











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkINTRODUCTION
Sedimentary dynamics have been shown to be biologically modified in environments where sediments are colonized by benthic microorganisms (Noffke and Paterson, 2008). However, one of the greatest challenges in the study of the geological record is to recognize biogenic influence in the sedimentary structures of ancient sediments. The study of modern microbial mats is especially useful for this purpose because it allows a precise analysis of present-day biogenic structures and the environmental conditions in which they are formed and can, then, be used as analogues for the fossil record (Cuadrado and Blasi, 2018; Cuadrado, 2020). On some occasions the general hydrodynamic regime in the modern sedimentary environment colonized by microbial mats may show differences between winter and summer due to water level fluctuations (Cuadrado, 2020). Such seasonal fluctuations can be recorded in the sediment through the precipitation of certain minerals. According to this, one of the most interesting aspects of microbial influence on sediments is their impact on carbonate precipitation, which has been proven to facilitate microbial mat preservation and lithification, especially in carbonate environments (Reid et al., 2000, Dupraz et al., 2004), enabling the identification of microbial mats in the fossil record. However, for precipitation to occur, it is essential to develop a delicate balance between physicochemical processes and microbial activities (Dupraz and Visher, 2005). Therefore, the study of mineral precipitation in these systems becomes a complex endeavour as it involves the interaction between biotic and abiotic factors (Maisano et al., 2020).
Mineral precipitation in microbial ecosystems has been studied in detail in modern and past carbonate environments, where rapid mineralization favours preservation of microbialites (Reid et al., 2000, Dupraz et al., 2004). In contrast, in modern siliciclastic environments, microbial mats are generally unlithified and form planar soft organic layers that colonize depositional areas that are subject to erosive episodes by high-energy currents and to sediment deposition (Noffke, 2003). Nevertheless, the present investigation documents evidence of calcium carbonate precipitation in a siliciclastic coastal environment that enables the lithification of ripple marks and microbial mats. Sedimentary structures such as ripple marks, formed by severe hydrodynamic events, are biostabilized by microbial activity and the presence of exopolymeric substances (EPS; Cuadrado, 2020), which may produce sinoidal structures. In addition, the presence of a carbonate lamination over these sand sedimentary structures possibly would enhance the lithification of the colonized ripple structure. The analysis of this carbonate precipitation is the focus of this paper, which coupled with the sinoidal structure, may provide potential for preservation in the fossil record. Paso Seco coastal area is an excellent case study to present the formation of carbonate lamination in microbial mats, which is a consequence of a combination of biogeochemical and physicochemical factors (Perillo et al., 2019, Maisano et al., 2020).
STUDY AREA
The Paso Seco coastal environment (40°38’40’’S; 62°12’22’’W; Fig. 1) is in the northern coast of Patagonia (Argentina), which is characterized by a mesotidal regime with mean and maximum tidal ranges of 1.62 m and 2.5 m, respectively. The climate of the region is semiarid, as stated by the recent climatic classification of the Argentinian Pampas proposed by Aliaga et al. (2017), characterized by a potential evapotranspiration greater than precipitation, which generates enough aridity to limit vegetation development. The elongated study area (3.5 km in length and 0.4 km in width) corresponds to an ancient tidal channel closed more than 100 years ago by the generation of a sand spit (1.8 km wide) due to longshore sediment transport. Thus, the study area is a coastal environment behind the sand spit, a semi-closed basin categorized as a supratidal zone because it is flooded by seawater under specific oceanographic conditions when storm surges provoke seawater overpass the sand spit (Stempels Bautista, 2019). The flooding might occur up to 43 times per year, but the area remains unaffected by daily flood tides (Perillo et al., 2019; Maisano et al., 2019). The high-energy flooding is followed by a slow receding current and the retention of a few cm-deep seawater layer over the sedimentary surface for several days to weeks. Furthermore, the extreme-energy water currents produce microbially-induced sedimentary structures of significant dimensions (e.g., large mat tears, between 2 and 8 m; large roll-ups ≈ 25 cm in height; Fig. 1c) (Maisano et al., 2019).
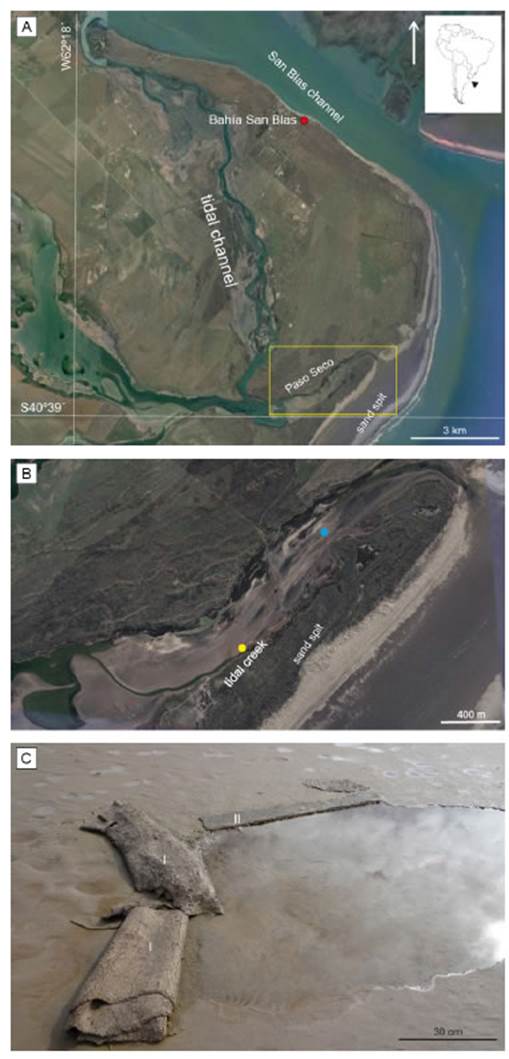
Figure 1 a) Location of the Paso Seco flat behind a sand spit (yellow square). b) Close-up view of the study area (yellow square in A). The blue mark indicates the location of the water level station. The yellow mark indicates the location of the tidal creek where a water sample was taken. c) Large microbially-induced sedimentary structures created by deformation processes over the tidal flat during extreme storm that occurred in June 2018, as seen in August 2018, 20 days after a moderate storm. I) microbial roll-ups, II) flipped-over edges.
The area is characterized by ≈ 1 cm-thick epibenthic mats that colonize the sediments (Cuadrado et al., 2015). The main constructor member of this microbial consortium is the filamentous cyanobacteria in the order Oscillatoria sp. where the Coleofasciculus (Microcoleus) chthonoplastes is the most abundant cosmopolitan cyanobacteria species in the area (Cuadrado and Pan, 2018). Other important microorganisms of the microbial association are pennate diatoms that are the first colonizers (Pan et al., 2017). Once the microbial mat is formed, the consortium of microorganisms, EPS and sediment produces a poorly permeable sediment surface, which causes slow water infiltration from the surface and allows gradual evaporation of seawater and evaporite precipitation (Perillo et al., 2019). Thus, Paso Seco can be considered a saline basin conditioned by the hydrological conditions and biological processes (Perillo et al., 2019).
METHODS
To perform hydrochemical analyses, six water samples were taken from the study area in September 2017: five samples from different ponds with different salinities and one sample from a tidal creek (Fig. 1b). Water Ca2+, CO32- and SO42- concentrations were determined by titration (Ca2+, CO32-) and turbidimetric (SO42-) methods following standard methods (APHA, 2005), whereas water salinity was determined gravimetrically as described by Strickland and Parsons (1972).
To know the frequency of seawater entering the study zone, water-level fluctuations were measured in the tidal flat using a HOBO water level logger (Onset-model U20; 2.5 cm diameter, 15 cm length). It was deployed into a vertically buried, perforated PVC pipe, 40 cm in depth from the sedimentary surface. The sensor recorded water level and temperature every 10 min. The water level data were corrected by atmospheric pressure by means of another logger placed in an upper level close to the tidal flat. The present study shows the water level and temperature recorded over a 10-month period in 2018.
Ripple field formation over the tidal flat was documented after the occurrence of a severe storm and was monitored throughout three field trips over a period of nearly six months since its creation, until December 2018. Firstly, ripples were examined two days after their creation, then in a following field trip carried out 20 days after a moderate storm that caused deposition of a thin sand layer over the previous ripples, and finally, six months afterwards. During the third field trip (December 2018) a sedimentary block (12 x 12 x 5 cm) which included the monitored ripples was taken for petrographic analysis. In addition, sedimentary cores were obtained using PVC tubes (3 cm diameter and 7 cm length) to examine the sedimentary texture and microstructures. They were opened in the laboratory, and subsequently analysed under a SMZ Nikon 1500 stereoscopic zoom binocular microscope. In addition, thin sections (thicker than the 30-μm traditional thickness of petrographic thin sections to avoid losing fragments of sample) obtained from the sedimentary block were prepared for petrographic analysis. Thin sections were analysed by a Nikon Eclipse POL 50i transmitted-light microscope, coupled with a camera. The analysis was carried out under plane-polarized light and cross-polarized light to identify mineral species. Three sections were stained with a solution of Potassium Ferrocyanide Ke (CN) 6, Alizarin and Hydrochloric Acid for determining the nature of the carbonate (Dickson, 1966).
RESULTS
Hydrodynamic processes and water evaporation
The topography of the study area presents a 0.03% general gradient towards inland. The maximum elevation (1.8 m above mean sea level) is in the northeasternmost area of the basin (Fig. 2a). On the other hand, the minimum elevation (0.5 m above mean sea level) corresponds to a tidal creek located in the inland area of Paso Seco (Fig. 2a), which is connected to a major tidal channel at the west of the study area (see Fig. 1).
The water level records show the storm-related inundations of the tidal flat (Fig. 2b). From March to June 2018, the sediment flat remained exposed for most of the time. On 25 June 2018, an extreme storm produced a seawater inundation of 70 cm over the microbial flat (green arrow, Fig. 2b) and the formation of a ripple field. The first field trip was done two days after the inundation (red arrow 1, Fig. 2b). Subsequently, seawater remained near the sedimentary surface until a moderate inundation on 22 July occurred (pink arrow, Fig. 2b). In this occasion, the water depth was 25 cm. Seawater remained near the microbial surface for 19 days including two subsequent inundations (black dashed line, Fig. 2b). A second field trip was conducted 20 days after the moderate inundation (red arrow 2, Fig. 2b). Then, the water level descended progressively below the tidal flat, causing subaerial exposure of the flat for 29 days (green dashed line, Fig. 2b). Later, during spring, the flat experienced eight floodings less than 20 cm water column, which alternated with flat exposures (orange dashed line, Fig. 2b), until the last field trip was conducted in December (red arrow 3, Fig. 2b).
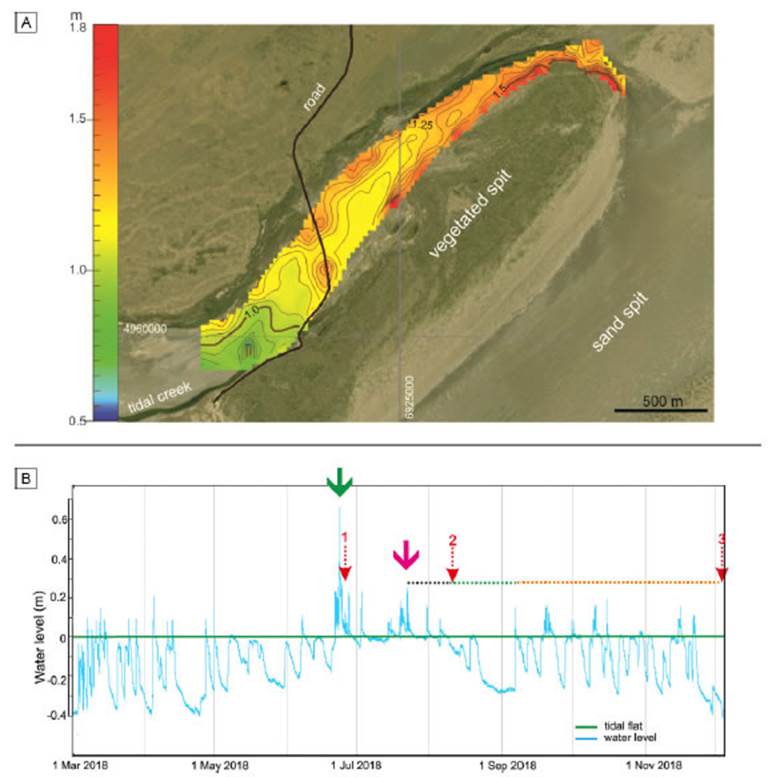
Figure 2 a) Topographic map of the study area. b) Water level record from March to December 2018. Green arrow indicates a high-energy flooding, during which the ripple field was created. Pink arrow indicates a subsequent moderate inundation. Black dashed line indicates a period during which the flat was episodically inundated. Green dashed line indicates a period during which the flat was subaerially exposed. Orange dashed line indicates a period of several floodings lower than 25 cm interspersed with subaerial exposure periods. Red dashed arrows indicate the date of the field trips. During field trip 3 a sedimentary block was sampled.
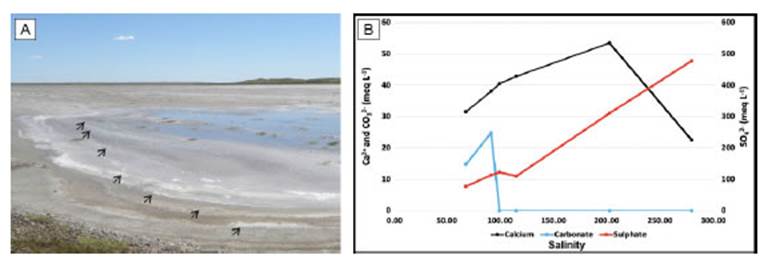
The topography of Paso Seco is characterized by several shallow ponds (≈ 4 cm in depth) containing seawater after inundations, whose volume progressively diminishes as seawater evaporates. Hydrochemical analysis were carried out in September 2017 to determine the evolution of water composition as it evaporates after an inundation. Salinity increased in the different tidal ponds in agreement to the visual signs of volume water reduction and evaporite presence (e.g., visual gypsum and halite precipitation; Fig. 3a). The chemical analysis of the water composition in these ponds indicated that CO32- concentration increased until salinity reached the value of 100, when an abrupt decrease in CO32- concentration occurred (Fig. 3b). In contrast, Ca2+ concentration showed an increase as salinity reached a value close to 202, when the maximum Ca2+ concentration was attained, and subsequently dropped rapidly. Moreover, the SO42- concentration raised gradually as salinity increased (Fig. 3b).
Ripple field characteristics
The extreme storm that occurred in June 2018 (green arrow in Fig. 2b) produced the modification of the microbial flat combined with the formation of sand ripple patches due to the sediment transport over the microbial mat. Ripples, made up of fine sand sediment, were characterized by asymmetrical 2D and 3D morphologies with straight and sometimes sinuous crestlines (Fig. 4a). Wavelengths ranged from 5 to 10 cm and heights reached 1 cm, resting on a sharp and planar microbial mat (green dashed line, Fig. 4b). The visualization of ripples immediately after the storm (1 shown in Fig. 2b) showed a sand layer on the sediment surface with mud present only in the trough (Fig. 4a).

Figure 4 a) Asymmetrical 2D ripples observed in Paso Seco flat two days after the storm of June 25, 2018. Red arrows indicate fine sediment deposited in the troughs of the ripple. b) A vertical section of a ripple structure obtained two days after the storm of June 25 2018. The microbial mat is delineated by a green dashed line. c) Tidal flat appearance on August 2018, 20 days after the moderate storm in July 2018. Sediment surface shows a smooth appearance caused by the colonization of the rippled surface by a biofilm. d) A vertical section of a ripple colonized by a biofilm (orange arrow). e) Close view of the red square in (d). Red dashed line: clay and silt sediment layer in the ripple trough. Orange arrow: biofilm on the sediment surface.
In July 2018 a moderate storm flooded Paso Seco (pink arrow in Fig. 2b). 20 days after this storm the study area was visited during a field trip (2 shown in Fig. 2b), allowing to observe that a thin biofilm colonized the whole ripple field (Fig. 4c). Moreover, cross-sections of the sediment permitted the recognition of the surficial biofilm (orange arrow in Fig. 4d, e), and below the biofilm the mud layer covering the ripple trough is identified (red dashed line in Fig. 4e).
Petrographic analysis
A sedimentary block containing the sand ripples deposited over a 1 mm-thick microbial mat (Fig. 5a) was petrographically analysed. The microbial mat (c frame in Fig. 5b) is composed of brownish organic matter laminae of ≈ 20 μm thickness (yellow arrows in Fig. 5c), interbedded with clay to fine-grained silt and micritic calcite laminae between 40 and 80 μm in thickness (orange arrows in Fig. 5c). The micritic layers sometimes are composed of peloids less than 100 μm in size (blue arrows in Fig. 5c- d), which may coalesce forming laterally continuous layers (pink arrows in Fig. 5c-d).

Figure 5 a) Vertical sedimentary profile of the block containing the ripple formed in June 2018 (yellow arrow). The location of the analysed thin section is indicated by the orange square. b) Thin-section cross-polarised light photomicrograph of the orange square in (a). Yellow arrow indicates the body of the ripple formed in June 2018; red dashed line indicates a layer made up fine silt and clay; green arrow indicates another ripple formed because of a moderate storm; orange arrow indicates a micritic layer covered by an organic matter lamina forming a biofilm. c) Cross-polarised light photomicrograph of a close-up of the microbial mat below the ripple (square c in b). Yellow arrows indicate brownish organic matter laminae; orange arrows indicate micritic calcite laminae; blue arrows indicate peloids; pink arrows indicate coalescent peloids. d) Cross-polarised light photomicrograph of a close-up of the microbial mat of (c). Blue arrows indicate peloids; pink arrows indicate coalescent peloids. e) Plane-polarised light photomicrograph of a close-up of the uppermost sand ripple (square e-f in b). Green arrow indicates the body of the ripple; b indicates an organic matter lamina forming a biofilm at the surface; orange dashed lines (m) indicate a laterally continuous dense micritic layer. Red dashed line indicates a lamina composed of fine silt and clay. f) Cross-polarised light photomicrograph of (e). g) Thin-section cross-polarised light photomicrograph of the square g in (b). Red dashed line indicates a lamina made up fine silt and clay. h) Plane-polarised light photomicrograph of a close-up of yellow square in (e). b indicates the biofilm covering the micritic layer (m). i) Cross-polarised light photomicrograph of (h).
The sand ripple deposited over the microbial mat is 6 mm in height and its trough is covered by a layer made up of fine silt and clay (limited by red dashed line in Fig. 5b). This fine-grained layer presents variable thickness, being thicker (≈ 1 mm) in the trough, and thinner (few tens of micrometres) towards the crest of the ripple. This layer presents a brownish interference colour under cross-polarized light (red dashed line in Fig. 5b, e-g) which is covered by a thin biofilm along the upper red dashed line (Fig. 5b, e-g). Upwards, another tiny train of sand ripples (up to ≈ 1 mm in height) covers the previous fine sediment layer and the thin biofilm (green arrows in Fig. 5b- e, f). This second train of ripples had not been recognized during the field trips due to its small size, and its identification was only possible under the microscope. Covering the previous laminae, a laterally continuous dense micritic calcite layer, 100-to-200 μm in thickness, is developed (m in Fig. 5e- f). The high-order interference colours of calcite under cross-polarized light allow to easily discern this lamina (Fig. 5h- i). The micritic layer is covered by an organic matter lamina forming a biofilm on the sediment surface (b in Fig. 5e- i).
DISCUSSION
The episodic storms control the water exchange between the study zone and the sea, while the geomorphology affects the sediment dynamics. Seawater enters the narrow mouth of the area forming fast currents that are essential for sand transport (Fig. 1b, 2a). During these severe storms, water depths over 70 cm are quickly reached over the microbial mats (Maisano et al., 2019). These quick currents produce large microbially-induced sedimentary structures created by deformation processes over the tidal flat, such as microbial roll-ups, flipped over structures, microbial folds, and erosional pockets (Fig. 1c). Moreover, the sediment transport during flooding forms ripple patches over the layered planar epibenthic microbial mat, and the presence of the microorganisms influences the stabilization of the ripples (Cuadrado, 2020). In that sense, once the ripples are created, when calm conditions are recovered, the filamentous microorganisms present in the underlying mat, such as O. Oscilatoriales (Cuadrado and Pan, 2018), can move quickly upward to the sand ripple due to their gliding motility (Shepard and Sumner, 2010). Consequently, the grains are entangled by microbial filaments and embedded by EPS, which causes the increase of the critical erosion threshold for the movement of particles (Paterson, 1994; Noffke et al., 2001).
The analysis of the sedimentary block obtained from the flat after the formation and colonization of the ripples (Fig. 5a), coupled with the analysis of the water level record (Fig. 2b), allows the recognition of the successive hydrodynamic events that took place in Paso Seco along 2018 and the identification of all the processes involved in ripple preservation. The strong storm of June 2018 (green arrow, Fig. 2b) created the ripple resting on the microbial mat (yellow arrow, Fig. 5a-b). After that strong event and during the subsequent relative quiet conditions, when there was no sediment reworking, a clay and silt sediment layer of varying thickness was deposited (red dashed line in Fig. 5b, e). Moreover, during this period, a reorganization of the microbial mat fabrics occurred, and binding and trapping processes took place. These processes would lead to the formation of a thin biofilm covering the sediment surface forming a sinoidal structure (see Noffke, 2010) (located along the upper red dashed line in Fig. 5b, e-g). The subsequent moderate storm in July (pink arrow in Fig. 2b) produced another ripple over that biofilm (green arrow in Fig. 5b), and finally the ripple was capped again by a thin biofilm formed during following subaerial exposure and several inundation events (green and orange dashed line in Fig. 2b). Although the genesis of the ripples was triggered by mechanical processes (thus abiogenic), that is, transport of non-cohesive detrital sediments (Kennedy, 1969), the presence of the filamentous microorganisms in the microbial mats over which the sand was deposited, produced a fast stabilization (Cuadrado, 2020).
The biofilm formed over the first ripple structure is not associated with a micritic lamina, probably because the evaporation rate between the storms in June and July 2018 was low. This is suggested by the fact that the sediment surface was not subaerially exposed during that period, as the water level remained approximately at the sediment surface (see period between green and pink arrow in Fig. 2b). On the other hand, the biofilm formed over the uppermost ripples covers a thin carbonate lamina (orange arrow in Fig. 5b). This carbonate was probably precipitated when the seawater retained in the saline basin reached CaCO3 supersaturation before the surface exposure (green dashed line in Fig. 2b). Attainment of CaCO3 supersaturation in Paso Seco is possible by a combination of factors, including a) the semiarid climatic conditions that characterize the area (Aliaga et al., 2017); b) seawater retention over the sediment surface, influenced by the topographic gradient towards inland; c) the biologically-induced changes in sediment permeability by microbial mats (Perillo et al., 2019), which helps seawater retention over the sediment; and d) the aid of fast blowing dry winds. The combination of all these factors induces the seawater evaporation that leads to precipitation firstly of calcite, and subsequently of gypsum when CO32- ion is completely depleted due to the precipitation of calcite but the water still contains calcium and sulphate anions (Fig. 3b).
Moreover, Maisano et al. (2020) suggested that biotic processes also play an important role in carbonate precipitation in Paso Seco. On one hand, these authors described the presence of occasional whitings in the study area and suggested that microorganisms in suspension in the water column could act as nucleation sites and could even promote carbonate precipitation in relation to photosynthesis processes. A similar explanation, increased photosynthetic activity and cell metabolism of cyanobacterial picoplankton, was sustained by Thompson et al. (1997) for whitings in a lacustrine water system. On the other hand, Maisano et al. (2020) also suggested that the peloidal textures shown by some carbonate laminae in Paso Seco may be related with precipitation of calcite in the EPS of the microbial mats, similarly to what has been described in microbial mats from a salina environment (Gerdes et al., 1994). Taking into account all these considerations, the formation of the carbonate lamina that covers the studied ripples resulted likely from the combination of CaCO3 supersaturation attainment due to intense seawater evaporation retained over the sediment surface plus the existence of nucleation sites (suspended microorganisms and/ or EPS in the mats) for carbonate precipitation.
The sedimentary processes identified in Paso Seco, in close relationship to the different hydrodynamic events that took place along 2018, allow us to interpret the main factors that contributed to the preservation of the studied ripples. First, the biostabilization of the ripples caused by the development of a thin biofilm over their surface could help to prevent these structures from erosion. Similarly, Cuadrado et al. (2021) demonstrated the important role of microbial mat development in the preservation of a vertebrate footprint formed in Paso Seco, which agrees with the postulation of Seilacher (2008), who argued that cyanobacterial films are instrumental in the preservation of animal tracks and drag marks. Likewise, numerous authors have highlighted the role of microbial mats in enhancing exceptional fossil preservation (Varejão et al., 2019). Secondly, the present study shows that carbonate precipitation within the microbial mats that colonize the ripples is crucial for the early lithification of these structures, which might significantly increase their preservation potential in the geological record.
Although this study on present-day microbial mats proves the importance that microbial colonization may have in the preservation of ripples in some environments the application of these results to the ancient successions is challenging due to the difficulties on recognizing microbial signatures in the fossil record. The observations on present-day ripples of Paso Seco show that a thin fine-grained sediment lamina over the ripple (upper red dashed line, Fig. 5b) is the result of the colonization of the sediment surface by a biofilm. However, this feature might be difficult to identify in ancient deposits, either because it is not preserved or because it is misinterpreted as a mud drape produced by purely physical processes (Cuadrado, 2020). Nevertheless, the recognition of a continuous fine-grained lamina and a continuous micritic carbonate lamina covering a ripple train is a good first step to consider that ripple preservation and early lithification may have been enhanced by the development of a biofilm or microbial mat.
CONCLUSIONS
This study documents ripple formation, stabilization, and the beginning of their lithification in a colonized coastal environment. Ripples are developed due to episodic high-energy events, and after that, subsequent calm conditions allow the accumulation of a clay and silt layer and the creation of a sinoidal structure by the development of a thin biofilm over the sedimentary sand structures. Furthermore, microbial mats change the permeability of the sediment, so the seawater that frequently enters the study area and provides Ca2+ and CO32- ions can progressively evaporate forming a continuous lamina of micritic calcite, which promotes the early lithification of the sediment.
Overlying the micrite layer a biofilm is formed, which can enhance the biostabilization of the ripples by migration of microbial filaments and production of EPS. Therefore, the structures would acquire a first preservation that may protect them from following erosional processes. These observations on a present-day sedimentary environment may be particularly useful to better understand the formation, biostabilization and early lithification of ripple marks and may have a potential application for the interpretation of the stratigraphic record.

