











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink1. INTRODUCCIÓN
Las superficies con suelos salinos se incrementan significativamente a nivel mundial debido a la actividad antrópica, al ampliar las zonas de regadío. Aproximadamente la quinta parte de las 2.800 millones de hectáreas cultivables del planeta están afectadas por la salinidad, situación que tiende a agravarse (Ben Dkhil y Denden, 2010). Las prácticas de riego inadecuadas contribuyen a la salinización de los suelos. Así, el uso de aguas con alta conductividad eléctrica, sistemas de riego ineficientes o sistemas de distribución defectuosos pueden ocasionar la acumulación de sales en la superficie del suelo. El ascenso de las napas freáticas, debido al riego excesivo puede acelerar el proceso de salinización (Sun et al., 2009; Rodrigues de Souza et al., 2012).
Este avance desmesurado de la cantidad de nuevas tierras que son afectadas por las sales, restringe la producción vegetal (Díaz- López et al., 2012). Los efectos adversos de la salinidad sobre el crecimiento vegetal, están asociados a la disminución del potencial hídrico del suelo, la toxicidad de los iones, el desbalance nutricional, o una combinación de estos factores (Munns y Tester, 2008). Esta situación exige la selección de especies tolerantes a la salinidad, y el desarrollo de prácticas agronómicas tendientes a incrementar la resistencia de las plantas al estrés salino.
El vinal (Prosopis ruscifolia G.) es una especie altamente tolerante al estrés salino, que presenta múltiples aplicaciones: madera, leña, forraje, etc. Por estas características, es apto para la realización de plantaciones en zonas salinas, donde no pueden desarrollarse otras especies de importancia forestal (Velarde et al., 2003). Su elevada tolerancia a la salinidad se basa en la realización de ajuste osmótico, la baja traslocación de Na+ a la parte aérea, y un eficiente sistema de enzimas antioxidantes, que detoxifican a los radicales libres producidos como consecuencia del estrés. Sin embargo el crecimiento es inhibido significativamente por 0,4 mol L-1 de NaCl, por lo que resulta de interés desarrollar un método simple que permita mitigar el efecto del estrés salino (Meloni et al., 2008).
La adición de Ca+2 ha mitigado la inhibición del crecimiento producido por el estrés salino en Linum usitatissimum (Khan et al., 2010), Sorghum bicolor (Lacerda et al., 2004) y Triticum aestivum (Zaman et al., 2005), mientras que en otras no tuvo un efecto significativo (Kinraide, 1999; Reid y Smith, 2000). En Chenopodium quinoa y Acacia. occidentalis, la adición de Ca2SO4 incrementó el efecto inhibitorio del NaCl sobre la germinación (Abo-Kassem, 2007).
Los mecanismos involucrados en la reversión del efecto inhibitorio del NaCl sobre el crecimiento, mediante la adición de Ca2+ han sido poco estudiados. En trigo (Triticum aestivum L. cv Punjab 85), Zaman et al. (2005) observaron que este efecto estuvo asociado a un incremento en las concentraciones foliares de K+, y en el contenido relativo de agua. Por otra parte no existen referencias sobre el rol del Ca+2 en la absorción de nitrógeno, y su reducción, en plantas sometidas a estrés salino.
En este trabajo se puso a prueba la hipótesis de que la adición de Ca+2 mitiga la inhibición del crecimiento en plántulas de vinal sometidas a estrés salino con NaCl.
El objetivo fue determinar el efecto de la adición de Ca+2 sobre el crecimiento de plántulas de vinal bajo estrés salino con NaCl, y su mecanismo de acción.
2. MATERIALES Y MÉTODOS
Material vegetal y condiciones de crecimiento.
Los frutos se cosecharon en un individuo de P. ruscifolia en la localidad de Maco (27°51’20” S; 64°13’27” W), Provincia de Santiago del Estero, Argentina. Esta metodología permite disminuir el impacto de la elevada variabilidad genética propia del género Prosopis (Meloni, 2014). Las semillas se extrajeron manualmente y escarificaron con ácido sulfúrico durante 10 minutos, luego se enjuagaron con agua corriente durante 30 minutos. La siembra se realizó empleando toallas de papel humedecidas con agua destilada, que se incubaron en cámara de crecimiento por un lapso de 15 días a 25 ºC y 12 horas de fotoperíodo (González et al., 2006).
Se seleccionaron 450 plántulas de tamaño homogéneo, y cultivaron hidropónicamente en recipientes de 10 litros, conteniendo solución nutritiva de Hoagland al 25 % (30 plántulas por recipiente). Luego de 7 días de crecimiento en solución nutritiva, se realizaron los siguientes tratamientos: testigo (solución de Hoagland al 25 %), NaCl (solución de Hoagland al 25 % + 0,4 mol L-1 de NaCl), NaCl + Ca (solución de Hoagland al 25 % + 0,4 mol L-1 de NaCl + 5 mmol L-1 de CaSO4). En el tratamiento salino, el NaCl se adicionó gradualmente (50 mM cada 24 horas), hasta lograr la concentración final, para evitar un shock osmótico. Las pérdidas por transpiración se compensaron mediante el agregado de agua destilada, manteniendo constante el nivel de solución en los recipientes. El pH se ajustó a 6,5 mediante la adición de 100 mmol L-1 de KOH ó HCl. El ensayo se realizó en invernáculo, donde la radiación fotosintéticamente activa osciló entre un mínimo de 120 μmol m-2 s-1 durante la puesta de sol, hasta 1600 μmol m-2 s-1 al medio día, la temperatura media del aire fue de 28 ºC y la humedad relativa del 52 %.
Producción de biomasa, y composición mineral.
Luego de 30 días de tratamiento, las plantas se extrajeron de la solución nutritiva, lavaron con agua destilada y se llevaron a estufa de ventilación forzada, a 62 ºC hasta peso constante. En ese material se determinó la materia seca y composición mineral. La concentración de Na+ y K+ se cuantificó mediante espectrofotometría de llama (Jameel Al-Kahayri, 2002), Ca+2 y Mg+2 mediante espectrometría de absorción atómica (Ferreira-Silva et al., 2008) y Cl- por titulación con AgNO3 (Ferreira-Silva et al., 2008).
Metabolismo del nitrógeno.
La concentración de nitrato se determinó a través del método de Cataldo et al. (1975). La actividad nitrato reductasa fue cuantificada empleando el método de Klepper et al. (1971), con algunas modificaciones. Para ello, se tomaron aproximadamente 100 mg hojas y 300 mg de segmentos de raíces de 5 mm de longitud. El material fue infiltrado con 5 mL de una solución de incubación conteniendo buffer fosfato de potasio 0,1 M, pH 7,5; KNO3 0,05 M e isopropanol 1 % (v/v). La incubación se realizó durante una hora, a 30 ºC, en oscuridad. Para estimar la cantidad de nitrito formado, se adicionó 1 mL de sulfanilamida 1 % in HCl 1 N y naphtylethylene diamine dihydrochloride 0.02 %. Por último se leyó la absorbancia a 540 nm. La concentración de proteínas solubles totales fue determinada de acuerdo a Bradford (1976) con albúmina sérica bovina como standard, y la actividad enzimática fue expresada en µmol NO2- mg-1 proteína h-1. La actividad glutamina sintasa se determinó según el método descripto por Berteli et al. (1995), con las modificaciones citadas por Silveira et al. (2003).
Diseño experimental y análisis estadístico.
Se utilizó un diseño completamente aleatorizado con 5 repeticiones; la unidad experimental estuvo representada por un recipiente de 10 L conteniendo 30 plántulas. De cada recipiente se tomó una muestra compuesta de 10 plántulas para determinación peso seco y cuantificación de iones, 10 plántulas para actividad nitrato reductasa, y 10 plántulas para actividad nitrito reductasa. Los resultados se analizaron con ANOVA y test de Duncan.
3. RESULTADOS Y DISCUSIÓN
La adición de 0,4 mol L-1 de NaCl en la solución nutritiva produjo una reducción significativa del 43 y 22 % en el peso seco de hojas y raíces, respectivamente (Figura 1). La suplementación con calcio, revirtió el efecto del estrés salino, alcanzándose valores de biomasa similares al testigo.
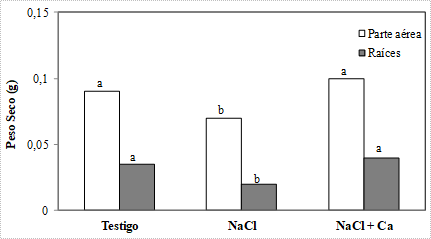
Figura 1. Peso seco de hojas y raíces de plántulas de vinal, crecidas en solución nutritiva de Hoagland 25 % (Testigo), con adición de 0,4 mol L-1 de NaCl (NaCl), y con adición de 0,4 mol L-1 de NaCl y 5 mmol L-1 de CaSO4 (NaCl + Ca). Para cada parte (hojas y raíces), barras seguidas por letras diferentes, difieren estadísticamente por el Test de Duncan al 5 %.
Estos resultados coinciden con los obtenidos en Licopersicum esculentum (Levent Tuna et al., 2007) y Atriplex halimus (Nedjimi and Daoud, 2009a), especies en las que la adición de Ca+2 revirtió la inhibición en el crecimiento producido por el estrés salino. Por otra parte difieren con lo reportado por Murillo-Amador et al. (2006), en plantas de Vigna unguiculata, quienes observaron que la adición de Ca+2 no logró revertir la inhibición del crecimiento, ni las alteraciones en la composición mineral. Sin embargo, tales diferencias pueden deberse a las condiciones experimentales, ya que dichos autores utilizaron Ca2NO3 aplicado como spray sobre las hojas.
La suplementación con Ca+2 también generó un aumento en la relación biomasa de raíces/biomasa de hojas, que presenta una gran importancia ecofisiológica, bajo ciertas condiciones ambientales, ya que dota a la planta una mayor superficie para la absorción de agua y nutrientes. Esta respuesta, puede justificar en parte, el rol benéfico de la adición del calcio, sobre el crecimiento de las plántulas sometidas a estrés con NaCl.
El estrés salino afectó la concentración de iones, tanto en hojas como en raíces (Tabla 1).
Tabla 1. Composición mineral en hojas y raíces de plántulas de vinal, crecidas en solución nutritiva de Hoagland 25 % (Testigo), con adición de 0,4 mol L-1 de NaCl (NaCl), y con adición de 0,4 mol L-1 de NaCl y 5 mmol L-1 de CaSO4 (NaCl + Ca).
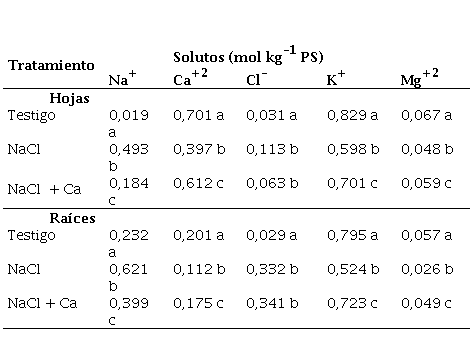
Para cada parte (hojas ó raíces), letras diferentes en cada columna indican diferencias significativas al 5 % por el test de Duncan
La suplementación con 5 mmol L-1 de Ca+2 disminuyó la concentración de Na+ en raíces y hojas de plantas crecidas en 0,4 mol L-1 de NaCl, y no modificó los niveles de Cl- . El efecto fue más notorio en hojas, donde la adición de calcio disminuyó en 63 % la concentración de Na+, en comparación con el tratamiento salino no suplementado. En raíces la disminución en la concentración de Na+ fue solamente del 36 %.
Estos resultados demuestran que el Ca+2 incrementa la exclusión del Na+ de la parte aérea, protegiendo a las hojas del efecto tóxico de este catión. Esta observación coincide con los resultados reportados por Tattini y Traversi (2009) en olivo, quienes concluyeron que el Ca+2 disminuye las concentraciones foliares de Na+, incrementando la tasa fotosintética, y el desempeño fotoquímico de la especie.
Una respuesta característica de las plantas sometidas a estrés salino es la disminución de la concentración de K+, y la sustitución de este catión por el Na+, que puede conducir al desbalance de iones. Dichos nutrientes pueden competir por su absorción en las células de las raíces, con un efecto adverso sobre el crecimiento y desarrollo. De este modo el mantenimiento de altas relaciones K+/Na+ es importante para la tolerancia a altas concentraciones salinas (Wu y Wang, 2012).
La salinidad disminuyó significativamente las relaciones K+/Na+ tanto en raíces como en hojas de vinal; sin embargo la adición de Ca+2 triplicó la relación K+/Na+ en hojas, con respecto al tratamiento de 0,4 mol L-1 de NaCl. En las raíces el efecto fue menos marcado, pasando de 0,8 en el tratamiento salino a 1,8 en el tratamiento con NaCl y suplementación con Ca+2. En coincidencia con este resultado (Nedjimi and Daoud, 2009a) demostraron que en Atriplex halimus la adición de Ca+2 al ambiente radicular de plantas sometidas a estrés salino, mantiene o mejora la absorción selectiva de potasio a altas concentraciones de sodio, lo que previene el efecto deletéreo de un exceso de Na+.
Bajo condiciones de elevada salinidad, disminuye la relación Ca+/Na+ en el suelo, lo que puede alterar las propiedades de la pared celular y la membrana plasmática, ya que el Na+ desplaza al Ca+2 asociado a las mismas (Guimarães, 2011). Como consecuencia de ello se incrementan de las concentraciones de Na+ dentro de las células, que puede inhibir la actividad de algunas enzimas, produciendo alteraciones en el metabolismo, en la absorción de K+, y su partición en la planta, afectando la apertura estomática (Siddiqui et al., 2012). En conjunto, estos cambios inhiben el crecimiento vegetal.
El NaCl produjo una importante disminución en las concentraciones de Ca+2 en hojas y raíces de vinal, y esa respuesta fue parcialmente revertida por la adición de 5 mmol L-1 de CaSO4. Un efecto similar se observó con las concentraciones de Mg+2.
El incremento en las concentraciones de Ca+2 en hojas y raíces de plántulas estresadas presenta una gran importancia metabólica, ya que está involucrado en la transducción de señales (Kader y Lindberg, 2010). La unión del Ca+2 a sensores de Ca+2, tales como la calmodulina, induce respuestas fisiológicas apropiadas, vía la modulación de las funciones de proteínas target, (por ejemplo las quinasas), involucradas en el transporte de iones, metabolismo, modificaciones postranscripcionales de proteínas, y expresión de genes (Kim et al., 2009). Además el Ca+2 puede activar la ruta SOS3-SOS2 quinasa proteica, que conduce al incremento de la expresión del transportador de la salida de Na+ (SOS1) del plasmalema, y probablemente restringe el ingreso de Na+ a través del transportador con alta afinidad para el K+ (HKT1), lo que permite mantener la homeostasis intracelular K+/Na+ (Türkan y Demiral, 2009).
Tanto el estrés salino, como la adición de calcio a la solución nutritiva, afectaron el metabolismo del nitrógeno en las plántulas de vinal. El NaCl redujo en 56 y 67 % la concentración de NO3-, en hojas y raíces respectivamente; en tanto que en plantas sometidas a estrés salino y suplementadas con Ca+2, la reducción fue sólo de 36 y 19 %, respectivamente (Figura 2 A).
La actividad nitrato reductasa in vivo (NR) fue considerablemente mayor en hojas que en raíces (Figura 2 B). El estrés salino redujo la actividad NR tanto en hojas como en raíces, en 40 y 50 % respectivamente; en el caso de plántulas suplementadas con Ca+2, la inhibición fue menos marcada, alcanzando valores 20 y 25 % inferiores al testigo no tratado, en hojas y raíces. Un efecto similar se observó en la actividad glutamina sintasa (Figura 2 C).
Los resultados obtenidos en este trabajo indican que las plántulas de vinal expuestas a estrés salino presentaron una disminución en las concentraciones de NO3-, efecto que fue parcialmente revertido mediante la adición de calcio en la solución nutritiva (Figura 2 A). En este sentido, la salinidad puede inhibir la absorción de N, y su utilización, interfiriendo con la reducción del NO3-, y la síntesis proteica (Siddiqui et al., 2008). Su interferencia con la absorción de nitrógeno puede darse en dos niveles: competencia entre iones Cl- y NO3-, y alteraciones en la integridad de las membranas celulares (Nedjimi and Daoud, 2009b). Estos resultados coinciden con los reportados por Silveira el al. (2001) en Vigna unguiculata, donde la adición de 100 mM de NaCl produjo una reducción de 35 a 70 % en las concentraciones de NO3- de plantas aclimatadas y no aclimatadas, respectivamente.
La actividad nitrato reductasa in vivo (NR) fue considerablemente mayor en hojas que en raíces (Figura 2 B), lo que también fue observado en plántulas de Prosopis alba (Meloni et al., 2003). El estrés salino redujo la actividad NR en hojas y raíces y la suplementación con calcio aumentó su actividad, aunque no se alcanzaron los valores observados en el testigo.
La actividad NR, es altamente dependiente del flujo de NO3- y es inhibida por el estrés salino (Ebrahimian and Bybordi, 2011). Carrillo et al. (2005) estudiaron la actividad y expresión de la NR en plantas de trigo sometidas a estrés salino con NaCl, bajo distintas concentraciones de NO3-. Estos autores observaron que a bajas concentraciones de NO3-, el NaCl incrementó la actividad NR, lo que sugiere que el Cl- puede imitar al NO3- como molécula señal, para inducir la transcripción en hojas y raíces. Sin embargo, determinaron que los niveles de proteína NR decrecieron como consecuencia del estrés salino, indicando un efecto inhibitorio de la sal sobre la traslación del NR mRNA, o un aumento en la degradación proteica.
Resultados similares se obtuvieron con la actividad de la enzima glutamina sintasa (GS), en la cual el Ca+2 mitigó el efecto inhibitorio del NaCl. En algunas especies, como Anacardium occidentale, el NaCl incrementó la actividad GS, (Silveira et al., 2003), la cual estuvo asociada a un aumento en la síntesis de prolina, un soluto osmocompatible que permite la realización de ajuste osmótico, con la concomitante absorción de agua en suelos salinos. Coincidiendo con este resultado Meloni et al (2011), observaron un incremento significativo en la concentración de dicho soluto en plántulas de vinal sometidas a estrés salino, y suplementadas con calcio.
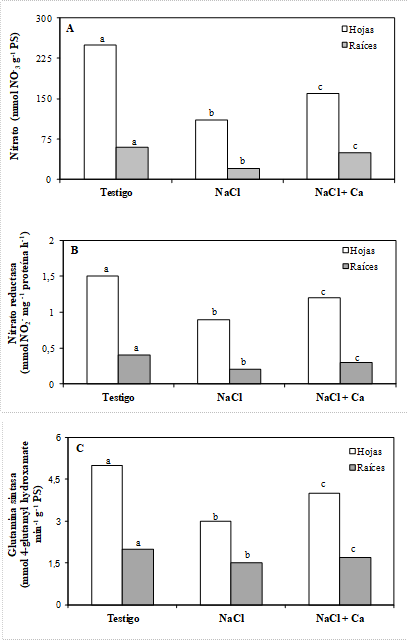
Figura 2. Concentración de nitrato (A), y actividades nitrato reductasa (B), y glutamina sintasa (C) en hojas y raíces de plántulas de vinal, crecidas en solución nutritiva de Hoagland 25 % (Testigo), con adición de 0,4 mol L-1 de NaCl (NaCl), y con adición de 0,4 mol L-1 de NaCl y 5 mmol L-1 de CaSO4 (NaCl + Ca). Para cada parte (hojas y raíces), barras seguidas por letras diferentes, difieren estadísticamente por el Test de Duncan al 5 %.
4. CONCLUSIONES
Los resultados obtenidos en este trabajo indican que la suplementación con Ca+2 mitiga la inhibición del crecimiento producida por el estrés salino en plántulas de Prosopis ruscifolia. Esta respuesta en parte se debe al incremento en las concentraciones de K+; Ca+2 y Mg+2, y una disminución en la concentración de Na+, tanto en hojas como en raíces. También puede atribuirse a la reversión del efecto inhibitorio del NaCl sobre sobre el metabolismo del N, a través de un incremento en las concentraciones de NO3-, y en la actividad de las enzimas nitrato reductasa y glutamina sintasa.

