Subsequently, the crown and the 300 mm upper section of the tap root, which represent around 76% of the total biomass of the root system (data not shown), were carefully washed with tap water and the crown was separated from the root by visually determining the transition zone between both tissue types, which coincides with the location of the last bud (Teixeira et al., 2007). Given that the analyzed treatment showed a similar root proportion in perennial biomass (data not shown), and previous works (Teixeira et al., 2007) show that, results in relation to %N in organs of reserve (root and crown) are not modified if they are analyzed separately or as a whole (i.e. analyzing %N in perennial biomass), the crown plus root was treated as a single variable, the perennial biomass (BPER), similar to Teixeira et al. (2007). The above-ground biomass (AB) was separated into that between the upper level of the crown and the cutting height (ABRES, representing the residual above-ground biomass after a cut) and that above the cutting height (ABHARV, representing the biomass harvested in a cut). Both ABRES and ABHARV were further separated into leaves (ABLEAF, laminae + petioles) and stems (ABSTEM).
In the ABRES, the stems were separated into old/dead (empty and solid to the touch and yellow-brown in color) and young stems, generated during the last regrowth (flexible to the touch, solid and green in color). The number of total nodes per plant in the ABRES (TNRES) was estimated as the product between the number of young stems and the number of nodes per young stem. The TNRES, represents the amount of sinks (axillary buds and internodes) available to produce new aerial biomass during the regrowth after cutting. The sum of green leaves and young stems constitutes the green ABRES (ABRES_green). All fractions were kept in a freezer at -20 °C until drying in a forced air oven at 60 °C for 72 hours. The leaf ratio (PROPLEAF = ABLEAF /AB) was calculated for both ABHARV and ABRES. The total aerial growth ABGROW was estimated through the sum of ABRES_green and ABHARV.
The residual leaf area index (LAIRES, m2 leaf/m2 soil) was determined as the product of the ABRES_green (g DM/m2), the PROPLEAF in the ABRES_green and the specific leaf area of the residual leaves (SLARES, m2 leaf/g DM leaf). To determine the SLARES, prior to oven drying, a sample of leaves was taken from the ABRES, laid out on a flat surface and photographed with a 14.1 Megapixel digital camera (SONY Cyber-shot, Japan). They were then oven-dried at 60°C to constant weight. The photos were processed to obtain the total leaf area per sample using ImageJ software (v. 1.46r; National Institute of Health; USA). The SLARES was calculated as the ratio between the total leaf area of the sample and its dry weight.
The nitrogen concentration (%N) in the leaves (%NLEAF) from the ABRES_green and in the perennial biomass (%NPER) was determined by total combustion of the sample in ultra-pure oxygen atmosphere, using a LECO equipment (LECO-FP528, Michigan, USA). Leaf nitrogen specific to the residual leaf area (SLNRES, g N/m2 leaf) was estimated as the quotient between the N in residual leaf (g N/m2 soil = %NLEAF * ABRES_leaf) and the LAIRES (m2 leaf/m2 soil). The absolute nitrogen content in perennial biomass (NPER, kg N/ha) was estimated as the product between %NPER and the perennial biomass.
Statistical analysis
Treatments were arranged as the factorial combination of two lucerne cultivars (G6 and G10) and two cutting heights (40 cm and 10 cm from soil level) in a completely randomized design with three replicates (i.e. three containers). For all the variables considered, on each date, the average of two plants per container was analyzed by ANOVA using the InfoStat statistical programme (Di Rienzo et al., 2015). When necessary, the means were compared using the least significant difference test with a significance level of 0.05.
RESULTS
Aerial biomass
Regardless of cultivar, the cumulative aerial growth (ABGROW) of plants cut at 10 cm in height was 25% higher (p<0.05) than that of plants cut at 4 cm in height, and no differences were detected between cultivars (table 2). Concerning to the accumulated ABHARV, no differences (p<0.05) were detected between treatments (table 2). Irrespective of treatments, approximately 87% of the aboveground biomass was produced during spring-summer, while the remaining 13% was produced in autumn-winter (data not shown).
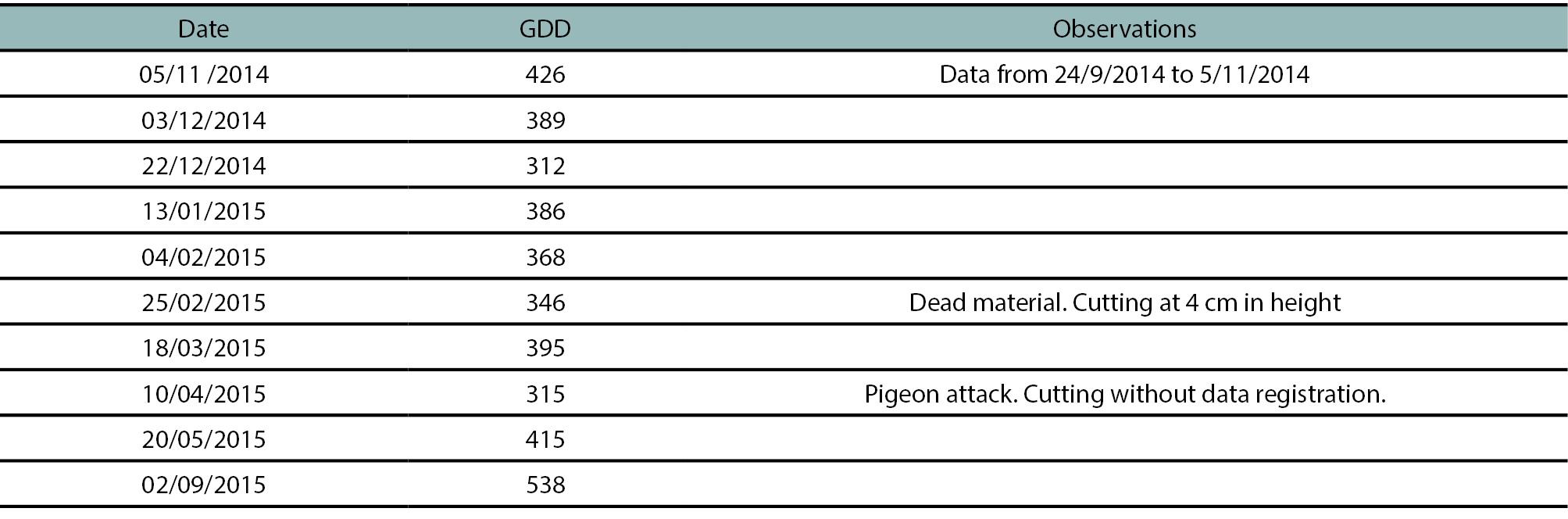
The total residual biomass (ABRES) increased over time and was higher (p < 0.05) in plants cut at 10 cm in height (figure 1a). The residual green biomass (ABRES_green) was mostly composed of green stem (~80-90%, data not shown) and plants cut at 10 cm showed significantly higher ABRES_green than those cut at 4 cm (figure 1b). The differences between cultivars were less common and were observed in some autumn-winter dates, in plants cut at 10 cm in height, when cultivar G6 showed higher values than G10 (figures 1a, b). The dead material in the residual biomass increased strongly towards mid-summer, particularly in plants cut at 10 cm in height (data not shown, but easily deducted from the difference between the data in figures 1a and 1b).
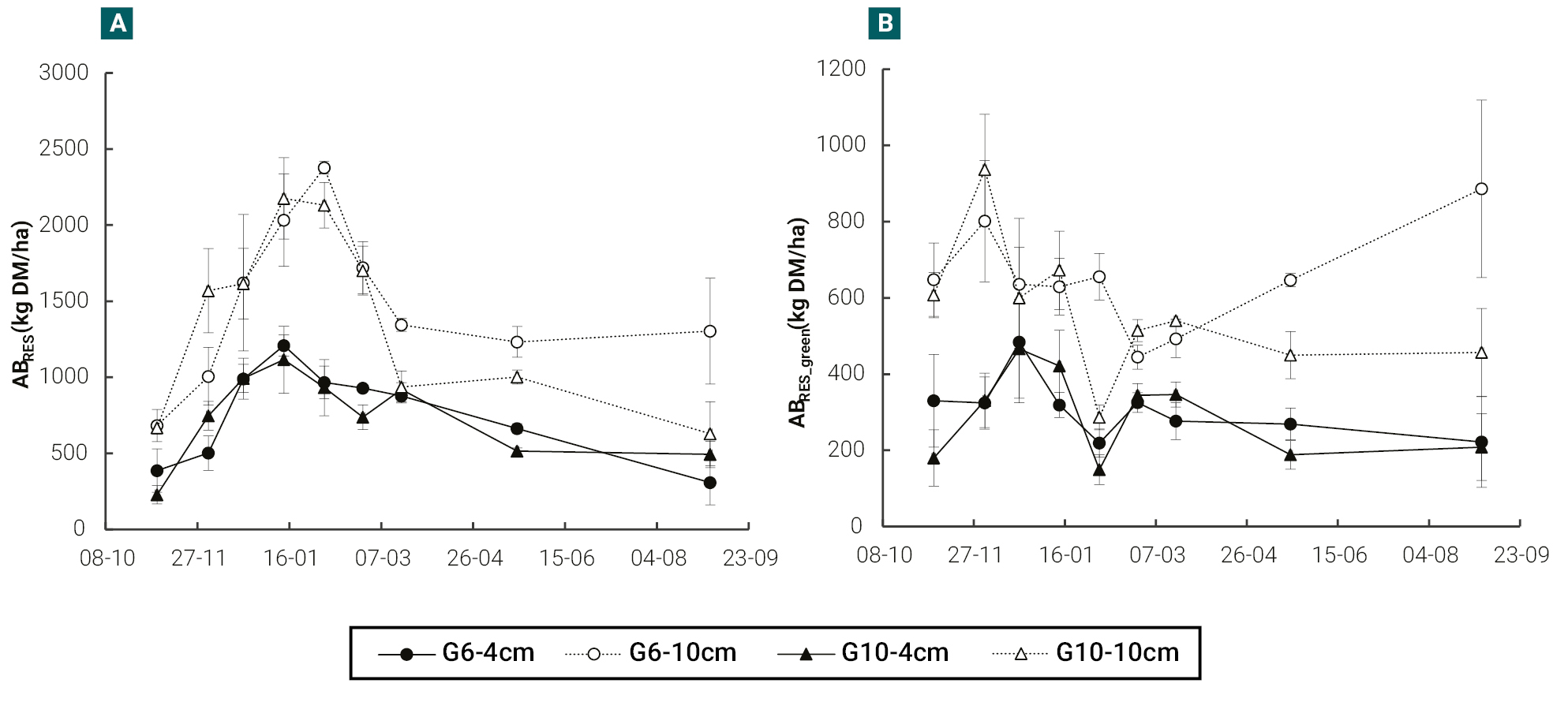
Figure 1.Evolution of (a) total residual biomass (ABRES) and (b) green residual biomass (ABRES_green), in a semi-dormant cultivar (G6) and a non-dormant (G10) cultivar of lucerne, cut at 4 and 10 cm in height with an average frequency of 370 cumulative degree days (base temperature 5 °C). Vertical bars indicate ± one standard error of the mean. In each date, means were compared using the least significant difference test with a significance level of 0.05.
The residual leaf area index (LAIRES) showed a similar behavior to that observed in the ABRES_green. Plants cut at 10 cm showed higher LAIRES (p<0.05) than those cut at 4 cm at all cutting dates (figure 2a), with differences between cultivars being less common. Except for two measurements at the end of spring, the cultivar G6 cut at 10 cm in height (G6-10 cm) showed, generally, higher LAIRES than the rest of the treatments from January onwards (figure 2a).
Regarding the number of total nodes per plant in the remnant (TNRES), the highest values (p<0.05) were observed in cultivar G6 with respect to G10 and in plants cut at 10 cm with respect to those cut at 4 cm (figure 2b). The cultivar G6 severely defoliated (G6-4cm) showed quite similar values of TNRES to those of cultivar G10 lightly defoliated (G10-10cm) and was even higher from March onwards (figure 2b).
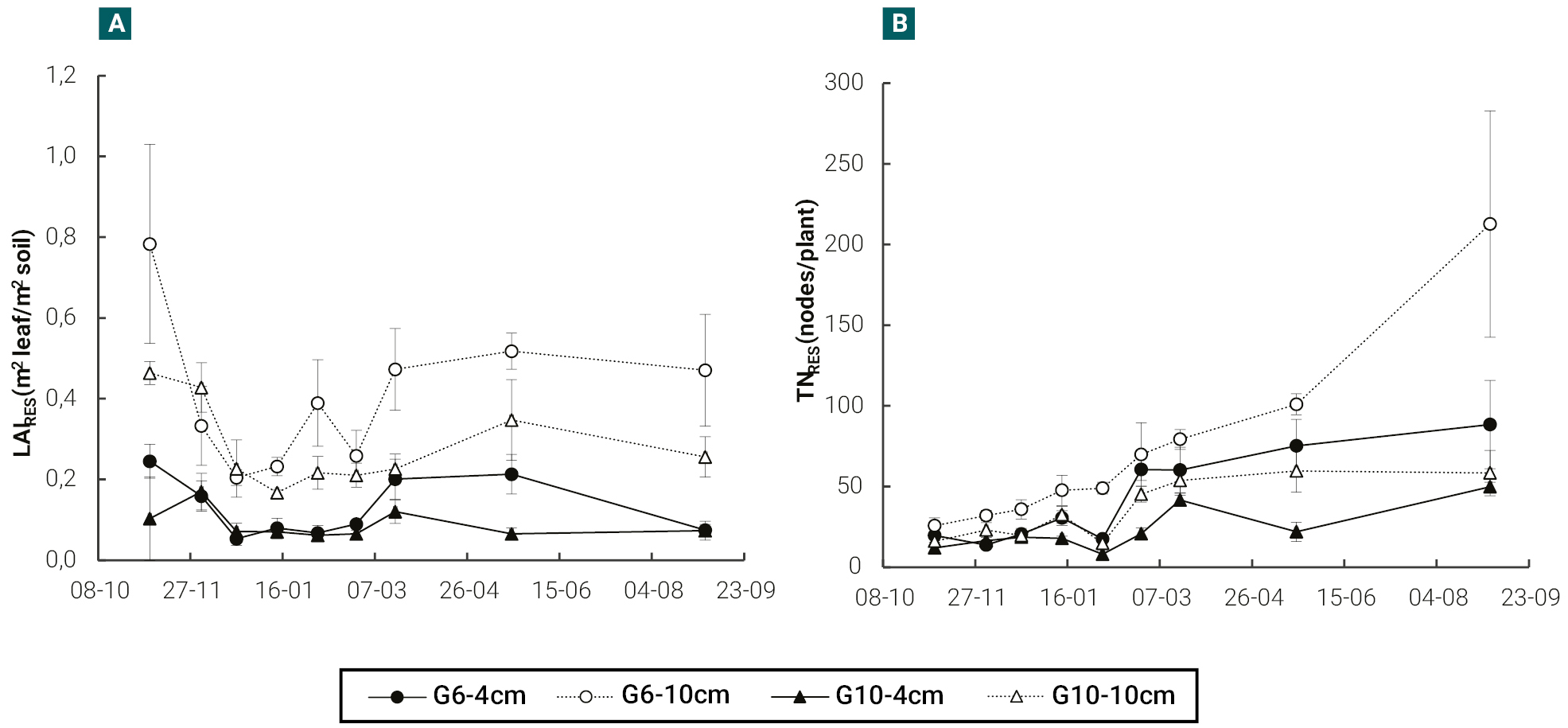
Figure 2Evolution of the residual leaf area index (LAIRES) (a) and the number of total nodes per plant in the residual green biomass (TNRES) (b), in a semi-dormant cultivar (G6) and a non-dormant (G10) cultivar of lucerne, cut at 4 and 10 cm in height with an average frequency of 370 cumulative degree days (base temperature 5 °C). Vertical bars indicate ± one standard error of the mean. In each date, means were compared using the least significant difference test with a significance level of 0.05.
In both cultivars, plants cut at 10 cm in height showed, with respect to those cut at 4 cm, higher (p<0.05) absolute N content (NFOL_RES) and specific leaf nitrogen (SLNRES) in the remaining leaf area, the differences between cultivars being practically irrelevant (figures 3a, b).
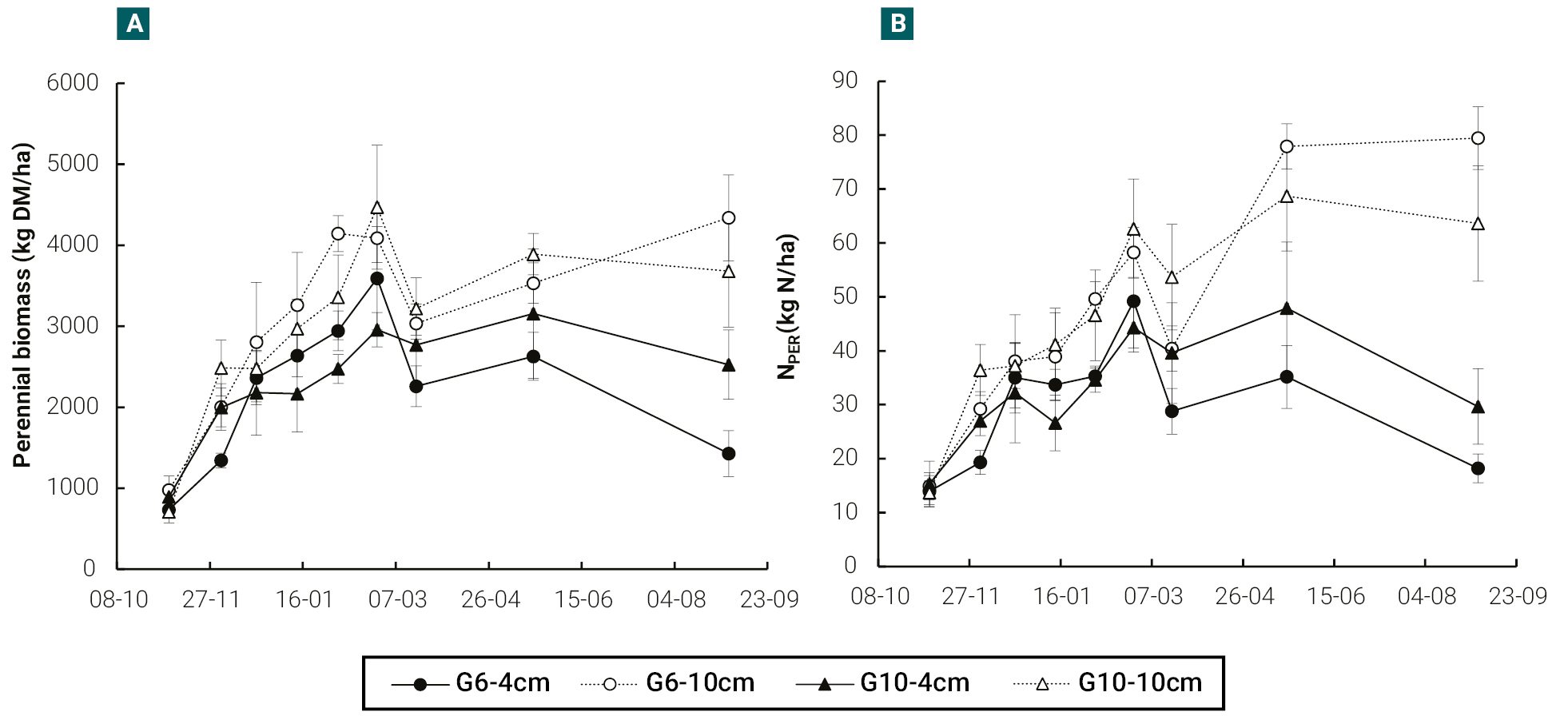
Figure 3.Evolution of absolute N content of residual leaf (NFOL_RES) (a) and specific leaf nitrogen of residual leaf area (SLNRES) (b), in a semi-dormant cultivar (G6) and a non-dormant (G10) cultivar of lucerne, cut at 4 and 10 cm in height with an average frequency of 370 cumulative degree days (base temperature 5 °C). Vertical bars indicate ± one standard error of the mean. In each date, means were compared using the least significant difference test with a significance level of 0.05.
Perennial biomass
In both cultivars, perennial biomass (BPER) increased until mid-late summer and plants cut at 10 cm showed higher BPER values than those cut at 4 cm, especially from mid-summer onwards (figure 4a). The nitrogen content of perennial biomass (NPER) was also superior in 10 cm cutting height, particularly from mid-summer onwards (figure 4b). For these variables, differences between cultivars were practically irrelevant.
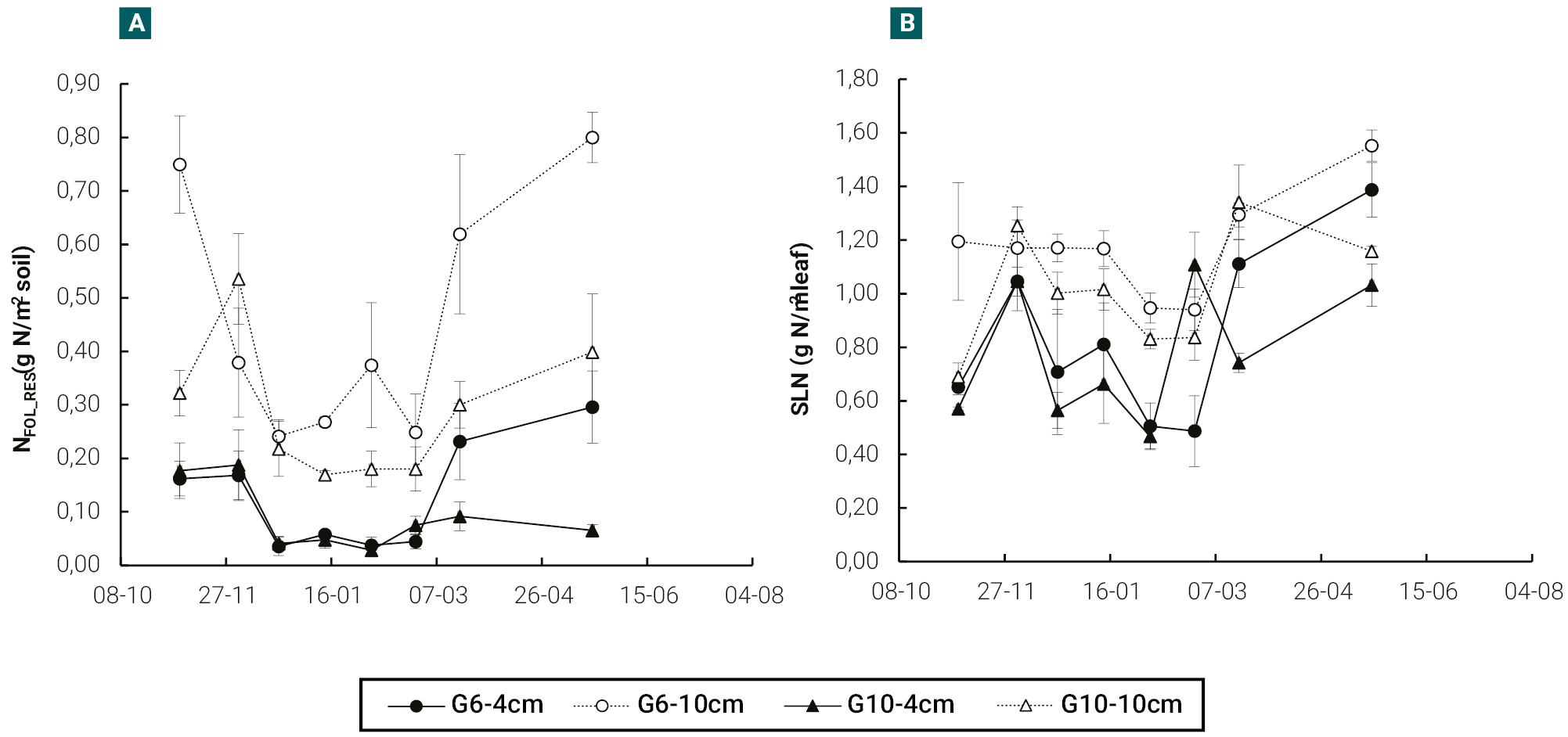
Figure 4.Evolution of perennial biomass (root plus crown) (a) and of the amount of N (NPER) in perennial biomass (b), in a semi-dormant cultivar (G6) and a non-dormant (G10) cultivar of lucerne, cut at 4 and 10 cm in height with an average frequency of 370 cumulative degree days (base temperature 5 °C). Vertical bars indicate ± one standard error of the mean. In each date, means were compared using the least significant difference test with a significance level of 0.05.
DISCUSSION
In this work, the impact of cutting height (4 cm vs. 10 cm) on the performance of two lucerne cultivars, contrasting in their winter dormancy (G6 vs. G10) and subjected to frequent defoliations during the establishment year was analyzed. Due to a general lack of differences between cultivars, which coincides with previous data reported for the region (Pece and Cangiano, 2003), the discussion will focus only on the impact of cutting height.
Aerial biomass
In both cultivars, the accumulated forage production (i.e. the biomass harvested above cutting height, ABCOS) was not affected by the cutting height and the total aerial growth (i.e. biomass produced from ground level, BAGROW) was higher in plants cut at 10 cm than in those cut at 4 cm (table 2). The higher ABGROW in less severely cut plants can be due, in part, to the higher amount of photosynthetically active tissue in the residual post-cutting biomass (figures 1b, 2a), as was previously reported (Langer and Steinke, 1965; Hodgkinson et al., 1972; Simon et al., 2004; Meuriot et al., 2005).
In lucerne, the aerial regrowth is derived from two types of shoots, those originated from the buds of the crown (crown shoots) and those originated from the axillary buds located in the nodes of the remaining stems (axillary shoots) (Goose et al., 1988). Although the origin of shoots was not quantified, the higher TNRES in plants defoliated at 10 cm in height (figure 2b) suggests, for plant defoliated at 10 cm, an important contribution of the axillary shoots in biomass yield. In fact, axillary shoots play an important role in intercepted radiation and dry matter production, due to their rapid emergence after cutting, at a time when the development of leaf area is the main factor limiting growth (Goose et al., 1988). The importance of the contribution of axillary shoots should be higher in frequently defoliated plants (i.e. short-lived regrowth) such as those of the present work. This is confirmed by previous research showing that, in plants severely cut, the slow shoot emergence limits the regrowth rate (Leach 1968; García et al., 2021).
In addition, plants cut at 10 cm in height showed, with respect to plants cut at 4 cm, higher nitrogen content in the residual leaf (figure 3). The higher specific leaf nitrogen in the residual leaf area of plants cut at 10 cm in height (SLNRES, figure 3b) suggests greater photosynthetic capacity than those cut at 4 cm (Teixeira et al., 2008). In addition, the higher nitrogen content in the residual leaf of plants cut at 10 cm in height (NFOL_RES) could be contributed as N source to the initial growth of nearby axillary buds during early phases of shoot regrowth (Dufour et al., 2004), supplementing the N provided by the crown and roots which was also higher in plants cut at 10 cm in height (figure 4b). Therefore, in plants defoliated at a higher frequency (i.e. ~350 GDD) than the usually recommended (i.e. 10% flowering stage), a higher cutting height promote residual leaf area with higher capacity to both, intercept the incoming solar radiation (due to a higher LAIRES and higher TNRES) and to convert the captured radiation in biomass more efficiently (due to a higher NFOL_RES, higher NFERES and higher NPER).
In addition to the higher photosynthetic source (LAIRES and NFERES), another factor contributing to the higher BAGROW in plants less severely defoliated is a higher amount of sinks available to produce new aerial biomass during the regrowth (Leach, 1968; 1969). In fact, the number of total nodes per plant in the young stems (axillary and crown) of the residual was also higher in plants cut at 10 cm in height (TNRES, figure 3b). Based on what was previously discussed, it is possible to conclude that in both cultivars, the higher aerial growth of plants cut at 10 cm can be explained by both, a higher photosynthetic source (i.e. higher LAIRES, higher NFERES) and a higher amount of sinks in the residual biomass (i.e. higher number of TNRES).
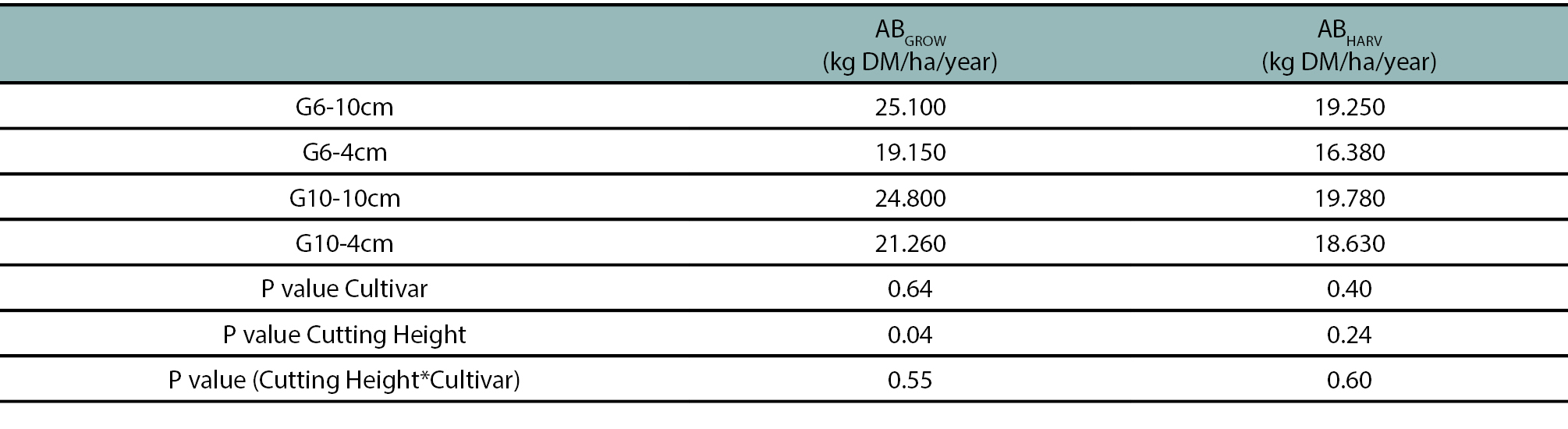
Table 2.Cumulative aboveground biomass produced (ABGROW) and cumulative harvested aboveground biomass (ABHARV) in a semi-dormant cultivar (G6) and a non-dormant (G10) cultivar of lucerne, cut at 4 and 10 cm in height with an average frequency of 370 cumulative degree days (base temperature 5 °C). Means were compared using the least significant difference test with a significance level of 0.05.
Perennial biomass
In both cultivars, defoliated at a frequency higher than that usually recommended for the first year (i.e. defoliations earlier than the 10% flowering stage), the plants cut at 10 cm in height showed higher perennial biomass than those cut at 4 cm. Moreover, plants cut at 10 cm in height quickly achieved, and maintained throughout the period, a perennial biomass level above the value (3000 kg DM/ha, figure 4a) at which root partitioning would start not to limit the above-ground biomass production (Sim et al., 2015). This result is in agreement with previous reports showing that plants less severely defoliated maintain a higher photosynthetic leaf residual and therefore shows a higher root biomass than plants cut more severely (Hildebrand and Harrison, 1939; Langer and Steinke, 1965; Meuriot et al., 2005).
Therefore, in our work, the lower amount of perennial biomass and nitrogen content of plants cut at 4 cm may be due to a lower photosynthetic capacity in the residual (e.g. lower LAIRES and NFERES, figure 3a) which was not enough to supply the resources demand exerted by the aerial sinks after the cutting (Ryle and Powell, 1975). In this respect, in lucerne, the aerial sinks exert a high control of carbon metabolism during post-cutting regrowth (Frankhauser and Volenec, 1989). Consequently, in plants cut at 4 cm there would be less availability and/or less partitioning of carbohydrates to the perennial biomass affecting negatively their carbon balance (Davidson et al., 1990; Meuriot et al., 2005).
Management implication
The results obtained in the present work demonstrate that, during the establishment year, a defoliation management that contemplates a higher post-cutting foliar residual, allows to reconcile the need to rapidly establish a vigorous root system (Sim et al., 2015) with the need for early and frequent grazing to maximize the secondary production in lucerne pastures (Berone et al., 2020). In turn, the lack of differences between the cultivars evaluated indicates that the recommendation to increase the residual leaf area during the year of establishment applies to both non-dormant and semi-dormant cultivars.
The dead material of the residual biomass increased strongly in the plants cut at 10 cm in height deriving in a cleaning cut (4 cm in height) towards the middle of summer, as was already mentioned (material and methods and result sections). It is important to mention that the presence of dead material did not affect the growth capacity of plants (‘mid-summer’ pasture growth rates: 10 cm in height= 145-180 kg DM/ha/day; 4 cm in height= 120-150 kg DM/ha/day). In turn, the cleaning cut did not have negative consequences on the defoliated plants at 10 cm in height. These results suggest that, if necessary, a midsummer cleaning cut would not have negative consequences on lucerne plants that were previously defoliated at 10 cm in height.
Another implication emerging from the present research is about the need for autumn rest in lucerne pastures. A resting lucerne is generally suggested in late summer– early autumn until approximately 50% flowering to allow root reserves to be restored before winter (Teixeira et al., 2007; Berone et al., 2020). Longer recovery intervals in autumn (~500-600 GDD) have been found to be important for productivity and persistence in high northern latitudes where cutting lucerne prior to winter can greatly reduce the persistence of plants (Bélanger et al., 1999). Such recommendation can be hard to follow for farmers because, in late summer– early autumn, the other forage resources are not available to be used (e.g. pastures recently sown, crops not harvested as silage) and the green lucerne pastures is a high valuable feed resource. Considering the results obtained in the present work, the need for such autumn rest would be more relevant in plants frequently and severely defoliated (i.e. every 370 GDD and 4 cm in height). However, if defoliations are not severe (i.e. every 370 GDD and 10 cm cutting height), the levels of perennial biomass and N reserves observed (figure 4) suggest that, for humid and temperate environments, the need for such a rest would be less relevant. Of course, to be more conclusive on this a direct comparison of defoliation strategies during late summer– early autumn (e.g. frequent/not severe vs. infrequent/severe) is needed.
CONCLUSIONS
In lucerne cultivars with contrasting degrees of winter dormancy, reducing the severity of defoliation during the year of planting allows maintaining a forage yield similar to those of more severely defoliated plants under high defoliation frequency. At the same time, less severely defoliated plants showed higher biomass and N levels in their perennial organs (i.e. root plus crown). Consequently, a defoliation management that contemplates a higher post-cutting foliar residual allows, during the establishment year, to reconcile the need for early use of lucerne pastures with the need to establish a vigorous perennial system.

