











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
La autolocomoción restringida expone a las plantas a condiciones que afectan la homeostasis celular y les provocan estrés. El estrés es el estado subóptimo causado por factores abióticos (e.g., temperatura) y bióticos (e.g., parasitismo) que actúan en forma aislada, simultánea o sucesiva (Mickelbart et al., 2015; Mosa et al., 2017). Los efectos del estrés pueden ser interactivos y sinérgicos, y varían en forma inter- e intraespecífica de acuerdo al tipo, la cantidad, intensidad, duración, recurrencia y sincronización con respecto al ciclo diario y anual de los factores causales, y la edad y el estado de la planta (Fowden et al., 1993; Verlues et al., 2006; Claeys & Inzé, 2013).
El estrés promueve cambios morfológicos, fisiológicos y comportamentales que promueven la adaptación y la evolución biológica (Bijlsma & Loeschcke, 1997; Calowa & Forbes, 1998; Badyaev, 2005; Sorensen & Loeschcke, 2007; Bita & Gerats, 2013; Mickelbart et al., 2015). Estos cambios pueden ser similares en diferentes especies que se desarrollan en ambientes equivalentes, a causa del origen monofilético o la convergencia evolutiva (Mickelbart et al., 2015). Las plantas también pueden exhibir atributos originados a través de la exaptación evolutiva, que representan ventajas frente a factores múltiples como la sequía y herbivoría (Milchunas et al., 1988; Armbruster, 1997). Los efectos del estrés pueden involucrar respuestas de tolerancia, evasión o escape (Badyaev, 2005; Gusta & Wisniewski, 2013; Mickelbart et al., 2015). La extinción de una población o especie suele ser el resultado de factores de estrés múltiples e interactivos (Kristensen et al., 2018).
El clima de una región incluye factores que pueden provocar estrés y promover adaptaciones específicas en las plantas (Bita & Gerats, 2013; Claeys & Inzé, 2013; Bonan, 2015; Pessarakli et al., 2015). En las regiones extratropical y de montaña, los factores de estrés climáticos tienen un impacto mayor que los bióticos debido a la mayor variabilidad e imprevisibilidad (Bijlsma & Loeschcke, 1997; Kreyling, 2010). Las plantas de estos ambientes experimentan temperaturas bajas en forma crónica (e.g., congelamiento de invierno) y aguda (e.g., helada intensa de verano). Los mecanismos de evasión incluyen la protección del tejido meristemático que retoma la multiplicación celular a partir de la primavera (Larcher, 2003; Baxter, 2014). La clasificación de las “formas de vida” (1) se basa en la posición y protección de la yema meristemática apical del tallo relativa a la superficie del suelo, que representa una estrategia de evasión durante la estación climática desfavorable (Box, 2019). Entre las categorías de esta clasificación se encuentran las hemicriptófitas (yemas superficiales), geófitas (yemas subterráneas), caméfitas (yemas cerca del suelo), fanerófitas (yemas a mayor altura; nanofanerófitas con 0,5 - 2 m de altura de la planta, y mesofanerófitas con 5 - 50 m) y terófitas (sin yemas).
La posición de la yema predice otras respuestas de la planta al ambiente, porque está relacionada con la altura del tallo, las características de la hoja, la fenología, la productividad primaria y las estrategias frente a perturbaciones (e.g., capacidad de rebrote a través de órganos subterráneos) (Lavorel & Garnier, 2002; Ott et al. 2019). Esta clasificación se diseñó inicialmente para interpretar la posición de los meristemas inactivos como una respuesta de la planta a los periodos desfavorables de los climas templados. Posteriormente el concepto fue extendido a los trópicos (Sarmiento & Monasterio, 1983; Irl et al., 2020). Los rasgos de las formas de vida constituyen un antecedente de los “tipos funcionales de las plantas”, utilizados en los modelos globales de clasificación y dinámica de la vegetación (Harrison et al., 2010; Baxter, 2014; Box, 2019).
El ecotono es un área de transición de ecosistemas naturales o antropogénicos adyacentes y divergentes, donde la variación ambiental conforma un gradiente intenso (Kark, 2017). En la transición andina - extraandina de la Patagonia argentina, el patrón de circulación atmosférica del Último Máximo Glacial impulsó el desarrollo de un marcado ecotono bosque - estepa, que está modelado por el régimen moderno de disturbios geológicos y humanos (Huber et al. , 2004). Este ecotono es una región de altitud intermedia, templada y ventosa, donde ocurre una marcada transición de humedad, y con influencias mediterránea, asociada a veranos secos e inviernos lluviosos, y continental, relacionada con grandes amplitudes térmicas diaria y estacional (Prohaska, 1976; Jobbágy et al., 1995; Paruelo et al., 1998;
Garreaud et al., 2013). En esta región, durante los últimos 50 años la temperatura aumentó 0,4 °C y la cantidad de días con helada se redujo 50%, y durante el s. XXI se proyecta un aumento de la temperatura entre 0,5 y 1 °C y una disminución de la precipitación de hasta 10% (Barros & Vera, 2015; IPCC, 2019). Se prevé que el invierno en las latitudes altas sea afectado de manera más significativa por el cambio climático (Kreyling, 2010).
En este ecotono, el déficit hídrico estival, resultante de la escasa precipitación y las altas temperatura e intensidad y frecuencia del viento, ejerce efectos marcados en la vegetación (Paruelo et al., 2001, 2004), y la adaptación de las plantas se ha asociado principalmente a la aridez (Cabrera, 1976; Golluscio et al., 1982; Soriano & Sala, 1983; Sala et al., 1989; Aguiar et al., 1996; Lauenroth, 1998; León et al., 1998; Ghermandi et al., 2004; Villagra et al., 2011; Oyarzábal et al., 2018; Matteucci et al., 2019). Sin embargo, las bajas temperaturas constituirían otro factor de estrés crónico y agudo, con manifestaciones en la forma y fenología de las plantas. El objetivo de este trabajo fue investigar el clima y las adaptaciones al frío en los diferentes tipos de vegetación que se desarrollan en un área de la transición andina - extraandina en el Norte de la Patagonia argentina, basado particularmente en el análisis de las formas de vida de las plantas vasculares.
En este trabajo, se hipotetiza que las plantas dominantes en composición y abundancia en el área de estudio son las que exhiben formas de vida compatibles con la adaptación al frío, independientemente de la variabilidad de la vegetación asociada al intenso gradiente ambiental. En particular se prevé la mayor abundancia de hemicriptófitas y la presencia de geófitas y terófitas, cuyos comportamientos estructural y fenológico están asociados a las condiciones invernales frías de la región. Las plantas constituyen un grupo focal clave para monitorear el cambio ambiental, el estado de los ecosistemas y el uso de la tierra. La comprensión de las relaciones clima - vegetación es particularmente importante en el contexto actual de impacto humano intenso, frecuente y de gran escala, asociado a la destrucción y fragmentación de los ecosistemas naturales, la contaminación química, la pérdida de diversidad biológica y el calentamiento global.
Materiales y Métodos
El área de estudio comprendió 1780 ha distribuidas en los campos Aguas Frías (38° 46' S, 70° 54' O) y Litrán (38° 54' S, 71° 01' O) (Fig. 1). La distancia en línea recta entre estas estaciones es 19 km. La geología de base corresponde a basaltos, andesitas, brechas, aglomerados volcánicos y depósitos glaciales no estratificados. Estos materiales están cubiertos por una capa de tefra Holocénica aportada por los volcanes activos de la cordillera de los Andes, que conforma el material parental de los suelos. La geomorfología consiste en montañas y valles con signos de glaciación y erosión, como lagos, circos, valles colgados y bloques erráticos. Los suelos dominantes corresponden a Andisoles Udivitrands y Endoaquands alofánicos. Aguas Frías presenta arroyos permanentes y temporarios cuyas aguas se desplazan en dirección Norte - Sur (Frugoni et al., 2016).
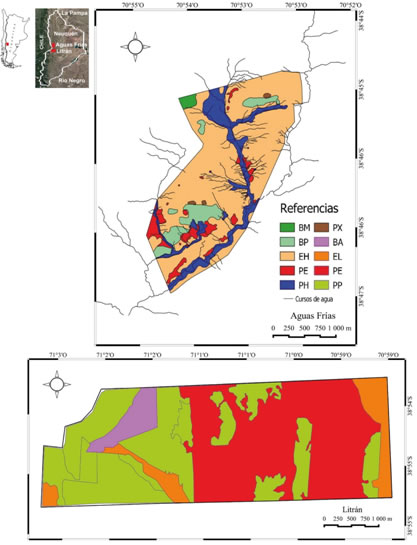
Fig. 1: Mapa de vegetación de Aguas Frías y Litrán localizados en el centro - oeste de la provincia de Neuquén en la Argentina. BA: bosque de A. araucana, BM: bosque de N. pumilio y A. araucana, BP: bosque de N. pumilio, EH: estepa herbáceo arbustiva, EL: estepa herbácea, PE: peladero, PH: pradera higrófila, PP: plantación de pino, PX: pradera xerófila.



La vegetación zonal es un mosaico de bosques altos y cerrados de latifoliadas caducifolias y coniferas, y pastizales y matorrales de menor cobertura aérea. La vegetación azonal e intrazonal está formada por praderas (León et al., 1998; Cingolani et al., 2000; Oyarzábal et al., 2018; Matteucci et al., 2019) (Fig. 1). Los disturbios están asociados a los incendios naturales e inducidos, y al pastoreo de herbívoros nativos e introducidos (Munist & Speck, 1982; Lauenroth, 1998; Gaitán et al., 2009). La región forma parte de una ruta histórica de trashumancia de subsistencia, caracterizada por el desplazamiento del ganado desde los valles en invierno y hasta las laderas elevadas en verano (Frugoni et al., 2016).
La caracterización del clima se basó en información sobre los valores medios mensuales de la precipitación (P m ), temperatura (T m ), humedad relativa (HR), y velocidad (V v ) y dirección del viento (V d ), colectada en las estaciones meteorológicas Litrán Abajo (38° 45’ S, 70° 52’ O) y Salida Lago Aluminé (38° 58’ S, 71° 03’ O) (periodo 1999 - 2019; AIC, 2020). Litrán Abajo se utilizó para describir el clima de Aguas Frías (distancia 2,9 km) y Salida Lago Aluminé el de Litrán (7,4 km). El clima se clasificó de acuerdo a Koppen & Geiger (1936), a partir de la temperatura media de los meses más cálido (T) y más frío (Tf), la precipitación media de los meses más seco de verano (P) y más húmedo de invierno (P/3), y la cantidad de meses con temperatura mayor a 10 °C (T 10 ). Para cada estación se determinaron la evapotranspiración potencial (ETp) y el balance hídrico climático (BH) mensuales, basado en la existencia de periodos
En una imagen satelital Aster (resolución 15 m, coordenadas Gauss Krüger banda 1, elipsoide WGS 1984), las unidades de vegetación se clasificaron en forma fisonómico - florística y se elaboraron mapas de vegetación. Esta clasificación se basó en la composición, el tamaño y la forma de crecimiento de las especies de plantas dominantes de acuerdo a la cobertura aérea y biomasa, y luego esta clasificación se verificó en el terreno. En cada unidad se localizaron sistemáticamente entre 10 y 30 estaciones de muestreo, a las cuales se les registró la posición geográfica (mediante un posicionador satelital), la altitud (con altímetro), la pendiente (con clinómetro) y la exposición (con brújula).
Desde el centro de cada estación se establecieron cuatro transectas de 10 m a lo largo de las direcciones Norte, Sur, Este y Oeste. En cada transecta, se localizó cada 1 m un punto de muestreo para determinar la presencia de plantas, utilizando el método del punto de intercepción (n = 5200) (Kent, 2011). La frecuencia basada en datos de incidencia presencia / ausencia permite estimar la abundancia de plantas cuando existe la imposibilidad de establecer el límite físico de cada individuo (Jost et al., 2011). Las especies se clasificaron de acuerdo a la forma de crecimiento (hierba, arbusto, árbol), el origen (nativa, exótica) y la forma de vida (terófita, geófita, hemicriptófita, caméfita, nanofanerófita, mesofanerófita), de acuerdo a las bases de datos de IBD (2021) y PLANEAR (2021).
Para cada unidad se determinaron la cantidad total de especies (S) y de especies únicas (SJ y raras (R5%). Representó las especies presentes en una sola unidad y R5% aquellas cuya frecuencia fue menor a 5% (McGill, 2011), según:
La similitud entre unidades de vegetación se basó en la composición (Sorensen, 1948) y la frecuencia relativa de especies para N y > 2 (Morisita, 1959; Hom, 1966), pp™11'
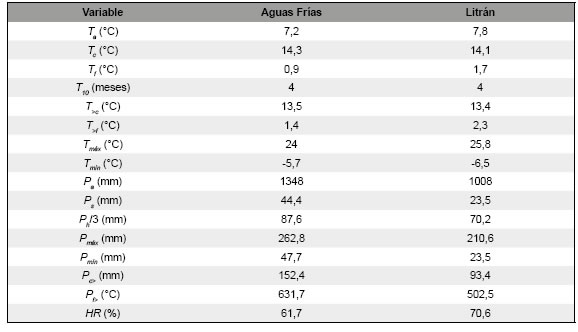
Tabla 1: Características climáticas de Aguas Frías y Litrán. Ta: temperatura media anual, Tc: temperatura media del mes más cálido, Tf: temperatura media del mes más frío, T10: cantidad de meses Tm > 10 °C, Tm: temperatura media mensual, T>c: temperatura media de los meses más cálidos, T>f: temperatura media de los meses más fríos, Tmáx: temperatura media mensual máxima, Tmín: temperatura media mensual mínima, Pa: precipitación total media anual, Ps: precipitación total media del mes más seco, Ph: precipitación total media del mes más húmedo, Pmáx: precipitación total media mensual máxima, Pmín: precipitación total media mensual mínima, Pc>: precipitación total de los meses cálidos, Pf>: precipitación total de los meses fríos, HR: humedad relativa media mensual.
La estructura de las formas de vida se estableció determinando los espectros biológicos florístico ( EBF ) y vegetacional ( EBV ). EBF representó el número de especies de cada forma de vida y en consecuencia, cada especie contribuyó de la misma forma, mientras que EBV consideró la abundancia de cada especie y, consecuentemente, cada una contribuyó al espectro en función de la frecuencia.
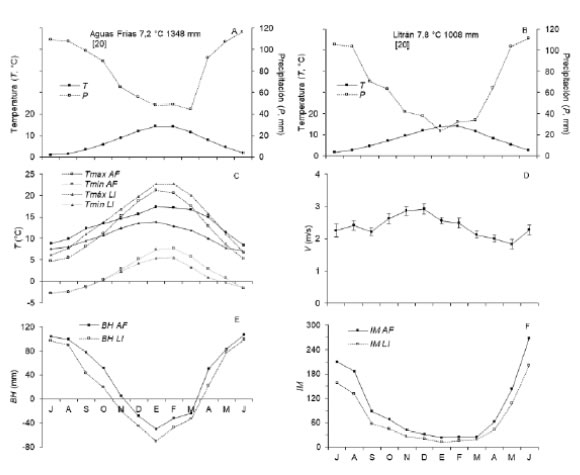
Fig. 2: Diagrama climático (A, B), temperatura máxima (Tmáx), mínima (Tmín) y diferencia de temperatura (ΔT) (C), intensidad del viento (V) (D), balance hídrico climático (BH) (E), e índice de aridez (IM) (F) en Aguas Frías (AF) y Litrán (LI). Las barras verticales indican el error estándar de la media (n = 20).
Resultados
Clima
El clima de Aguas Frías es templado húmedo y frío (Cfb, Peel et al., 2007) (Tabla 1). La temperatura media anual T a = 7,2 °C y la precipitación total media anual P a = 1348 mm (Fig. 2A). La diferencia de la temperatura media entre los meses cálidos (diciembre, enero, febrero) y fríos (junio, julio, agosto) AT cf = 12,1 °C. La diferencia de la temperatura máxima y mínima media de los meses cálidos y fríos AT m = 29,7 °C. Entre junio y setiembre, T . < 0 °C (junio: -1,7 °C, julio: -2,9 °C, agosto: -2,6 °C y setiembre: -1,2 °C). El coeficiente de variación medio de la temperatura mensual CV t = 33,3% (Fig. 2C). La diferencia de precipitación media entre los meses cálidos y fríos AP = 479,4 mm. El coeficiente de variación medio de la precipitación mensual CV p = 67,5%. Entre diciembre y febrero se registró P m = 0 mm y en mayo de 2005 se registró la precipitación mensual máxima absoluta P = 460,0 mm. CV = 5,6%, BH varió entre -24,2 mm (marzo) y 106,7 mm (junio), y se produjo déficit hídrico entre diciembre y marzo (Fig. 2E). I varió entre 23,5 (enero) y registros exhibió V v = 1,5 - 3 m/s y El 94,5% una dirección del viento S - SO y SO - O (n = 163) (Fig. 2D). CV hr = 4,3%, BH varió entre -70,4 mm (enero) y 98,6 mm (junio), y se produjo déficit hídrico entre noviembre y marzo (Fig. 2E). I M varió entre 11,8 (enero) y 200,3 (junio); entre abril y octubre I M > 40 (húmedo, n = 7), noviembre y diciembre I M = 20 - 30 (semiárido, n = 2) y entre enero y marzo I M = 10 - 20 (árido, n = 3) (Fig. 2F).
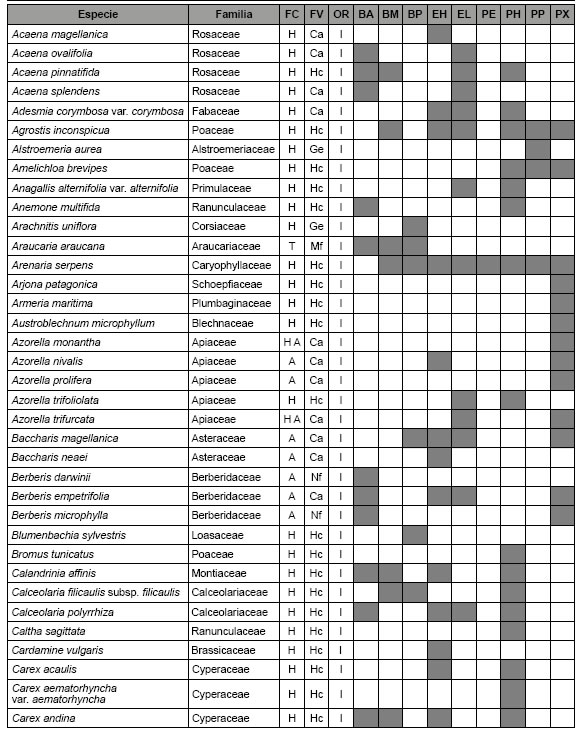
Tab 2: Características de las plantas vasculares de Aguas Frías y Litrán. FC: forma de crecimiento, A: arbusto, H: hierba, T: árbol, FV: forma de vida, Ca: caméfita, Ge: geófita, Hc: hemicriptófita, Mf: mesofanerófita, Nf: nanofanerófita, Te: terófita, OR: origen, E: exótica, I: nativa, BA: bosque de A. araucana, BM: bosque de N. pumilio y A. araucana, BP: bosque de N. pumilio, EH: estepa herbáceo arbustiva, EL: estepa herbácea, PE: peladero, PH: pradera higrófila, PP: plantación de pino, PX: pradera xerófila.
Vegetación
La vegetación forma parte de la transición entre los Distritos Subandino y Occidental de la Provincia Patagónica (Región Neotropical, Dominio Andino Patagónico) y el Distrito del Pehuén de la Provincia Subantártica (Región
Antártica, Dominio Subantártico) (Cabrera, 1976; Oyarzábal et al., 2018). Las unidades de vegetación fueron el bosque puro de Nothofagus pumilio (BP; 2,0% del área de estudio), el bosque mixto de N. pumilio y Araucaria araucana (BM; 0,3%), el bosque abierto de A. araucana (BA; 3,5%), la plantación de Pinus ponderosa (PP; 30,4%), la estepa herbáceo arbustiva (EH; 18,3%), la estepa herbácea (EL; 6,5%), la pradera higrófila (PH; 4,0%), la pradera xerófila (PX; 0,3%), y el peladero (PE; 34,7%) (Fig. 1).
En estas unidades de vegetación se identificaron 148 especies de plantas, de las cuales PE (S = 22) y PH ( S = 68) exhibieron la menor y mayor riqueza, respectivamente (Tabla 2). El 42,7% de las especies estuvo presente en una sola unidad de vegetación (Su = 64) y el 2,7% de las especies fue cosmopolita (C = 4), de las cuáles una fue nativa ( Poa obvallata) y tres exóticas (Cerastium arvense, Rumex acetosella y Taraxacum officinale). En cada unidad, S u varió entre 0 (BM) y 19 (PH), y la relación S u :S varió entre 0 (BM) y 27,9% (PH). En PP, S = 9, mientras que en EL, S = 2 y en EH, S = 11 (Fig. 3). La similitud global entre unidades
de veg etación CS Nv>2) = 0,67 y CM (N v>2) = 0,11
El 69,4% de las unidades exhibieron CS entre 0,21 y 0,40, mientras que el 27,8% fue entre 0,41 y 0,60. El 61,1% de las unidades exhibieron una similitud CM (Nv = 2) entre 0 y 0,20, mientras que sólo dos comparaciones tuvieron una similitud mayor a 0,61 (BM - BP, CM m „ = 0,834; EH - PX, CM m „
(Nv=2) (Nv=2)
= 0,778). CS N¡v = 2) varió entre 0,24 (BA - BP) y 0,78 (EH - PH). Entre las unidades de bosque, CS varió entre 0,24 (BA - BP) y 0,56 (BM - BP) y entre las de pradera PX - PH fue CS (Nv = 2) = 0,33. La mayoría de las comparaciones exhibieron CM (NJv = 2) < 0,10. Entre las unidades de bosque, CM (NJv = 2) varió entre 0,038 (BA - BP) y 0,834 y entre las unidades de pradera PX - PH, CM (Nv=2) = 0,23 (. 4).
En el área de estudio, la cantidad de especies que exhibió P = 0 - 4,9% fue 141 (95,3%), P = 5 - 9,9% fue 4 (2,7%) y P = 10 - 14,9% fue 3 (2,0%); ninguna especie presentó P > 15%. El valor mínimo de R 5 % fue 0,806 (BM) y el máximo fue 1 (PE). En cada unidad, entre 83,9% (BM, S = 26) y 100% de las especies (PE, S = 22) presentó P < 10%. Sólo una especie alcanzó P = 50 - 59,9% (en PP, P ponderosa) y sólo dos especies alcanzaron P = 40 - 49,9% (en BA, A. araucana y en BP, N. pumilio) (Fig. 1 A). El índice de dominancia global C McN = 10,9 y el valor mínimo fue 11,9 (PH); PP (CMcN = 35,3) presentó el valor máximo de dominancia (Fig. 2 B).
Las especies que exhibieron la mayor frecuencia en cada unidad fueron A. araucana (46,1%; BA), N. pumilio (38,9%; BM y 47,0%; BP), Festuca pallescens (37,5%; EH y 25,2; PX), Festuca
acanthophylla (36,4%; EL), Chusquea culeou (4,3; PE), Cortaderia egmontiana (26,8; PH) y P ponderosa (57,9; PP). En el bosque los árboles presentaron la mayor frecuencia, en la estepa y pradera las herbáceas, y en el peladero las arbustivas. La cantidad total de familias fue 49; la cantidad de familias en las unidades de vegetación varió entre 9 (PE) y 24 (PP). Las familias más diversas fueron Asteraceae (S = 26; 17,3% de las especies) y Poaceae (S = 23; 15,3%), mientras que 23 familias (46,9%) sólo contaron con una especie. Poaceae fue la familia más frecuente en 7 de las 9 unidades de vegetación, mientras que en PP fue la segunda más frecuente (24,6%), y en BA la tercera (11,6%).
Atributos de las plantas
La cantidad de especies herbáceas fue 115 (77,7%), herbáceas o arbustivas 5 (según el tipo de vegetación; 3,4%), arbustivas 23 (15,5%), arbóreas o arbustivas 1 (según el tipo de vegetación, Discaria chacaye, 0,7%) y arbóreas 4 (P. ponderosa, A. araucana, N. pumilio y Nothofagus antarctica ; 2,7%). El 24,7% de las especies fueron graminoideas (gramíneas, ciperáceas y poáceas; S = 36), y el 15,3% fueron gramíneas (S = 22). La frecuencia de especies herbáceas fue 58,3%, herbáceas o arbustivas 1,6%, arbustivas 13,6%, arbóreas o arbustivas <0,1% y arbóreas 16,1%. La frecuencia de gramíneas fue 64,6% y de graminoideas 70,7%. El 93,9% de las especies fue nativa (S = 139); las exóticas fueron Cerastium rvense , Epilobium brachycarpum , Melilotus albus , P. ponderosa , Poa pratensis subsp . pratensis , Rumex acetosella, Taraxacum officinale, Trifolium repens y Veronica serpyllifolia. Las especies nativas representaron el 77,1% de la frecuencia. La cantidad de especies herbáceas varió entre 61,7% (PX) y 91,2% (PH) (Fig. 6 A), y la de nativas entre 83,9% (BM) y 93,6% (PX) (Fig. 6 B). La frecuencia de especies herbáceas varió entre 30,9% (BM) y 96,3% (EL), mientras que la de árboles fue 31,0% en BP, 45,5% en BA, 49,3% en BM y 54,5% en PP (Fig. 6 ). La frecuencia de especies nativas varió entre 37,2% (PP) y 98,7% (PX) (Fig. 6 D).
El EBF estuvo dominado por las hemicriptófitas (S = 94; 63,5%) y caméfitas (S = 21; 14,2%), mientras que las geófitas, nanofanerófitas, terófitas y mesofanerófitas fueron 11 (7,4%), 11 (7,4%), 7 (4,7%) y 4 (2,7%) especies, respectivamente. La cantidad de hemicriptófitas fue mayor en todas las unidades de vegetación, y varió entre 59,6% (PX) y 86,6% (PH). La cantidad de caméfitas varió entre 3,0% (PH) y 29,8% (PX), la de geófitas entre 0% (BM, BA, PX) y 12,5% (BP), y la de terófitas entre 0% (BP, PX) y 9,7% (BM). Las nanofanerófitas fueron importantes en PX (10,6%), BA (11,5%) y PE (13,0%) (Fig. 7A). El EBV estuvo dominado por las hemicriptófitas (52,7%) y mesofanerófitas (26,9%), mientras que las nanofanerófitas, caméfitas, geófitas y terófitas representaron el 11,3%, 6,0%, 2,0% y 1,3%, respectivamente. La frecuencia de hemicriptófitas fue mayor en todas las unidades de vegetación, y varió entre 30,2% (BM) y 80,8% (EL), y la de mesofanerófitas fue mayor en BP (31,0%), BA (45,5%), BM (49,3%) y PP (54,5%). La frecuencia de caméfitas varió entre 0% (BM) y 26,1% (PX), la de geófitas entre 0% (BM y BA) y 10,3% (PH) y
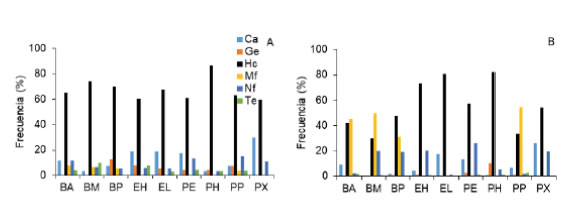
Fig. 7: Espectro biológico florístico (A) y vegetacional (B) en las unidades de vegetación de Aguas Frías y Litrán. Ca: caméfita, Ge: geófita, Hc: hemicriptófita, Mf: mesofanerófita, Nf: nanofanerófita, Te: terófita, BA: bosque de A. araucana, BM: bosque de N. pumilio y A. araucana, BP: bosque de N. pumilio, EH: estepa herbáceo arbustiva, EL: estepa herbácea, PE: peladero, PH: pradera higrófila, PP: plantación de pino, PX: pradera xerófila
Discusión
En el área de estudio, la latitud, la cordillera de los Andes, la corriente de Humboldt, la temperatura relativa del océano y el Sistema de Alta Presión del Pacífico determinan un clima templado ventoso, con inviernos fríos y húmedos y veranos cálidos y secos, y con amplitud térmica estacional mayor a 10 °C que aumenta hacia el Este. Durante la estación cálida, la intensidad de la radiación solar aumenta y la humedad relativa, la nubosidad y la precipitación disminuyen, y aumenta la demanda evaporativa de la atmósfera (Prohaska, 1976; Paruelo et al., 1998; Nicholson, 2011; Sagredo & Lowell, 2012; Garreaud et al., 2013). El aumento de la altitud provoca la disminución de la temperatura, y el aumento de las nevadas y heladas, la humedad relativa y el viento (Paruelo et al., 1998; Garreaud et al., 2013). A su vez, las laderas de exposición Sur reciben la menor cantidad de radiación, que disminuye a medida que aumenta la pendiente del terreno (Bonan, 2015).
Este patrón climático general es compatible con la información de las estaciones de muestreo. Aguas Frías (Cfb) y Litrán (Csb) exhiben un clima templado frío, con la temperatura del mes más cálido igual o mayor a 10 °C, y con la temperatura del mes más frío entre -3 y 18 °C, y la de cada uno de los cuatro meses más cálidos igual o mayor a 10 °C y menor a 22 °C. El clima frío del área de estudio está asociado a la presencia de temperaturas mínimas medias mensuales por debajo del punto de congelación durante 4 ó 5 meses del año. Sin embargo, Aguas Frías presenta un clima más húmedo y con la precipitación distribuida más uniformemente a lo largo del año, mientras que Litrán presenta un clima con verano seco, y con la precipitación del mes más seco de la mitad cálida del año, menor a 30 mm y menor a 1/3 de la precipitación del mes más húmedo de la mitad fría del año.
La diferente distancia de los campos al límite de altas cumbres de la cordillera de los Andes promovería en Aguas Frías (distancia ~ 4 km) un clima más frío y húmedo que en Litrán (~ 33 km), y con una distribución más uniforme de la precipitación, vinculado a la ausencia de estación de sequía relativa, mayores precipitaciones totales, máxima, mínima y de los meses más cálidos y fríos, y menores temperaturas medias, máximas, mínimas y de los meses fríos, y menor amplitud térmica, aridez y evapotranspiración potencial (Jobbágy et al., 1995). Este fenómeno está asociado a la cordillera de los Andes, que provoca cambios adiabáticos del aire húmedo que proviene del Oeste, y cuya subsidencia causa el descenso exponencial intenso de la precipitación a medida que aumenta la distancia de la cordillera hacia el Este (Jobbágy et al., 1995; Smith & Evans, 2007). El clima hacia el Este y el Sur de las estaciones de muestreo se vuelve más frío y árido (clima semiárido templado frío, Bsk, y clima árido templado frío, Bwk) (Peel et al., 2007).
La variación intensa del clima, el relieve y el suelo determinó la enorme heterogeneidad fisonómico - florística presente en una superficie relativamente pequeña. En los sitios más occidentales de altitud intermedia y húmedos se desarrolló el bosque natural, el puro de N. pumilio y mixto con A. araucana al Oeste sobre laderas de exposición Sur y Oeste, y el abierto de A. araucana al Este. La estepa fue la unidad de vegetación más extensa en los suelos zonales, pero en las áreas de mayor altitud los arbustos fueron muy escasos (Frugoni et al., 2016). Las praderas edáficas de herbáceas no graminosas crecieron en los fondos de valle sobre suelos intrazonales anegados (higrófila), y en los afloramientos rocosos basálticos y andesíticos de altura sobre suelos azonales superficiales (xerófila) (Szarzynski, 2000; Frugoni et al., 2016; Matteucci et al., 2019). El peladero es un área de acumulación y deflación intensamente modelada por factores naturales y antropogénicos, cuya superficie está cubierta por tefra volcánica (lapilli y ceniza gruesa y fina) no alterada post-deposicionalmente y dominada por pumita (Frugoni et al., 2016). A pesar de la enorme heterogeneidad, las especies fueron preferentemente herbáceas (81%) y nativas (94%).
En las unidades de vegetación se registró una abundante cantidad de especies de plantas comunes, a juzgar por el alto valor de similitud global basada en la composición. Sin embargo, el desempeño de estas especies fue desigual, teniendo en cuenta que la similitud basada en la frecuencia fue extremadamente baja, y sólo aumentó entre unidades de fisonomía equivalente (e.g., bosque puro y bosque mixto). En estas unidades también se registró una enorme cantidad de especies únicas y una escasa cantidad de cosmopolitas. Este patrón caracterizado por la presencia en el área de estudio de un amplio conjunto de especies comunes que exhibieron un desempeño equivalente sólo entre las unidades fisonómicamente similares, estaría explicado por la pronunciada variabilidad ambiental que promovería la existencia de una escasa cantidad de plantas capaces de adaptarse a ella. Esta distribución desigual de la abundancia de especies, definida por una enorme mayoría de especies raras y no dominantes, es además compatible con el patrón generalizado presente en las comunidades de plantas (McGill, 2011). La pradera higrófila fue la unidad que exhibió la mayor riqueza y proporción de especies únicas, que es compatible con la diversidad de los humedales de la Patagonia (Chimner et al., 2011; Weigandt et al., 2015). La dominancia de las asteráceas y gramíneas fue previamente indicada en la Patagonia (Oyarzábal et al., 2018; Matteucci et al., 2019).
En el área de estudio, la dominancia de hemicriptófitas en cantidad y frecuencia de especies es indicativa del efecto del frío sobre la vegetación; estas plantas producen vástagos en primavera, pero sólo sobrevive al invierno la parte basal que porta las yemas protegidas por las porciones secas. Las caméfitas fueron particularmente importantes en la pradera xerófila. Esta unidad sería más fría y seca que las circundantes, porque estaría expuesta a menor temperatura y mayor amplitud diaria y estacional en comparación debido a la mayor insolación y frecuencia e intensidad del viento y la menor humedad del aire en verano (Szarzynski, 2000). En consecuencia, la forma de vida caméfita representaría al mismo tiempo una adaptación al frío y la aridez. Estos afloramientos rocosos se interpretan como refugios de paleoflora adaptada al frío (Speziale & Ezcurra, 2012), e impondrían restricciones al desarrollo de otras formas de vida (e.g., a las geófitas por el escaso desarrollo de suelo).
Las terófitas ocuparon casi todas las unidades de vegetación pero exhibieron muy baja frecuencia, probablemente porque el clima ejercería un efecto negativo sobre especies que dependen exclusivamente de la reproducción sexual (Korner, 2003). Las geófitas estuvieron presentes en la mayoría de las unidades de vegetación pero con una frecuencia baja; estas plantas atraviesan la estación fría mediante órganos subterráneos protegidos bajo el suelo. Las nanofanerófitas, representadas por los arbustos, ocuparon todas las unidades pero con una frecuencia particularmente baja en la estepa de altura, lo que reflejaría la baja capacidad de adaptación de esta forma de vida al frío más extremo (Korner, 2003; Marini et al., 2012).
Las mesofanerófitas, que incluyeron árboles de mediana altura, alcanzaron la mayor riqueza y abundancia en las áreas occidentales de altitud intermedia, que es una tendencia global de la distribución arbórea asociada con el aumento de la humedad y la ausencia de frío extremo (Vázquez & Givnish, 1998; Bhattarai & Vetaas, 2003). Los cambios dimensionales y arquitecturales a lo largo de la altitud de N. pumilio y N. antarctica, que conforman el límite altitudinal superior del bosque y el matorral, constituyen mecanismos de adaptación al frío que se suman a la condición de especies deciduas (González et al., 2006; Stecconi et al., 2010; Peri et al., 2013; Matteucci et al., 2019). Pinus ponderosa ocupa el 95% de la superficie plantada en la Patagonia andina, debido principalmente a la capacidad de tolerar la sequía de verano, y las bajas temperaturas de invierno (Oliver & Ryker, 1990; Abella & Denton, 2009; MAGyP, 2014).
Las plantas exóticas espontáneas fueron hemicriptófitas y terófitas, y exhibieron una riqueza y frecuencia muy baja en todas las unidades de vegetación. Este patrón es compatible con la “hipótesis del filtro climático”, que propone que la distribución de especies nativas y exóticas está regulada fundamentalmente por el clima y no por la actividad humana (e.g., creación de nuevos nichos ecológicos, esfuerzo de introducción de especies) (Weiher & Keddy, 1999). Por el contrario, en muchas regiones la distribución de las plantas exóticas se relaciona con la presión de los propágulos y la disponibilidad de nuevos nichos creados por la actividad humana (Marini et al., 2012).
En los hábitats fríos permanentes y ocasionales, la supervivencia y el desarrollo de las plantas depende de la sincronización del ciclo de vida al periodo más cálido, una forma compacta de baja estatura y la protección basal y subterránea de las yemas (Kaplan et al. , 2003; Korner, 2003, Ott et al. , 2019). Esto permite desacoplar la temperatura del aire y la de planta, provocando una acumulación de calor en el dosel y la capa superior del suelo durante los períodos de radiación moderada a alta. Como consecuencia de estas adaptaciones, el espectro florístico tiende a estar dominado por hemicriptófitas y caméfitas; las geófitas son importantes en los climas más estacionales, y las terófitas sólo adquieren relevancia en los ambientes más cálidos (e.g., pisos altitudinales inferiores) (Korner, 2003).
El patrón de las formas de vida de Aguas Frías y Litrán fue consistente con el de otras regiones a lo largo de gradientes de altitud y latitud. En el NO de India entre 4180 a 6000 m, el espectro florístico estaba dominado por hemicriptófitas (62,1%) y terófitas (22,3%), y por encima de los 4900 m, las terófitas desaparecieron y las hemicriptófitas adquirieron una importancia mayor (Klimes, 2003). En los Montes Hindu Kush del NO de Pakistán, entre 4000 y 5400 el aumento de la altitud se relacionó con el aumento de hemicriptófitas y geófitas, y la disminución de fanerófitas, caméfitas y terófitas (Agakhanyanz & Breckle, 1995). En la cordillera de Qilian del N de China, entre 1400 y 2000 m, la riqueza de terófitas fue 48%, pero por encima de los 3600 m disminuyó hasta menos de 5%, mientras que las hemicriptófitas aumentaron con la altitud, y fueron la forma de vida dominante por encima de 2400 m y la única por encima de 4000 m (Wang et al. 2002). En el Monte Amstrong de Nueva Zelanda entre 1.200 y 2.200 m, la dominancia de hemicriptófitas y caméfitas aumentó con la elevación (84%), mientras que las geófitas y nanofanerófitas se restringieron a las altitudes más bajas (Mark et al. 2000). En la Sierra Nevada de California entre 3045 y 3350 m, la riqueza y cobertura de terófitas disminuyó en forma intensa con el aumento de la altitud (Jackson & Bliss, 1982). En las latitudes altas, a medida que disminuye la cantidad acumulada de días con temperatura media por debajo de un determinado umbral térmico, las meso- y microfanerófitas dan paso a las nanofanerófitas y caméfitas, y las hemicriptófitas se vuelven dominantes y las terófitas raras (Kaplan et al., 2003; Harrison et al., 2010).
En la transición bosque - estepa de la Patagonia argentina, las formas de vida dominantes fueron compatibles con la adaptación al frío. Sin embargo, existen adaptaciones a factores simples o múltiples de estrés abiótico y biótico con manifestaciones en el tamaño y la forma de los individuos. En particular, en esta área de estudio las plantas tendieron a presentar en forma simultánea tamaño pequeño, forma cespitosa y en cojín compacto, cutícula gruesa y reproducción vegetativa; estos atributos de estructura y desarrollo representarían ventajas frente a la aridez, el frío, el viento y la herbivoría (Cabrera, 1976; Golluscio et al., 1982, 2011; Soriano & Sala, 1983; Aguiar et al., 1996; Lauenroth, 1998; León et al., 1998; Ghermandi et al., 2004; Villagra et al., 2011; Oyarzábal et al., 2018; Matteucci et al., 2019).
Conclusiones
El intenso gradiente ambiental del área de estudio modeló la variabilidad composicional y fisonómica de la vegetación, pero el frío sería un regulador significativo de los patrones y procesos ecosistémicos a juzgar por la dominancia de hemicriptófitas, y la presencia de terófitas, geófitas, que representan plantas que durante el invierno mueren, o se les reduce completa o significativamente la parte aérea. Sin embargo, la sequía de verano también juega un papel clave y en consecuencia, el clima patagónico frío y ventoso, de transición húmedo árido, continental y mediterráneo, impone enormes restricciones al desempeño de las plantas. Los modelos climáticos regionales proyectan durante el presente siglo, un aumento de la temperatura y una disminución de la precipitación que afectaría en forma particular al invierno. En el ecotono bosque - estepa, el mayor estrés hídrico y térmico resultante promovería el aumento de las especies xerófilas, la disminución de las criófilas y la retracción de la vegetación arbórea, hidrófila y la que habita en los afloramientos rocosos. Las comunidades estarían expuestas a una mayor tensión ambiental, que además podría aumentar su sensibilidad a la degradación antropogénica actual. Es necesario realizar más investigación descriptiva y experimental sobre la relación clima - vegetación, teniendo en cuenta la presencia de factores de estrés naturales y artificiales que ponen en riesgo la funcionalidad de los ecosistemas patagónicos.
Contribución de los autores
Todos los autores colectaron la información de campo, AD fue el responsable principal del diseño, el análisis de los datos y la redacción del trabajo, AMe clasificó las plantas y AMo llevó a cabo los análisis basados en SIG

