











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkINTRODUCCION
En la Región Pampeana Argentina, los suelos ocupados por pastizales presentan ambientes edáficos que pueden restringir el crecimiento vegetal, principalmente debido a deficiencias nutricionales, salinidad y déficit o exceso de humedad (Rodríguez y Jacobo, 2012). En este sentido, la producción de pasturas en el sudeste de la Provincia de Buenos Aires, es altamente dependiente de la disponibilidad de fósforo (P) y nitrógeno (N). Así, en la última década, dado que el contenido de P disponible en los suelos ha disminuido, la fertilización fosfatada constituye una práctica generalizada (Sainz Rozas et al., 2019). Por ello, resulta importante la capacidad de una especie para asociarse con microorganismos simbiontes que puedan mejorar su acceso al P disponible, tales como los hongos micorrícicos arbusculares (HMA). Varios autores han sugerido a los HMA como indicadores para monitorear los efectos del cambio de uso de la tierra y la salud del suelo (Nogueira et al., 2016; Silva et al., 2017; Šarapatka et al., 2019). Los HMA, a través de la colonización de raíces de las plantas hospedadoras establecen micorrizas, constituyendo el tipo de asociación simbiótica más extendida en la naturaleza entre dichos hongos (subphylum Glomeromycotina) y la mayoría (>72%) de las plantas vasculares (Brundrett y Tedersoo, 2018; Spatafora et al., 2016). Uno de los beneficios de la simbiosis es una mayor absorción de nutrientes (donde los poco móviles como el P resultan de mayor importancia), y consecuentemente en crecimiento, en plantas micorrizadas (Astiz Imaz et al., 2014; Barea et al., 2009). La mayor absorción por plantas micorrizadas es resultado del aumento de volumen de suelo explorado por la raíz micorrizada donde el micelio extraradical de los HMA tienen acceso al P más allá de la zona de agotamiento. La simbiosis es del tipo mutualista y obligada para los HMA dado que reciben fotosintatos por la planta hospedadora al hongo con el consecuente costo para el hospedador que la simbiosis conlleva (Allen et al., 2020). Es conocido que la formación de micorrizas se reduce en suelos con altas concentraciones de P disponible en la mayoría de las condiciones (Grant et al., 2005). Covacevich et al. (2007) establecieron que contenidos de P-Bray hasta 27 mg/kg en suelos del sudeste de la Provincia de Buenos Aires deprimen la colonización micorrícica a razón de 2,8% mg P/kg, y por encima de dicho valor la colonización se estabiliza al 10%. Asimismo, Covacevich et al. (2006) detectaron depresión de la colonización a campo en pasturas de agropiro y festuca alta resultado de la fertilización con fuentes de elevada solubilidad, aspecto que debería ser tomado en cuenta en la planificación de fertilización con fines productivos.
Festuca alta, Festuca arundinacea Schreb., es uno de los principales componentes de los pastizales y pasturas de la Pampa Deprimida Argentina (Roitman y Prelasco, 2018; Scheneiter et al., 2016). Se caracteriza por su rusticidad, plasticidad fenotípica y calidad en los sistemas extensivos de producción ganadera (Lattanzi et al., 2007; Mazzanti et al., 1992). Al igual que otras gramíneas C 3 puede establecer una relación simbiótica con hongos endófitos asexuales de transmisión vertical, es decir, a través de las semillas infectadas del hospedante (Clay y Schardl, 2002). El hongo endófito que infecta festuca alta, Epichloë coenophiala (Morgan-Jones & W. Gams) C.W. Bacon & Schardl (= Neotyphodium coenophialum (Morgan-Jones & W. Gams) Glenn, C.W. Bacon & Hanlin (Leuchtmann et al., 2014), crece sistémica y asintomáticamente en la parte aérea de la planta (Clay y Schardl, 2002). La asociación es considerada mutualista; por un lado, el endófito obtiene nutrición, dispersión y supervivencia, y por otro, las plantas infectadas se benefician por los alcaloides que produce el endófito (i.e., lolinas y peraminas) que les otorgan resistencia a adversidades bióticas y abióticas en condiciones de estrés (Lee et al., 2021; Malinowski y Belesky, 2000; Omacini et al., 2001). En este sentido, se ha demostrado, entre otros beneficios, que el endófito mejoraría la adaptación de las plantas hospedantes a la deficiencia de P, desarrollando raíces significativamente más largas y de mayor peso respecto a las producidas por plantas libres (Ren et al., 2007). Sin embargo, algunos de los alcaloides (i.e., ergoalcaloides) son tóxicos para el ganado en pastoreo y pueden causar diversos síndromes clínicos y subclínicos que provocan importantes pérdidas económicas en la producción ganadera (Evans et al., 2004). En los últimos años se han registrado valores superiores al 90% de infección endofítica en las plantas de festuca alta muestreadas en campos ganaderos de diferentes partidos de la Pampa Deprimida de la Provincia de Buenos Aires (Petigrosso et al., 2019).
Algunas investigaciones en gramíneas seleccionadas agronómicamente, han demostrado que la colonización de la raíz por HMA se reduce cuando el hospedante está infectado con endófitos (Antunes et al., 2008; Guo et al., 1992; Keenan & Rudgers, 2008; Liu et al., 2011; Omanici et al., 2006), sin embargo, otros estudios realizados en especies nativas, han registrado un aumento en la colonización (Novas et al., 2005, 2009, 2011; Vignale, 2016). Estos resultados estarían asociados a la especificidad de la asociación dentro de cada simbiosis y a su vez, a la posible interacción entre los endófitos Epichloë sp. y los hongos micorrícos arbusculares sobre la respuesta de la planta hospedante (Vignale, 2016).
Dado que los suelos de la Región Pampeana Argentina presentan baja disponibilidad de P, y que son escasos los trabajos que analizan la respuesta de plantas con endófito al P y su posible interacción con los HMA, el objetivo de este trabajo fue evaluar la producción de biomasa y su partición en plantas de festuca alta infectadas con endófito E. coenophiala e inoculadas con hongos micorrícicos arbusculares (HMA) en respuesta a la fertilización fosfatada (P). La hipótesis del presente estudio es que, en condiciones de baja oferta de P, las plantas infectadas con el endófito E. coenophiala e inoculadas con HMA, presentarán mayor producción de biomasa que aquellas libres de endófito y de HMA. En caso de verificarse este sinergismo, podría explicarse la mayor aptitud ecológica de plantas infectadas con endófito E. coenophiala y por consiguiente, su avance en pasturas y pastizales.
MATERIALES Y MÉTODOS
Sitio y diseño experimental
Se realizó un experimento bajo condiciones controladas en una cámara de germinación de la Unidad Integrada Balcarce, Facultad de Ciencias Agrarias, UNMdP - EEA Balcarce, INTA (37°45’ S, 58°17’ O, 130 m s.n.m.). Se trabajó con plantas de festuca alta con endófito silvestre E. coenophiala (SE+) y libre de endófito (SE-) pertenecientes a la misma población. La semilla original proveniente de la población naturalizada se cosechó en un pastizal del Partido de Mar Chiquita (37º 32’ S; 57º 55’ O) y se almacenó en heladera a 5°C para mantener la viabilidad del endófito. La semilla libre se obtuvo a partir de la eliminación del endófito mediante el almacenamiento de la semilla infectada a 20°C de temperatura y 52% HR durante 12 meses. El experimento se llevó a cabo con semilla cosechada en el mismo año y con comprobada condición endofítica (Petigrosso et al., 2021).
Se empleó un diseño completamente aleatorizado con arreglo factorial con 5 repeticiones. Previo a la siembra, las semillas de festuca alta se esterilizaron y se pusieron a germinar según las reglas International Seed Testing Association [ISTA] (2008). Una vez germinadas las semillas, se trasplantó una plántula de similar porte por cada maceta de 500 cc (unidades experimentales). Las macetas contuvieron sustrato homogéneo (1:2:1 volumen) formado por arena de río, perlita esterilizada y suelo esterilizado con formaldehído a una concentración de 10 µl/g de suelo, de modo de evitar la colonización por HMA nativos según Covacevich & Echeverría (2003). Para la condición micorrícica, se evaluaron dos niveles: con y sin inoculación de HMA (HMA+ y HMA-, respectivamente). Para la condición HMA+, se realizó inoculación de las raíces de las plántulas con esporas de HMA extraídas (empleando el protocolo descripto por el INRA, 2001) de muestras de suelo de los primeros 20 cm de un ensayo de larga duración, con predominio de soja (Glycine max L. Merr.) sin fertilización, ubicado en la EEA Balcarce sobre un Argiudol Típico con textura superficial franco limosa (Soil Survey Staff, 2014). Previo a la inoculación se determinó que los HMA mayoritarios en el inóculo estuvieron constituídos por morfotipos glomoides (géneros Glomus, Funneliformis, Rhizophagus). Al momento del trasplante, las plántulas fueron inoculadas con el consorcio de esporas de HMA, extraído antes de la esterilización del suelo (20 esporas/maceta). Además, a los 7 días desde el trasplante, se realizó una reinoculación con la misma dosis. Para la condición de fertilización con P se partió con un nivel inicial de 15 ppm y se evaluaron dos niveles: con y sin fertilización, es decir, sustrato fertilizado luego del trasplante con el agregado de 15 ppm de P (P15) y sin agregado de P (P0), respectivamente. Las macetas se mantuvieron a 25°C, con fotoperiodo (12 h luz/12 h oscuridad en cámara de crecimiento, durante 160 días donde fueron regadas con agua destilada cada 72 h. En total se usaron 40 macetas, resultantes de la combinación factorial de: 2 niveles de infección (SE+ y SE-), 2 niveles de inoculación (HMA+ y HMA-) y 2 niveles de fertilización fosfatada (P0 y P15) y 5 repeticiones.
Determinaciones
Cada vez que aparecieron tres nuevas hojas en las plantas de festuca (≈ 550ºCd, temperatura base de festuca alta 4°C) se realizó un corte de la biomasa área a 10 cm de altura desde el nivel del suelo. Luego de 160 días, se realizó el descalzado de las macetas y se registró la biomasa fresca total (BFT) por gravimetría. Posteriormente, las plantas fueron fraccionadas, separando la parte aérea de la subterránea. Se determinó la biomasa fresca aérea (BFA) y radical (BFR). Para ello, las raíces se separaron del sustrato, lavándolas y secándolas con papel absorbente antes de pesarse. La fracción aérea, se colocó en bolsas de papel rotuladas para secarse en estufa a 60°C hasta peso constante (PSA). Luego de la cuantificación del BFR, las raíces fueron procesadas para la tinción de acuerdo a INRA (2001) y posteriormente se procedió a la cuantificación de la colonización micorrícica de acuerdo a Trouvelot et al. (1986, descripto en INRA, 2001).
Análisis estadístico
Se ajustaron modelos lineales que consideraron los efectos principales y de interacción entre los factores experimentales: nivel de infección endofítica, nivel inoculación con HMA y nivel de fertilización fosfatada. Previamente al análisis, realizó la verificación del cumplimiento de los supuestos del ANOVA (normalidad de los residuos y la homogeneidad de varianzas). Las medias se compararon con la prueba de la Mínima Diferencia Significativa (MDS); se consideró significativa toda diferencia con un valor de probabilidad p<0,05. Se empleó el software estadístico R (R Development Core Team, 2018).
RESULTADOS
Al analizar la biomasa fresca total y su partición en aérea y radical, no se halló interacción triple significativa entre los factores experimentales (Tabla 1) pero sí interacciones dobles significativas (Tabla 1) entre el nivel de infección endofítica y el nivel de inoculación con HMA, y entre el nivel de inoculación con HMA y el nivel de fertilización con P. Por un lado, no hubo diferencia significativa entre la producción de biomasa de las plantas HMA+ y HMA- cuando estas eran SE+, pero si, cuando eran SE-, donde las plantas HMA+ produjeron menor BFT, BFA y BFR que las HMA- (Tabla 2). Por otro lado, no se detectaron diferencias significativas en la producción de biomasa entre plantas HMA+ y HMA- cuando fueron fertilizadas (P15) (Tabla 2). Sin embargo, cuando las plantas no recibieron P (P0), las plantas HMA+ produjeron menor BFT, BFA y BFA que las HMA- (Tabla 2).
Al analizar el PSA, no se halló interacción triple significativa entre los factores evaluados (Tabla 1); aunque sí fue significativa la interacción entre el nivel de inoculación con HMA y el nivel de fertilización con P (Tabla 1). También se registró efecto simple del nivel de infección endofítica (Tabla 1). Por un lado, las plantas HMA- presentaron mayor PSA que las HMA+, en el tratamiento sin agregado de P (Tabla 2). Por otro, no se registraron diferencias de PSA entre plantas HMA- y HMA+ en los tratamientos P15 (Tabla 2). Las plantas SE- presentaron mayor PSA que las SE+ (Tabla 2).
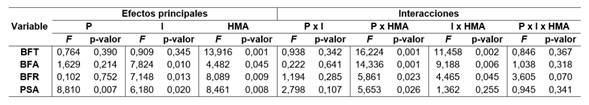
Tabla 1 Efectos principales e interacciones (ANOVA) entre los factores experimentales: fertilización con fósforo (P), infección endofítica (I), inoculación con hongos micorrícos arbusculares (HMA), para las variables respuesta evaluadas: biomasa fresca total (BFT) y su partición en aérea (BFA) y radical (BFS) y peso seco aéreo (PSA). Table 1. Main effects and interactions (ANOVA) between the experimental factors: fertilization with phosphorus (P), endophytic infection (I), inoculation with arbuscular mycorrhizal fungi (HMA), for the response variables evaluated: total fresh biomass (BFT) and its partitioning into aboveground (BFA) and belowground (BFR) and air dry weight (PSA).
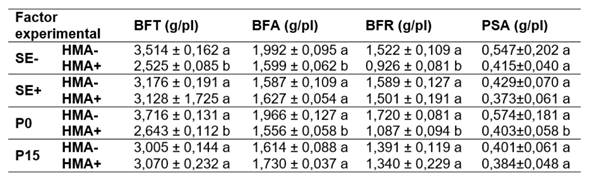
Tabla 2 Biomasa fresca total (BFT) y su partición en aérea (BFA) y radical (BFS) y peso seco aéreo (PSA) (media ± EE; g/planta; n=5). SE+ = con endófito silvestre; SE-= libre de endófito; HMA+ = con agregado de hongos micorrícicos arbusculares (HMA); HMA- = sin inoculación de HMA; P0 = sin agregado de P; P15 = con agregado de 15 ppm de P. Letras distintas por columna para cada combinación de factores experimentales indican diferencias significativas (p<0,05). Table 2. Total fresh biomass (BFT) and its partitioning into aboveground (BFA) and belowground (BFS) and air dry weight (PSA) (mean ± ES; g/plant; n=5). SE+ = with wild endophyte; SE- = free of endophyte; HMA+ = with added arbuscular my-corrhizal fungi; HMA- = without HMA inoculation; P0 = without added P; P15 = with added 15 ppm P. Different letters per column for each combination of experimental factors indicate significant differences (p<0.05).
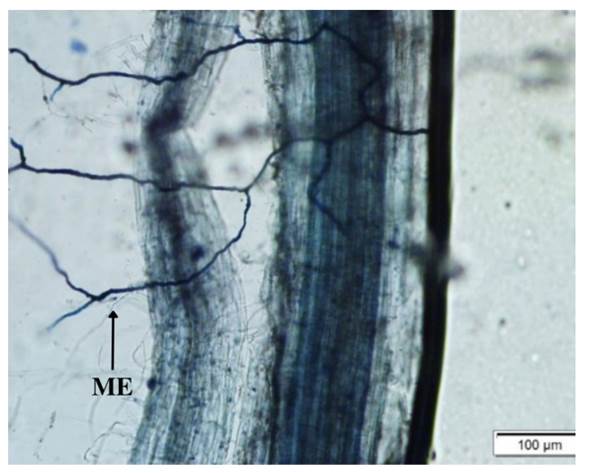
No se detectó evidencia de colonización micorrícica en las raíces de las plantas no inoculadas con HMA, corroborando la erradicación de HMA antes de la inoculación. Contrariamente a lo esperado, no se registró colonización intrarradical en las raíces de las plantas inoculadas con HMA, no evidenciándose arbúsculos ni vesículas. En este sentido, la colonización de raíces por HMA en todas las situaciones evaluadas fue del 0%. Solo se detectó micelio extraradical (Figura 1) en tres de las cinco repeticiones para las plantas SE+ HMA+ crecidas bajo la condición de P0 en una intensidad del 10%, 15% y 13%, lo que resultó en un grado de presencia de micelio extraradical para dicha condición del 7,6% (desvío estándar 7,2%). Para la condición de P15 en plantas SE- HMA+ se registró presencia de micelio extraradical solo en una repetición (intensidad de micelio extraradical 14%), lo que correspondió a una intensidad total del micelio externo del 2,8% (desvío estándar 6,3%).
DISCUSIÓN
Bajo las condiciones del presente experimento, no se detectaron efectos de presencia de los endófitos E. coenophiala (SE+) y de la inoculación con HMA para ninguno de los niveles de fertilización con P en la producción de BFT y su partición en las plantas de festuca alta (Tabla 1). Estos resultados no permitirían aceptar la hipótesis planteada. Era de esperar que una asociación simbiótica como la de HMA con las raíces de las plantas vasculares, fuese promovida o al menos tolerada por los endófitos Epichloë, dado que, si bien los endófitos y los HMA obtienen los mismos beneficios de su hospedante (i.e. hidratos de carbono), difieren en su ubicación y en el momento de la colonización (Vignale, 2016). Los endófitos Epichloë sp. están localizados en los tejidos foliares (Clay & Schardl, 2002), es decir, en donde se lleva a cabo la fotosíntesis, es por ello que, acceden primero a los fotosintatos; mientras que, los HMA colonizan las raíces de sus hospedantes de manera “horizontal”, después de que las semillas germinan (Vignale, 2016).
Por otro lado, si bien en este estudio no se determinó el contenido de P en el sustrato, es esperable que éste haya sido suficiente dado que no se detectaron efectos en la producción de biomasa por la fertilización fosfatada (Tabla 2). Esto haría suponer que los niveles de P en el sustrato, tanto para la situación P0 como para la P15 podrían haber sido elevados, situación que podría haber conspirado contra la formación de micorrizas en las raíces de las plantas. Covacevich et al. (2006), observaron que la colonización espontánea por HMA en festuca alta fue deprimida por la fertilización con fuentes de P solubles. Por otra parte, Covacevich & Echeverría (2009) determinaron efectos positivos en la producción de biomasa de festuca alta por la inoculación con un aislamiento no nativo (Glomus claroideum) de HMA cuando las poblaciones nativas habían sido eliminadas con formaldehído (tal como en este estudio), particularmente oferta de roca fosfórica o cuando las plantas superfosfato triple de calcio; aunque no se detectaron respuestas positivas cuando el aislamiento consistió en propágulos de HMA de la especie Acaulospora longula. En este estudio, la inoculación se realizó con un consorcio de esporas de HMA nativas y, en base a la metodología utilizada, no se detectó colonización micorrícica (i.e., arbúsculos e hifas intraradicales) ni tampoco efecto alguno por la inoculación. Esto indicaría que las plantas hospedantes de esta población de festuca alta, independientemente de su nivel de infección endofítica, fueron capaces de cubrir sus requerimientos de absorción de nutrientes independientemente de la formación de simbiosis micorrícica.
La potencialidad de los HMA en favorecer la microbiota y estructura del suelo, es indiscutible (Gupta, 2020). Sin embargo, la potencialidad de los HMA en favorecer la nutrición y/o crecimiento de plantas, podría depender no solamente de la interacción entre el endófito E. coenophiala y la especie de HMA involucrada, sino también, del genotipo de la planta hospedante, debido a los resultados contrastantes hallados en la literatura (e.g., Antunes et al., 2008; Novas et al., 2005). Cabe desatacar que, en todas las condiciones evaluadas se registró micelio extraradical, sin la formación de arbúsculos, vesículas, ni observación de micelio intraradical, estructuras característica que intervienen durante la simbiosis micorrícica entre las plantas y los HMA (Bagó et al., 2000). El micelio extraradical detectado pero sin formación de arbúsculos, indicaría que la inoculación se realizó con esporas de HMA viables. Futuros estudios deberán evaluar la respuesta de otros genotipos de festuca alta infectados con el hongo endófito E. coenophiala al ser inoculados con diferentes consorcios de HMA. Además, se deberán considerar otras variables que influyen sobre la respuesta a la colonización micorrícica, como son el tipo de suelo, el estado de desarrollo de la planta hospedante y la disponibilidad de P en el suelo y contenido en planta (Smith & Read, 2008). Sería importante, además, considerar un mayor periodo de evaluación debido a los tiempos que poseen algunas especies de HMA para completar su ciclo de colonización e investigar mecanismos que podrían explicar la influencia de los hongos endófitos sobre la colonización de las plantas por HMA, tales como son la lixiviación y exudación de compuestos químicos producidos por los endófitos (Antunes et al., 2008; Casas et al., 2011; Vignale, 2016).
CONCLUSIONES
En este estudio, no se registraron las respuestas esperadas en producción de biomasa en plantas infectadas con el hongo endófito y los HMA ni sinergismo entre ambos simbiontes, durante el periodo de tiempo contemplado. Además, no se evidenció efecto modulador de la fertilización fosfatada en las respuestas esperadas. Son muy escasos los reportes acerca de la interacción entre el hongo endófito E. coenophiala asociados a F. arundinacea y los HMA y la modulación por la fertilización fosfatada, que nos permita explicar el motivo por el cual no se registró colonización micorrícica en las plantas de esta población de festuca alta. En este sentido, aun con falta de respuesta, los resultados de este estudio constituyen un aporte original sobre la asociación de dos endosimbiontes en festuca alta ante ofertas variables de P. De este modo, en futuros estudios se deberá evaluar la respuesta de otros genotipos de festuca alta infectados con el hongo endófito E. coenophiala a la inoculación con diferentes consorcios de HMA, considerando además un mayor periodo experimental y otras variables que influyen sobre la respuesta a la colonización micorrícica.

