











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkINTRODUCTION
Cotton is the main source of natural fiber worldwide, used for textile industry and other purposes such as oilseed and animal feed. This crop is cultivated in nearly 100 countries, occupying about 2.5% of the world’s arable land, under a great diversity of agroclimatic conditions and widely varied farming practices, being of great importance for the economy of such countries (Boopathi et al., 2015; Scarpin et al., 2022; Zhang et al., 2008). Upland cotton (Gossypium hirsutum L.) represents about 95% of the global cotton production, due to its environmental adaptability, high production, and better yield potential (Ijaz et al., 2019).
Regarding plant breeding, numerous technologies have emerged over the past decades to join the conventional crossbreeding, such as genomic selection (i.e. molecular marker-assisted selection), gene editing and mutation breeding (Jiang, 2016; Suprasanna et al., 2015; Voss-Fels et al., 2019). Induced mutations have been successfully used in plant breeding programs to artificially generate genetic variation for the development of new varieties with novel traits. Since the 1920s, more than 3,400 mutant varieties have been released worldwide, including cereals, oilseeds, as well as vegetables and industrial crops (https://nucleus.iaea.org/sites/mvd/SitePages/Home.aspx). Specifically, in cotton, the NIAB‑78 and NIAB‑846 varieties, obtained by irradiation, present heat and bollworm tolerance and improved yield and fiber quality, respectively (Aslam et al., 2018; Maluszynski et al., 2009). In addition, a cotton variety with tolerance to imidazolinones was recently released, which was obtained by mutagenesis using sodium azide (Tcach et al., 2022).
Although mutations occur spontaneously in nature, their frequency is very low, and can be increased by exposing biological material to mutagens (Landau et al., 2021; Mba et al., 2010; Oladosu et al., 2016).
The chemical mutagens most commonly used in plant breeding are: ethyl methanesulfonate (EMS), sodium azide (SA) and N-methyl-N-nitrosourea (MNU) and they mainly produce point mutations. Physical mutagens most commonly used are gamma- or X-rays, which often cause large deletions, leading to loss of several genes, DNA breakage and chromosome aberrations.
The first step of breeding mutation is to optimize the mutagen dose to achieve a high mutation rate while avoiding unintended damage, serious effects on germination and plant development (Ke et al., 2019; Mba, 2013; Mba et al., 2010; Nielen et al., 2018b; Oladosu et al., 2016). Initial studies on induced mutations have mainly aimed at finding the optimum combination of mutagen and dose to elicit the best response.
The level of damage induced on the M1 generation determines the number of seeds that can be used in the M2 or M3 populations to select the desired phenotype. Several parameters can be assessed to determine the somatic effects after a mutagenic treatment, including delay in seed germination, level of disturbances in the cell cycle, frequency of chromosomal aberrations in meristematic tissues, reduced seedling emergence, reduced seedling and plant growth, appearance of chlorophyll deficiencies, reduced fertility and plant survival to maturity, and sterility (Maluszynski et al., 2009).
In the last decade, some researchers have reported mutagenesis experiments on cotton, which include treatments using seeds or somatic embryos as plant material, and employing EMS, SA, gamma or X-rays. These studies have been used for morpho-physiological feature analyses, selection of lines with enhanced agronomical traits such us fiber quality and yield or stress tolerance among others, which could also serve as a valuable resource for functional genomic research on complex allotetraploid traits (Abid et al., 2020; Aslam et al., 2016, 2013; Auld et al., 2007, 1998; Bechere et al., 2010, 2007, 2014; Brown et al., 2012; Ganesan et al., 2005; Kalwar and Dahot, 2017; Lian et al., 2020; Muthusamy and Jayabalan, 2013, 2011; Patel et al., 2014; Shim et al., 2019; Tcach et al., 2022; Witt et al., 2018a, 2018b). In those mutagenesis studies, most experiments have followed previously published basic protocols, but few cotton-specific works have assayed dose sensitivity with different type of mutagens in cotton seeds.
In this context, the aim of the present work was to comparatively evaluate the phenotypic effects of EMS, SA, X-rays and a combined treatment of SA+X-rays on M1 plants coming from seeds of an elite variety of cotton (Guazuncho 3) as a guidance to choose the most adequate doses of each mutagen to generate an M2 population in order to select mutants of interest for a cotton breeding program.
MATERIALS AND METHODS
Plant material
Experiments were performed using seeds from tetraploid cotton (G. hirsutum) of the variety Guazuncho 3, belonging to the germplasm collection of the EEA INTA Saenz Peña, Chaco, Argentina. This line is an intermediate-short cycle crop, with good yield and quality features, high percentage of ginning, intermediate vigor, low number of vegetative branches and low tolerance to abiotic stress (Bonacic Kresic et al., 2004).
Mutagenic treatments
The mutagenic treatments were carried out at Instituto de Genética “Ewald A. Favret” (IGEAF) CICVyA-CNIA-INTA. A total of four doses were evaluated for each mutagenic treatment as summarized in table 1, using 100 healthy seeds, coming from the same production lot, for each assay. No special handlings (including delinting) were applied to the seeds before each mutagenic treatment.
For X-rays treatments, dry seeds were irradiated with an X-rays generator Philips MG160 (160 kV max. and 30 mA max.) at the indicated doses at 120 kV and 15 mA. For SA mutagenesis, different concentrations were prepared in phosphate buffer at pH 6.0, and seeds were incubated in each solution for 18 hrs at 18°C and 165 rpm in an orbital shaker. Then, the seeds were thoroughly washed seven times with water and air-dried. Similarly, EMS treatments were performed by incubating the seeds in the corresponding dilution following the same treatment conditions mentioned above for SA. For the combined treatment (SA+X-rays), the SA protocol was carried out first, and then the air-dried seeds were subjected to a 100 Gray (Gy) X-rays dose. Finally, the same number of seeds without mutagenic treatment was used as control.
Plant material handling after mutagenic treatments
The treated and control seeds were sown in trays of 0.5 m x 0.35 m, using inert substrate previously sterilized by autoclave. These trays were placed in a growth chamber at 30°C for 15 days to evaluate germination and other parameters described below. Then, 15 representative seedlings from each mutagenic treatment and control were randomly selected, transplanted into 5-liter pots (2.5 kg soil/pot) and grown under semi-controlled conditions in a greenhouse for further studies. In this step, only 15 plants of each treatment were selected due to the limited capacity of the greenhouse and personnel to handle them.
Evaluation of different mutagenic treatments in generation 1 (M1)
For each treatment, the number of emerged seedlings (fully expanded cotyledons) was evaluated 15 days after sowing to determine the germination percentage (GP). The germination velocity index (GVI) was also calculated as described in Meneses et al. (2011) and Nakagawa (1994). Finally, survival percentage (Sv) was calculated as the total emerged seedlings divided by the number of plants that emerged and continued their development until the complete expansion of the second true leaf (Spencer Lopes et al., 2018). Growth rate (g/days) on vegetative stage was also determined as: (plant weight 40 days after emergence – plant weight 20 days after emergence)/20 days.
Additionally, some morphological features were evaluated during the entire growth period, such as the presence of malformed plants or off-type plants, chlorophyll deficiencies or chimeras. Furthermore, phenological data were recorded from 15 plants selected from each treatment that were transplanted into pots as described below, until maturity.
Post-harvest analysis
At plant maturity, some agronomical and morphological traits were analyzed in the 15 selected plants from each mutagenic treatment: i) plant height, ii) fruit retention percentage, which is the percentage of retained fruits in the plant over the total reproductive positions [(N° retained fruits/N° of reproductive positions)*100], iii) number of fruit abortions, identified by scars instead of developed fruits retained on the plant, iv) number of reproductive positions, v) harvest index (HI), which is the ratio of harvested product (lint and seed) to the above ground plant dry weight or biomass (stems, leaves and fruit), and vi) number of seeds per boll.
Statistical analysis
Statistical analysis was carried out using general linear and mixed models with the statistical program InfoStat (Di Rienzo et al., 2011) through its interface with the R program. Fisher’s LSD test was used to compare the mean values at a significance value of 5% (p <0.05).
RESULTS
Effects of mutagenic treatments on seed germination, survival and vegetative growth rate
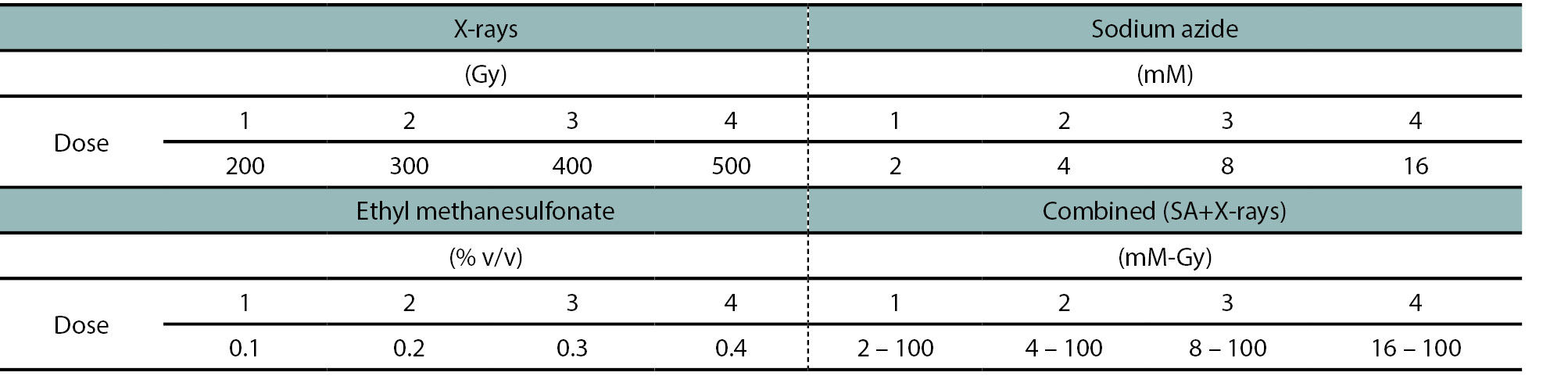
In order to determine the most suitable dose of each mutagen or treatment for cotton and to analyze the differences between them, some important phenotypic characteristics, such as germination percentage (GP), germination velocity index (GVI) and seedling survival were studied. The parameters analyzed showed differences in germination depending on the mutagen employed and dose, as shown in supplemental figure 1. The major effect on GP of M1 cotton seeds was observed in EMS treatments, where it strongly decreased as the concentration of mutagen increased, with an R2 of 0.96 for its regression (p-value: 0.003), attaining only 20% of germinated seedlings with 0.4% EMS (figure 1A). Additionally, SA treatments showed significant differences in GP (p-value: 0.029) between doses, presenting an R2 of 0.84. However, no significant differences were observed in the GP values for the X-rays treatments, as well as in the combined treatment (SA+X-rays) as the mutagen doses increased (p-value: 0.196 and 0.081, respectively) (figure 1A).
Figure 1B shows the GVI obtained in this study. Considering that a low index could be attributed to a low number of germinated seeds and/or a delay in germination, it was considered that GVI should be analyzed in conjunction with GP results. EMS treatments showed a clear decrease in GVI values as the concentration of mutagenic agent increased, resulting in a GVI 10-fold lower at 0.4% EMS, compared to the control (figure 1B). In contrast, X-rays treatments showed non-significant differences for this index among the doses applied.
Interestingly, SA assays exhibited a lower GVI at higher doses, leading to a 2-fold reduction at the highest SA concentration compared to the control (figure 1B). In addition, in combined treatments (SA+X-rays), a similar but less pronounced behavior was observed compared to SA (figure 1B).
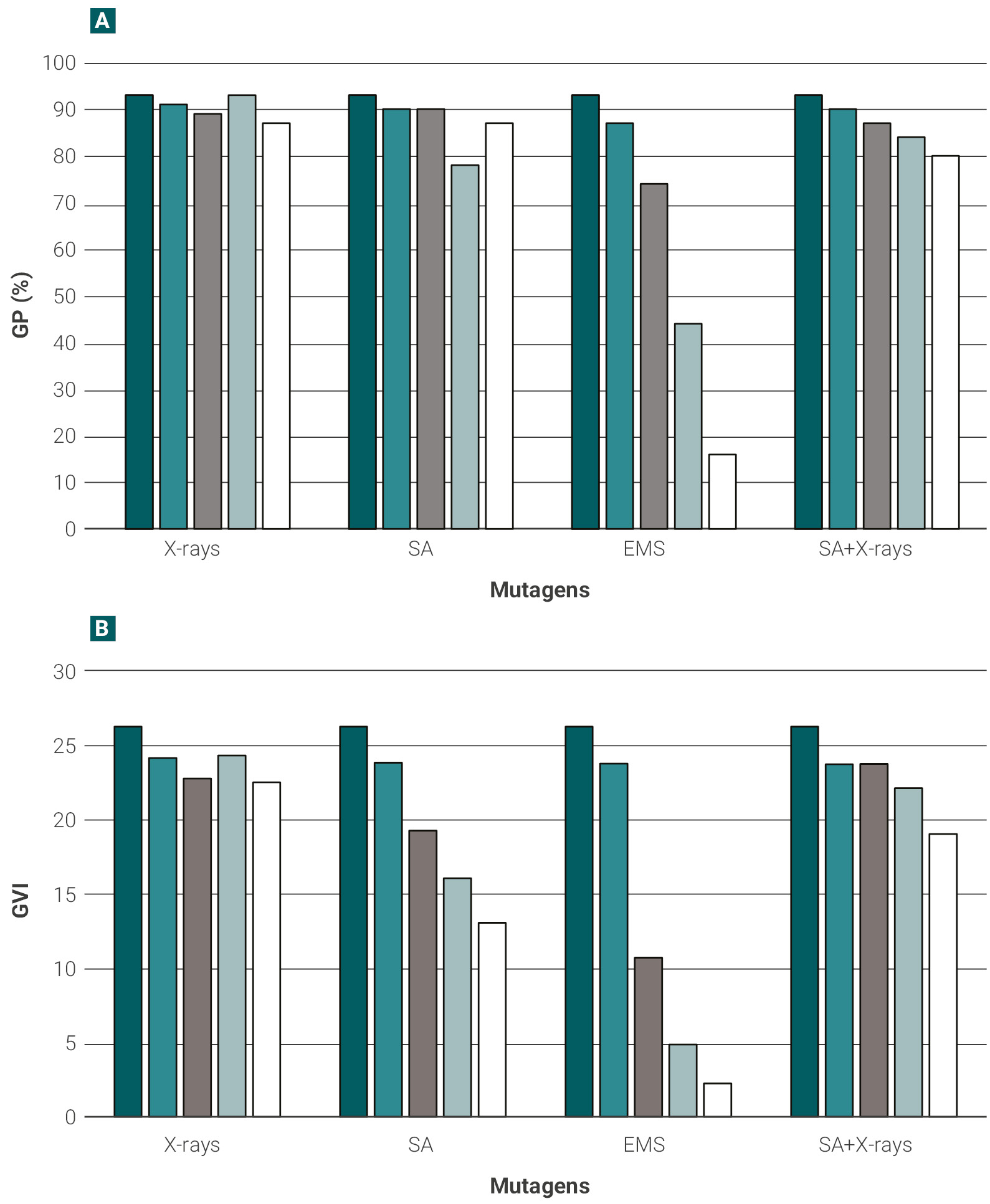
Figure 1.Germination percentage (A) and germination velocity index (B) of Guazuncho 3 cotton seeds under mutagenic treatments. Results obtained for the untreated control plants (dark blue bars), dose 1 (blue bars), dose 2 (gray bars), dose 3 (light blue bars) and dose 4 (white bars), for each mutagenic treatment. EMS: ethyl methanesulfonate, SA: sodium azide.

Supplemental figure 1.Cotton seeds germination. The figure shows the obtained seedlings with the different mutagenic treatments and doses whose quantification is presented on figure 1.
Figure 2 shows the survival of the seedlings over the total number of germinated plants for each dose of the mutagenic treatments assayed. The most damaging treatment was the highest concentration of EMS (0.4%), causing more than 60% of plant mortality, while the survival of seedlings was above 66-73%, for SA, X-rays treatments or their combination, even for the highest doses (figure 2).
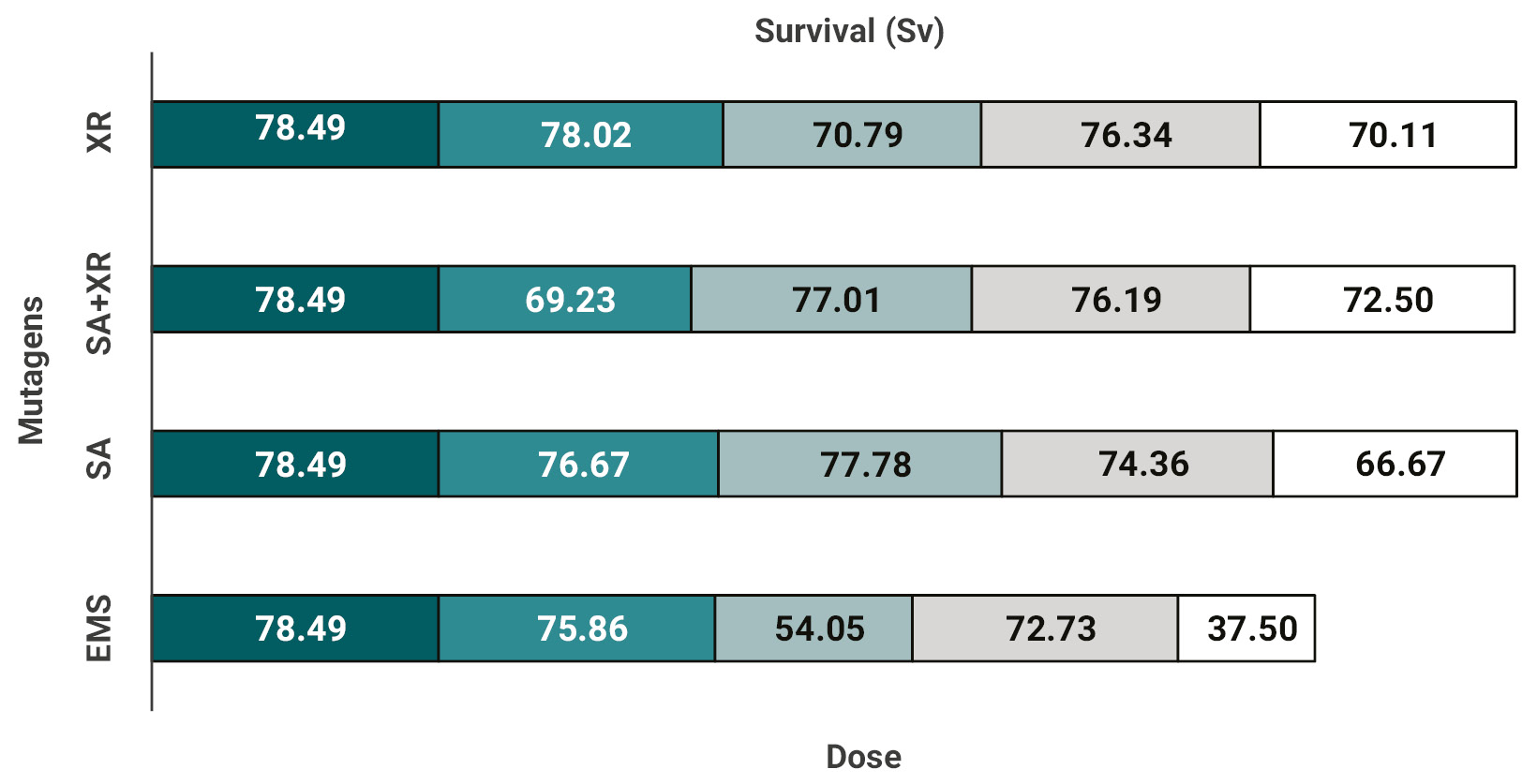
Figure 2.Seedling survival percentage. Percentage of seedlings that survived until the second leaf was fully expanded, over the total number of germinated seeds, under the different mutagenic treatments and doses. Dark blue bars (control), blue (dose 1), light blue (dose 2), gray (dose 3), white (dose 4). Survival percentage was determined as described in Materials and methods. EMS: ethyl methanesulfonate, SA: sodium azide.
Figure 3 shows the effect of the different mutagenic treatments and their doses on growth rate of cotton at vegetative stage. Both chemical mutagens (EMS and SA) generated a significant reduction in the growth rate, being more pronounced for EMS, which produced almost no growth per day at the highest dose. On the other hand, with X-rays and the combined (SA+X-rays) treatments the growth rate in the evaluated stage was not significantly affected.
Overall, for the germination and seedling stage parameters determined in this study, the EMS treatments produced greater effects on the cotton variety used (Guazuncho 3) than the other mutagens tested, even in the SA+X-rays combination. Finally, the X-ray treatments showed small changes in the survival of seedlings, but no differences were observed on GP or GVI, while their combination with SA exhibited a distinctive behavior, probably due to SA.
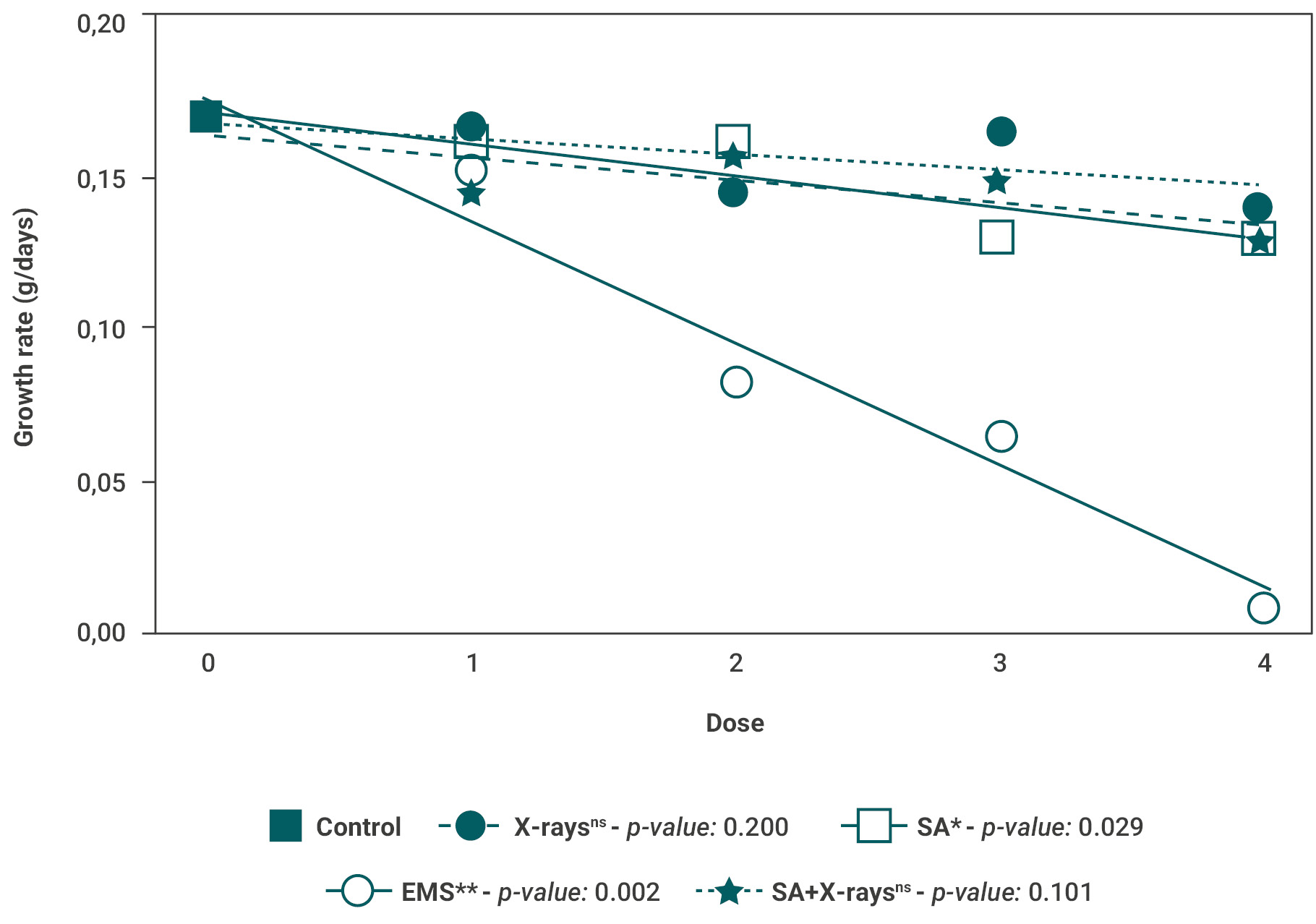
Figure 3.Growth rate vs. mutagen dose determined in vegetative stage. The growth rate was calculated as described in Materials and methods. EMS: ethyl methanesulfonate, SA: sodium azide.
Phenological and phenotypic alterations in M1 plants
Cotton (Guazuncho 3) showed a significant delay in the development of the vegetative stage when treated with X-rays or EMS, as presented in Figure 4. The time from seedling emergence to squaring for the X-rays treatments was of 54.3 and 53.8 days for 300 Gy (dose 2) and 500 Gy (dose 4) respectively, compared to 48 days in control plants (figure 4). Similarly, the plants from trials with 0.3% and 0.4% EMS exhibited the longest emergence-squaring time, 60.9 and 57 days, respectively, while for SA and SA+X-rays treatments no significant differences were found at this stage regardless of the dose used (figure 4). Although all doses tested for EMS generated a shorter time to reach maturity (from the first open boll to the harvest time) compared to the control, the shortest time (25.2 days) was obtained in 0.3% EMS-treated plants, while 34.4 days were recorded for the control plants (figure 4). Finally, no significant differences were found in SA or SA+X-rays mutagenic treatments regardless of the doses tested.
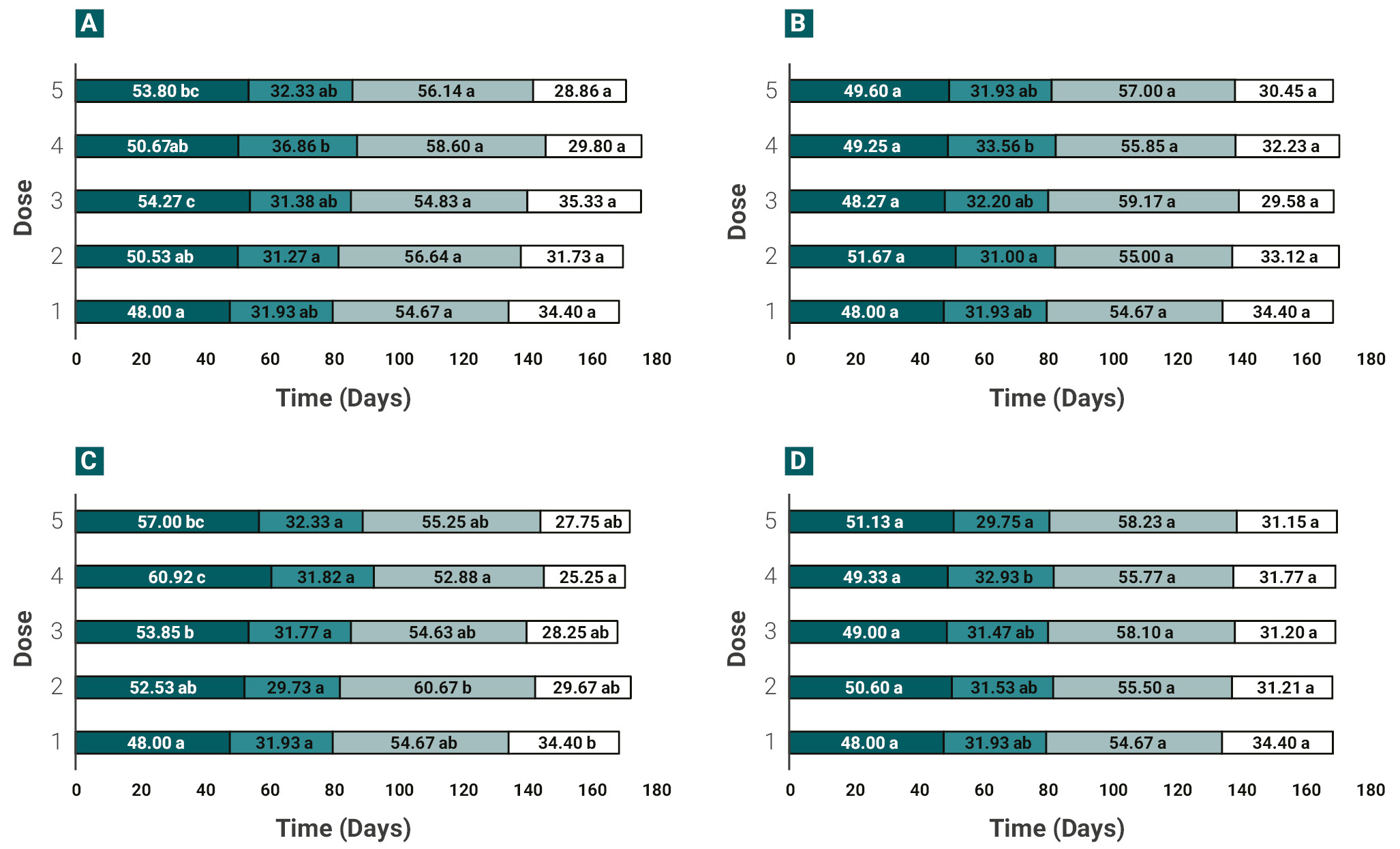
Figure 4.Phenological development of cotton crop under different mutagenic treatments. (A) X-rays, (B) SA, (C) EMS and (D) SA+X-rays in different doses. Phenology was divided in: the number of days from seedling emergence to squaring (dark blue; vegetative stage), number of days from first square to first flower (blue), days from first flower to first open boll (light blue) and days from first open boll to harvest time (white). EMS: ethyl methanesulphonate, SA: sodium azide. *<0.05, **<0.01, ***<0.001, ns: no significance.
Phenotypic alterations were also observed in some treated plants (supplemental figure 2). Phenotypic effects were classified into two groups: i) Malformed or off-type plants, and ii) Plants with chlorophyll deficient irregular-shaped spots as indicators of chimeras. Regardless of the applied dose of the mutagen, phenotypic variations were grouped and analyzed according to the type of mutagen (figure 5). Results indicated that more than 50% of the total plants with malformations came from EMS treatments, followed by X-rays (around 30%), while the SA treatments and the combination SA+X-rays resulted in less than 10% of malformed or off-type plants (figure 5A). Similarly, X-rays and EMS mutagenic treatments showed the highest proportions of plants with chlorophyll deficient irregular-shaped spots accounting for 50% and 44%, respectively. Whilst only 5% corresponded to the combined mutagens (SA+X-rays) and 2% of plants treated with SA presented this type of phenotypic effects (figure 5B), this chlorotic effect is even less marked for the latter compared with X-rays or EMS (supplemental figure 2 H-I).

Supplemental figure 2.Representative pictures from M1 mutagenized cotton seedlings and plants with observed abnormal phenotypes. A: control plant, EMS-treated seedlings/plants with malformations (B-C-D), chlorotic regions and malformed branches (E-F), malformed EMS plant at maturity (G), chlorotic effect with SA (H) and with X-rays (I).
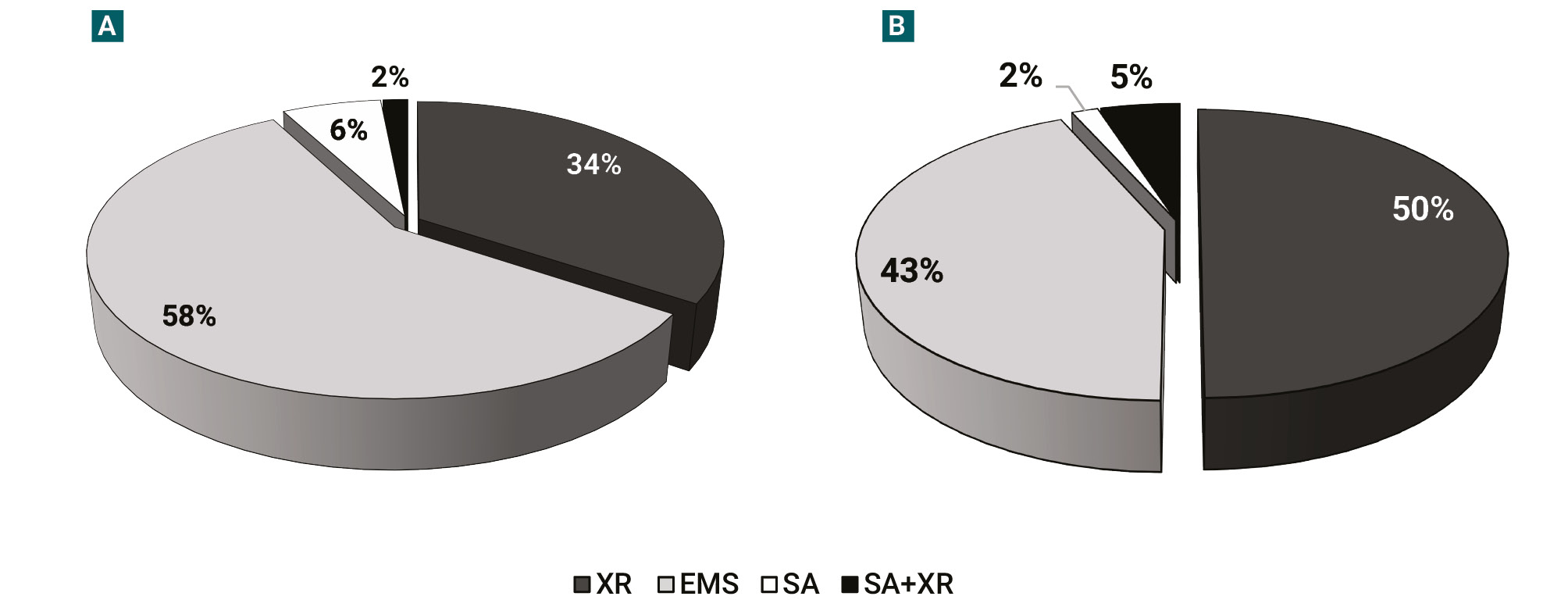
Figure 5.Phenotypic effects of mutagenic treatments on cotton seedlings and plants. (A) Percentages of malformed or off-type plants.(B) Plants with chlorophyll deficient irregular-shaped spots. EMS: ethyl methanesulfonate, SA: sodium azide.
Morphological and agronomic traits in selected plants
At maturity or harvest time, some morphological and agronomic characteristics were determined in 15 selected plants from each mutagenic treatment. Both EMS and SA showed differences in plant height among the doses used (supplemental table 1). Plants coming from SA treatments exhibited a slight increase in height at lower doses compared to the control, while this effect reverted with the higher concentration of SA. Plant height from EMS treatments decreased as the dose increased, reaching only 16.27 cm at 0.4%, compared with the control plants (supplemental table 1).
Other agronomic traits, such as the percentage of retained fruits, the number of reproductive positions and the harvest index (HI), were affected in plants subjected to mutagenic treatments. A reduction of 2.7- and 1.8-fold was determined for retention percentage and number of reproductive positions, respectively, with 500 Gy (dose 4) of X-rays. However, no significant differences were found for the number of abortions and the number of seeds per boll in plants that were treated with X-rays (supplemental table 1). Likewise, the EMS mutagen triggered similar effects on these evaluated agronomic traits, reducing fruit retention and the number of reproductive positions by 4.4- and 4.0-fold, respectively, at the highest dose (0.4%). Nevertheless, the plants treated with EMS conserved their few reproductive positions, since they presented fewer abortions when the dose increased, showing a mean of 3.50 and 1.60 abortions with 0.3% and 0.4% EMS, respectively (supplemental table 1). In addition, no significant differences were obtained for the HI or number of seeds per boll. The trial with SA did not show differences in the agronomic characteristics studied, nor did the combination of SA+X-rays.
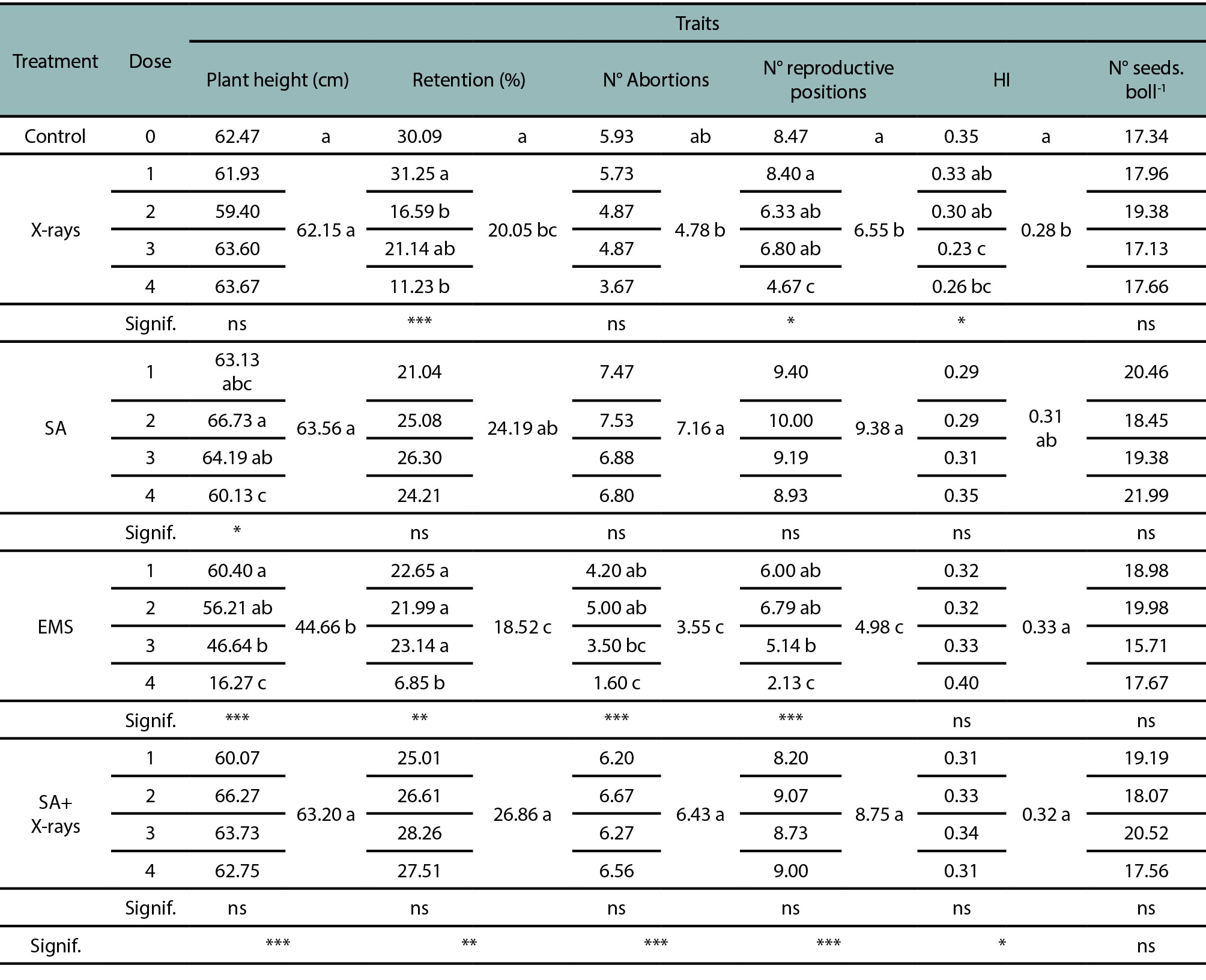
Supplemental table 1.Morphological and agronomic traits analyzed in different treatments and doses. Parameters were evaluated at maturity. Retention: percentage of retained fruits on the plant, N° abortions: number of bolls that do not develop fully, HI: harvest index. Signif.: statistical significance, ns: no significance, *<0.05, **<0.01, ***<0.001.
DISCUSSION
The dosimetry of any mutagen is a crucial aspect for a mutation breeding program. In cotton, mutants for diverse characters have been obtained using different mutagenic agents at distinct doses (The Joint FAO/IAEA Mutant Variety Database (MDV) https://nucleus.iaea.org/sites/mvd/SitePages/Home.aspx). Tcach et al. (2022) isolated a mutant tolerant to imidazolinones in advanced cotton lines using 1 mM sodium azide pH 3 or 6. However, Ul-Allah et al. (2019) concluded that sodium azide doses of 15 and 25 mM induced genetic variation in cotton germplasm suitable for use in a breeding program in nine different genotypes. In addition, Auld et al. (1998) generated variability in fiber quality parameters in the upland cotton variety HS 2000 using 3% v/v EMS. Similar studies were described by Brown et al. (2015). Comparable doses of EMS (2.45%) were used by Cutts (2013) in different germplasm that exhibited herbicide tolerance. Conversely, Deho et al. (2020) observed differences in morphological and agronomic parameters in eight mutant lines with 0.03% EMS. Regarding cotton treatments with ionizing radiation, there are several mutants obtained mostly with gamma rays annotated in the IAEA MDV using different doses (https://nucleus.iaea.org/sites/mvd/SitePages/Home.aspx).
So, there are many differences in the range of doses between varieties of the same crop, making it difficult to establish a correlation between them. This might happen because the dose and type of mutagen can vary between genotypes and it is therefore advisable to perform a dosimetry in each case before the final treatment.
Germination Percentage
Both chemical mutagens, SA and EMS, significantly reduced the GP compared to the controls. It is well known that the SA and EMS have an effect over seed germination, delaying or decreasing it (Espina et al., 2018; Gruszka et al., 2012).
The greatest effect was observed in the EMS treatment, which produced an abrupt reduction in the number of germinated seeds as the dose was increased. High doses of radiation or chemical agents produce physiological and biochemical alterations, i.e. they can alter hormones such as auxins, produce changes in the content of ascorbic acid, generate free radicals and increase the α-amylase activity, among others. Besides, some authors suggest that this reduction in seed germination could be due to an effect of mutagens in the meristematic tissue of the embryo. Together, these alterations inhibit vital functions that can result in the death of certain cells and even the embryo (Kurtar et al., 2017; Lagoda, 2012).
Although there were significant differences in GP for SA treatments, an increase in germinated seeds was observed with the highest dose evaluated. Similar results were obtained for other cotton germplasm by Baimuhametova et al., (2017), who even recommended 1 to 5 mM SA for cottonseed treatment. These authors suggested that SA has high mutagenicity and at certain concentrations this chemical could have a stimulating effect on germination. Additional studies showed a stimulating effect of SA on germination of pure lines in wild oats (Avena fatua L.) (Upadhyaya et al., 1982) due to an alternative respiration mechanism or pathway, since a stimulation in oxygen uptake was observed in the presence of SA (Upadhyaya et al., 1983). Nevertheless, more studies are needed to confirm the relationship between the alternative respiration by SA and the stimulation of germination.
On the other hand, no effects on germination of cotton seeds were observed with X-rays regardless of the dose used. These results are in agreement with studies that suggest that ionizing radiation only affects cell division and not the elongation of predetermined cells, because large damage to DNA will be evidenced at replication at cell division, thus not affecting germination (Prina, 1989). However, other studies showed a decrease in GP as the ionizing radiation dose increased in different crops, such as soybean (Mudibu et al., 2012), linen (Bhat et al., 2017), wheat (Ahumada-Flores et al., 2021), sorghum (Wanga et al., 2020). Results from Aslam et al. (2013) showed more than 50% decrease in GP at 300 Gy of gamma rays treatment in cotton, in contrast to what was observed in this work.
It is important to consider that the level of the effects observed from irradiation treatments in the different germplasm can be affected by different biological, environmental and chemical factors, such as, the water content of the seeds, the atmosphere, temperature, storage after irradiation, genetic differences, cell cycle phases, etc. (Spencer Lopes et al., 2018). Thus, we could assume that our results are due to the type of seed, its hardness, as well as the absence of delinting and hydration of the seeds.
Germination Velocity Index
Mutagens have been reported to cause a delay in germination time, determined by the germination velocity index (GVI) (Adeosun et al., 2020; Khan et al., 2009; Nakagawa, 1994). EMS, SA and combined (SA+X-rays) treatments showed significant differences in GVI. EMS exhibited a clear tendency to decrease the GVI as the dose increased, mainly influenced by the number of germinated seeds than by the germination time. In contrast, our results showed that SA treatment did not affect the number of germinated plants, thus the decrease observed in GVI as the dose of SA increased was the result of a delay in seed germination. Finally, a similar but milder behavior compared to SA was observed in GVI for combined treatments. This lower phenotypic effect of the combined treatments could be attributed to a protection effect exerted by sodium azide against physiological damage, chromosomal damage and lethality caused by radiation, as it was previously suggested (Prina and Favret, 1983).
The delay in seed germination is a common effect in chemical mutagenic treatments such as SA. This can be attributed to the inhibition of necessary physiological and biological processes, including enzyme activities alteration, hormonal imbalance, and inhibition of mitotic processes. The metabolite formed between azide and alanine, L-azidoalanine, causes mutations by interacting with enzymes and DNA in the cell, being a strong inhibitor of cytochrome oxidase, consequently inhibiting the oxidative phosphorylation process. Furthermore, azide acts as a potent inhibitor of the proton pump, altering mitochondrial membrane potential. Together, these SA-induced effects cause an obstruction in the main ATP biosynthesis pathway, which can directly influence the slowing of the germination rate (Gruszka et al., 2012; Khan et al., 2009; Leitao, 2012).
Survival
Previous studies have reported that the mutagens affect not only germination but also seedling survival (Sv), at least until second leaf is fully expanded, in different plant species (Ahumada-Flores et al., 2021; Aslam et al., 2013; Espina et al., 2018; Ingelbrecht et al., 2018; Kodym and Afza, 2003; Nielen et al., 2018a; Spencer Lopes et al., 2018). Plant survival was also affected by the mutagenic treatments. The highest percentage of seedling mortality was obtained with 0.4% EMS, being 3-fold higher compared to the control. While, for SA, X-rays and the combined treatment of both, a survival of approximately 70% was observed at their respective higher doses. Therefore, the results revealed that EMS caused more seedling lethality than X-rays and SA, and this reduction in survival rate could be associated with the toxic effect of EMS (Bhat et al., 2017). Besides, Espina et al. (2018) suggested that the effect of EMS on germinated soybean seedlings survival could be reduced due to cell damage and poor root and shoot development. This result has also been observed in pepper (Arisha et al., 2015) and cucumber (Shah et al., 2015). Finally, for SA a significant reduction in survival was also observed for the highest dose. Similar results were observed by Khan et al. (2009), who attributed this effect to physiological alterations and imbalance between promoters and inhibitors of growth regulators, which led to seedling death.
Growth rate and height
A reduction in seedling or plant growth in different crops after mutagenic treatments have been reported before (Ingelbrecht et al., 2018; Joshi et al., 2011; Spencer Lopes et al., 2018; Suprasanna et al., 2012). An increase in the concentration of the mutagen has often shown more physiological damage and direct DNA damage, biochemical disturbances, auxin destruction and changes in ascorbic acid content (Joshi et al., 2011). Together, these damages are phenotypically reflected in plant growth (Kodym et al., 2012).
Our results showed a significant reduction in the growth rate of cotton plants as the chemical treatment dose increased; EMS caused more reduction in plant growth than SA. Also, a significant effect on plant height was observed in both chemical treatments, obtaining a linear reduction in height in EMS treated plants as the dose increased, similarly to previous studies in cotton (Aslam et al., 2013).
Interestingly, a low dose of SA recorded taller plants compared to the control. This effect was also previously observed in cotton, Calendula officinalis L., Salvia splendens, Amaranthus, onion (Baimuhametova et al., 2017; El-Nashar and Asrar, 2016; Joshi et al., 2011). These results could be explained by “hormesis” effect, which has been defined as a favorable response to exposures at low levels of adverse conditions (Jalal et al., 2021). Hypothetically, this mild stimulation of plant height would be related to an increase in the rate of cell division, as well as by an activation of growth hormones such as auxins (Joshi et al., 2011). Although no significant differences were obtained in growth or height for X-rays treatment in our study, the hormesis effect has been mainly observed in ionizing radiation treatments (Nielen et al., 2018b).
Phenology
EMS and X-rays treatments produce chromosomal aberrations that can negatively affect cell division (Hadebe et al., 2018). In this context, these damages could be responsible for the observed significant delay in the vegetative stage of cotton plants treated with these mutagens. On the other hand, SA+X-rays combination did not show significance compared with X-rays treatment. As it was discussed previously, the differences between these treatments could be attributed to a protection effect exerted by SA against the damages caused by radiation (Prina and Favret, 1983). Additionally, a reduction in maturity time was observed at the end of the plant cycle (crop maturity) for EMS treatment, in higher doses compared with the control.
Somatic mutations
Morphological changes such as variations in leaf shape, dwarfism, branching pattern, physical lesions and chimerism were observed in our study. These phenotypic characteristics have been commonly observed in different crops after mutagenic treatments since they are easily measurable in M1 generation and are good indicators of mutagenic efficacy (Abid et al., 2020; Aslam et al., 2013; El-Nashar and Asrar, 2016; Espina et al., 2018; Favret, 1960; Landau et al., 2021; Lian et al., 2020; Prina et al., 2012). Our results showed that EMS and X-rays treatments originated the highest number of malformed plants and irregular-shaped spots with chlorophyll deficiencies.
Chimerism of M1 can be observed as irregular spots in dicotyledonous plants. The sectors with notable changes in the amount of chlorophyll, called chlorophyll-deficient sectors, are easily seen and are usually counted to estimate the frequencies of induced somatic mutations (Prina, 1989).
Prina (1989) pointed out a relationship between somatic mutations and mutations in germ lines in barley after treatments with different mutagens. A ratio of M2 mutations to M1 mutated sectors was established, which was approximately 0.4 for X-rays, 2.5 for EMS, and 6 in SA treatments. The author assumed that the frequency of somatic mutations in M1 can serve as an indicator of the frequency of mutations to be expected in M2, the generation where a selection process can begin (Prina, 1989). However, these values or relationships between somatic mutations (M1) and germinal mutations (M2) are specific to each crop and species. Although this ratio has not been established for cotton yet, the somatic mutations observed and recorded in the present work are good parameters to indicate the level of damage per dose for each mutagenic agent.
Fertility or sterility
M1 plants typically show reduced fertility, mainly caused by chromosomal rearrangements and genomic mutations during meiosis (Lagoda, 2012). Reduced fertility is also considered as a reduction in reproductive capacity, including a severe growth retardation or growth inhibition that prevents flowering, formed flowers lacking reproductive structures, abortion of pollen or ovules, abortion of fertilized embryos before maturity and formed seeds that do not germinate properly or die after germination (Nielen et al., 2018b). Therefore, fertility or sterility could be assessed as the number of flowers, fruits and seeds produced at crop maturity.
In our work, fertility was considered as the number of seeds produced per boll that reached physiological maturity. No significant differences were found between treatments or between doses. All the plants that reached the reproductive stage produced seeds, considering then that there were no mutagenic effects on meiosis.
On the other hand, significant differences were observed in the number of reproductive positions, abortions and fruit retention percentage. The greatest effects were recorded in EMS and X-rays treatments, assuming that the effects on these variables could be mainly due to physiological effects (Nielen et al., 2018b).
CONCLUSIONS
The treatments with EMS were the most aggressive for cotton seeds, since they showed the greatest number of plants with altered phenotypic effects (PG, height, survival, etc.), followed by X-rays and then SA treatments. Finally, the combined treatment SA+X-rays showed the least efficacy for the evaluated traits. According to our results, the intermediate doses (300 Gy, 4 mM, 0.2%, for X-rays, SA and EMS, respectively) would be the most recommended for cotton (Guazuncho 3 elite variety). In summary, this study provided an approximation of the most appropriate doses of each mutagen to use to generate the M1 population. The frequencies of mutations generated by each of these doses can only be evaluated in the M2 generation.

