











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
En Mendoza, entre 1990 y 2011, la su perficie urbanizada superó el 40% ( Mesa & Giusso, 2014), extendiéndose en las zonas agrícolas del este y del sur provin cial, mientras que los departamentos de Capital y Godoy Cruz avanzaron hacia el oeste, sobre el piedemonte andino. Esta expansión urbana de baja densidad, en búsqueda de tierra de menor precio o de mayor calidad ambiental, generó los problemas característicos de las ciudades dispersas. Pese a la Ley 5804 de Ordena miento del Piedemonte, la urbanización desordenada sigue avanzando sobre esta unidad geomorfológica.
El piedemonte es un sistema muy vul nerable no solo por la extracción de la vegetación, de áridos y creación de repo sitorios a cielo abierto, sino también, por la alteración del balance hídrico entre el ecosistema urbano y el no urbanizado. Constituye una extensa unidad geomor fológica que conecta el frente oriental de la Precordillera con la llanura al este. Altitudinalmente, varía entre los 2000 y los 800 m s.n.m. con una pendiente ge neral del 10-15%. Las precipitaciones de 200 mm/año se concentran en el perío do estival (70%); son intensas y de cor ta duración ( Martínez Carretero, 2010). Toda esta región está expuesta a riesgos naturales que la tornan vulnerable por el avance de la urbanización. De esta ma nera, la fauna, en particular algunas aves nativas, se ven forzadas a desplazarse a otros sitios en busca de condiciones fa vorables para su supervivencia.
A nivel global, hay abundantes eviden cias de la disminución de la diversidad de fauna y flora nativa e incremento de taxones exóticos por impacto de la urba nización ( Stagoll et al., 2010). Dentro de la trama urbana la densificación edilicia, tráfico vehicular, peatonal, cambios en el uso del suelo, en el verde urbano y de oferta alimenticia, influyen en la biodi versidad por efectos múltiples y sinérgi cos. Cobra importancia la presencia de bosques urbanos implantados ( Fernán dez-Juricic, 2004) que introduce cambios en la oferta de hábitat. Así, se entremez clan la malla gris y la verde que se super ponen en el espacio. En una primera eta pa, en Mendoza el verde urbano quedó conformado por el perfil constituido por vereda, acequia, arbolado, calle, arbola do, acequia y vereda, posteriormente se agregaron plazas como lugares seguros durante los sismos ( Bochaca, 2005).
Se ha considerado a las aves como in dicadores de la calidad del hábitat urba no ( Fernández-Juricic, 2004; Sandstrom et al., 2006). Si bien, la mayoría de estos estudios se realizaron en el hemisferio norte, hay aportes significativos latinoa mericanos que permiten establecer com paraciones, que señalan diferencias con otros continentes ( Mac Gregor-Fors & Escobar 2017), lo que evidencia la nece sidad de evaluar el carácter indicador de las aves en la región.
En este trabajo se busca definir indica dores del impacto generado por el avan ce urbano en el piedemonte mendocino utilizando a las aves por su sensibilidad a los cambios. Se hipotetiza que se puede utilizar un grupo de aves nativas como un indicador válido del efecto de la ur banización.
Material y Método
Para poner a prueba la hipótesis se com para un sector urbano con otro de urba nización consolidada y no consolidada en el piedemonte.
El área urbana (UR) (646 ha), por su heterogeneidad, se dividió en 9 sectores más pequeños para su análisis, a fin de registrar diferentes situaciones. En di chos sectores hay presencia de grandes arboledas alineadas, áreas verdes, calles asfaltadas y diferente intensidad vehicu lar y peatonal ( Figura 1).
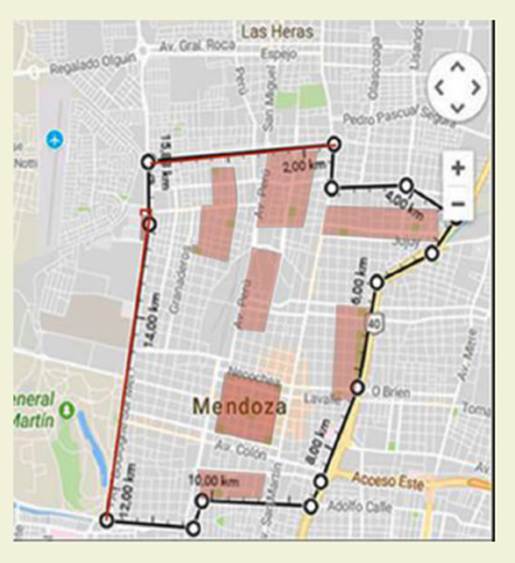
El tramo del piedemonte seleccio nado (646 ha) se extiende en sentido Este-Oeste. Presenta un sector ubicado junto a la ruta Panamericana con urba nización consolidada, calles, locales comerciales, arboledas adultas de árboles exóticos implantados, con remanentes de vegetación espontánea nativa en las primeras hectáreas. Los últimos sectores del área pedemontana presentan aún ve getación autóctona, sin presencia de ve getación exótica implantada, excepto en muy pequeños y escasos sectores, donde crecen árboles menores a 6 años. El área No consolidada (PNC) no presenta ur banización o es muy incipiente y cuenta con vegetación autóctona. Para facilitar el estudio del piedemonte, se dividió en subáreas, tres en el Piedemonte Conso lidado (PC) y dos en el No consolidado (PNC) ( Figura 2).
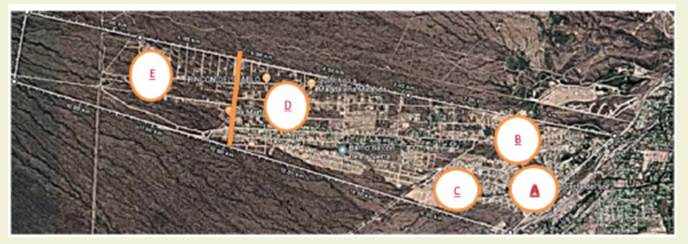
Figura 2: Piedemonte consolidado (A, B, C) y No consolidado (D, E). Figure 2: Consolidated (A, B, C) and unconsolidated piedmont (D, E)
Para estimar los impactos que produce la urbanización se utilizó una matriz de tres dimensiones: Presencia de verde, de Infraestructura gris y de Amenazas para aves.
Dentro de la dimensión Verde se con sideraron 7 variables: la existencia de plazas, parques y muchos sectores ver des, parques lineales, calles arboladas, arboles de más de 30 años, vegetación nativa, presencia de todos los estratos de vegetación. Estas variables se seleccio naron en base a la conspicua bibliogra fía que señala la importancia de la pre sencia, composición, estructura, edad y distribución del verde en la estructura urbana ( Chace & Walsh, 2006; Fernán dez-Juricic & Jokimäki, 2001; Perepelizin & Faggi, 2006; Ortega-Álvarez & Mac Gregor-Fors, 2009).
En la dimensión Infraestructura Gris ( Leveau & Leveau, 2004; McGregor-Fors & Schondube, 2011; Faggi & Caula, 2017), se consideraron 5 variables: espacios pa vimentados, construcciones de más de 30 años, edificios horizontales , residencia les , locales comerciales. En la dimensión Presencia de amenazas, se registraron 6 potenciales amenazas: accidentes por ani males domésticos, tránsito peatonal, trán sito vehicular, infraestructura, contingen cias climáticas y desmontes.
Las variables fueron estimadas en una matriz de impactos, valorándolas según una escala de 1 (menor presencia) a 5 (mayor presencia) ( Tabla 1).
Tabla 1: Valoración estimada de las variables estudiadas según tres dimensiones: Infraestructura Verde, Gris y Amenazas para las aves. Table 1: Assessment of the studied variables according to three dimensions: Green, Gray Infrastructure and birds Threats
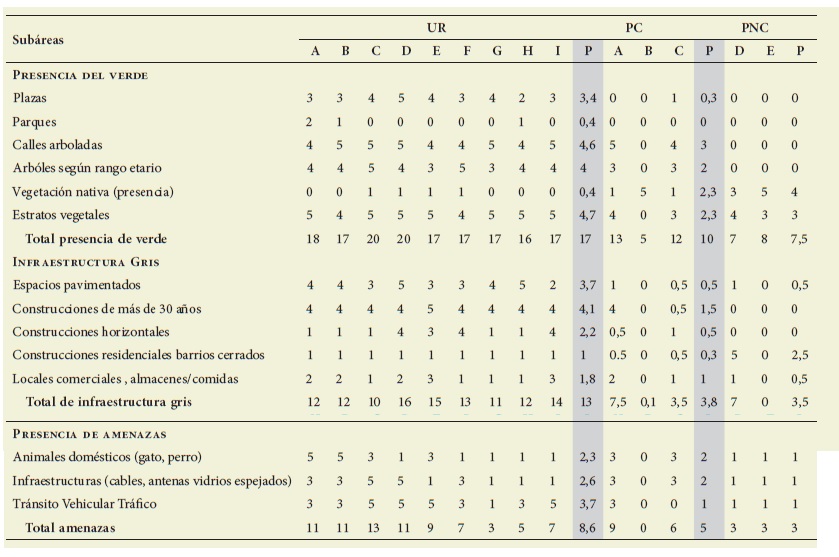
UR: Urbano, PC: Piedemonte Consolidado, PNC: Piedemonte No Consolidado
Al mismo tiempo, se estimó la rique za florística y de aves (número de espe cies), composición vegetal y abundancia de aves a lo largo de transectas sobre las cuadras de las manzanas. En el pie demonte no consolidado para relevar la vegetación se utilizó el método de Point quadrat modificado ( Passera et al., 1989), empleando 40 transectas de 200 m con lecturas cada 30 cm, ubicadas aleatoriamente, separadas 200 m entre sí. Para el relevamiento y análisis de la avifauna se siguió a Ralph et al. (1996). Se recorrie ron las mismas transectas, en un ancho de 30 m, durante las primeras horas de la mañana y las últimas horas de la tarde, registrando las aves vistas o escuchadas, observando aleros, techos, antenas, sue lo, ramas o troncos de árboles, sitios de nidada, refugio y alimento.
Las aves se relevaron con ayuda de guías de identificación ( Narosky-Yzurie ta, 2010), y se clasificaron según gremios de alimentación y de hábitat. Mediante la presencia-ausencia de aves indicadoras se estimaron los impactos.
Resultados
La Tabla 1 muestra comparativamente las características de las subáreas del sec tor UR, PC y PNC, según la presencia de verde, de infraestructura gris y potencia les amenazas para las aves.
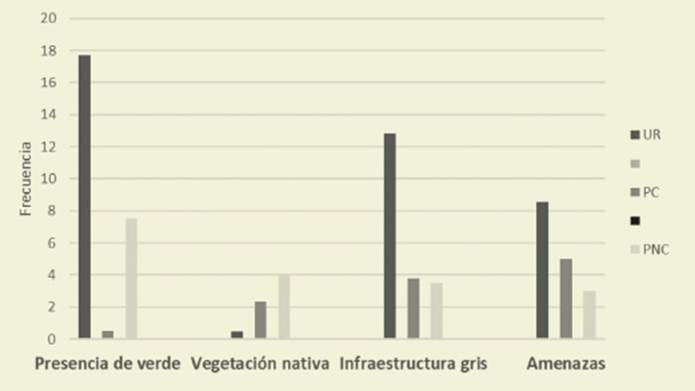
La presencia de la infraestructura ver de (IV) y gris (IG) disminuye según el gradiente urbano (IV: 17,7; IG: 12,8); piedemonte consolidado (IV: 10; IG: 3,77) y no consolidado (IV: 7,54; IG: 3,53). En contraposición, la presencia de vegeta ción nativa crece de manera inversa a ese gradiente y alcanza un valor promedio de 4 en el PNC. Las estimaciones de las amenazas para las aves fueron similares en UR (8,56) y PC (5,01) y menores en PNC (3).
Respecto a los indicadores de impac tos urbanos se observa mayor infraes tructura verde en UR (17,7) que en PC (10) y PNC (7,54). La cobertura vegetal autóctona en UR es escasa, solo alcanza un valor de 0,44, mientras que en PC fue de 2,33 y en PNC de 4. La infraestructura gris en la zona urbana fue mayor (12,8), comparada con la del PC (3,77) y de PNC (3,53), debido a que el piedemon te se encuentra en proceso de construc ción. Las estimaciones de las amenazas para las aves fueron en UR (8,56) y PC (5,01) y menores en PNC (3) ( Figura 3).
Las plazas predominaron en el sector urbano (UR: 3,44) a respecto a PC (0,2) y PNC que carece de ellas. Los valores de parques y verdes introducidos fueron si milares para UR (3,89) y PC (3,67). No así en PNC (0,5), al igual que calles ar boladas: UR (5), PC (4), PNC (0,9). Los árboles de más de 30 años predominaron en UR (4), mientras que la vegetación nativa fue más abundante en PNC (5), intermedia en PC (1,5) y mínima en UR (0,89) ( Figura 4).
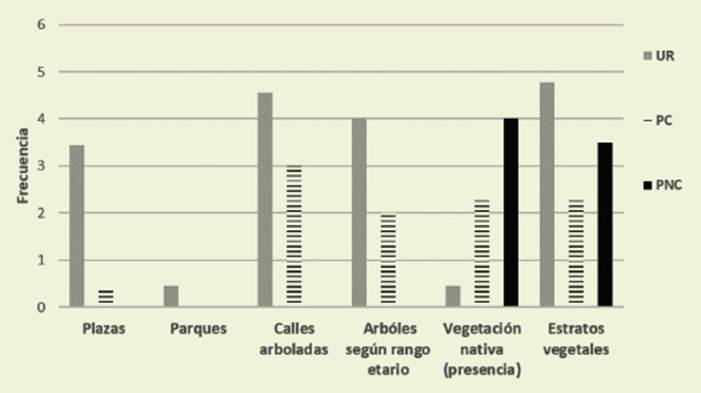
Figura 4: Estimación de la dimensión Presencia de verde en las zonas urbanas (UR) y en pie demontes consolidado (PC) y no consolidado (PNC). Figure 4: Estimation of the dimension Presence of green in urban areas (UR), in consolidated (PC) and unconsolidated (PNC) piedmont
La estratificación de la vegetación fue mayor en PNC, intermedia en UR y menor en PC.
Los ataques por animales domésticos fueron mayores en el sector urbano (UR 4,6), intermedio en PC (3,7) y escasos en PNC (0,5) ( Figura 2). Las potencia les coaliciones con edificios se estimaron como más abundantes en UR (4), inter medias en PC (2,3) y menores en PNC (0,5). Otras amenazas como las contin gencias climáticas, tránsito vehicular y peatonal y desmonte disminuyeron se gún el gradiente de urbanización.
La riqueza específica de vegetación exótica (V) y aves nativas y exóticas (A) disminuyó según el gradiente urbano -piedemonte no consolidado, con valo res para UR: 111 (V), 61 (A), PC: 56 (V), 52 (A) y PNC: 9 (V), 20 (A), inverso a la vegetación autóctona UR (11), PC (15), PNC (50) ( Tabla 2).
Tabla 2: Características de las áreas estudiadas en base a riqueza florística y riqueza y abun dancia de aves. Table 2: Characteristics of the studied areas based on floristic richness, richness and abundance of birds
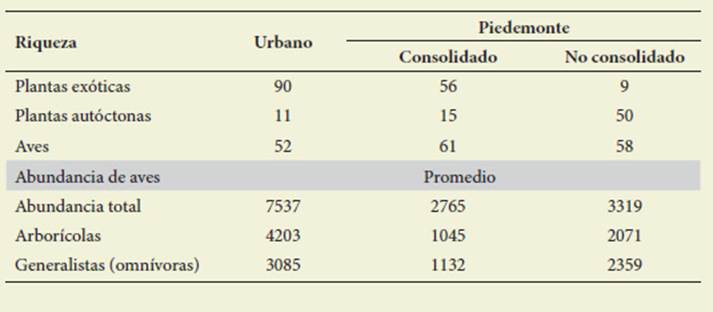
La distribución de la abundancia de aves presentó un pico en el sector urba no, seguido del piedemonte no consoli dado y un mínimo en el consolidado. Las aves generalistas y arborícolas siguieron la misma tendencia ( Tabla 2).
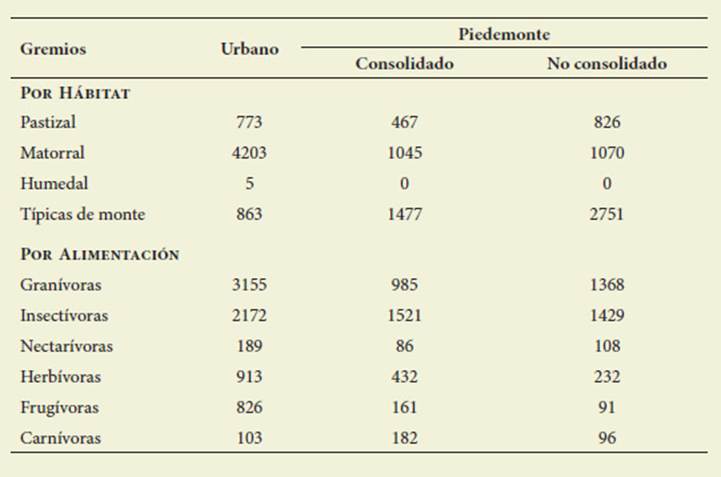
En UR la abundancia de las aves ar borícolas superó (4203), a las de monte (863) y de pastizal (773). En PC y PNC predominaron las aves típicas de monte (PC 1477, PNC 2751), seguidas de aves arborícolas (PC 1045, PNC 1070) y de pastizal (PC 467, PNC 826) ( Figura 5).
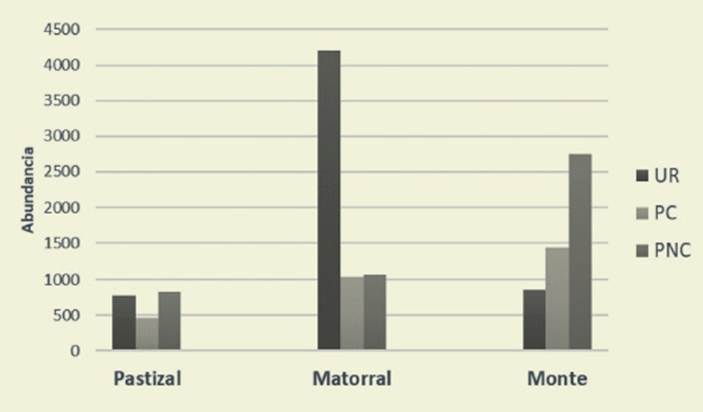
Figura 5: Abundancia de aves según tipos de hábitats. UR: Urbano, PC: Piedemonte Con solidado, PNC: Piedemonte No consolidado. Figure 5: Birds abundance by habitat guilds. UR: Urban, PC: Consolidated piedmont, PNC: Unconsolidated piedmont
Respecto a la alimentación, en UR pre valecieron las aves granívoras y en el piedemonte las insectívoras ( Figura 6, Tabla 3).
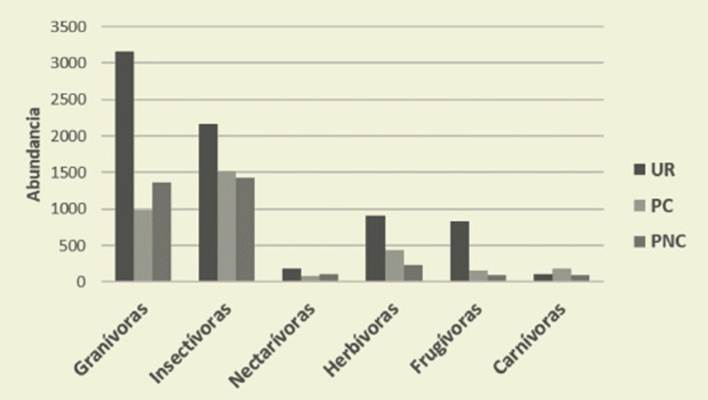
Figura 6: Abundancia de aves por gremios de alimentación. UR: urbano, PC: Piedemonte Consolidado, PNC: Piedemonte No consolidado. Figure 6: Birds abundance by feeding guilds. UR: urban, PC: Consolidated piedmont, PNC: Unconsolidated piedmont
Tabla 3: Abundancia de aves según gremios de hábitats y de alimentación. Table 3: Abundance of birds according to habitats and feeding guilds
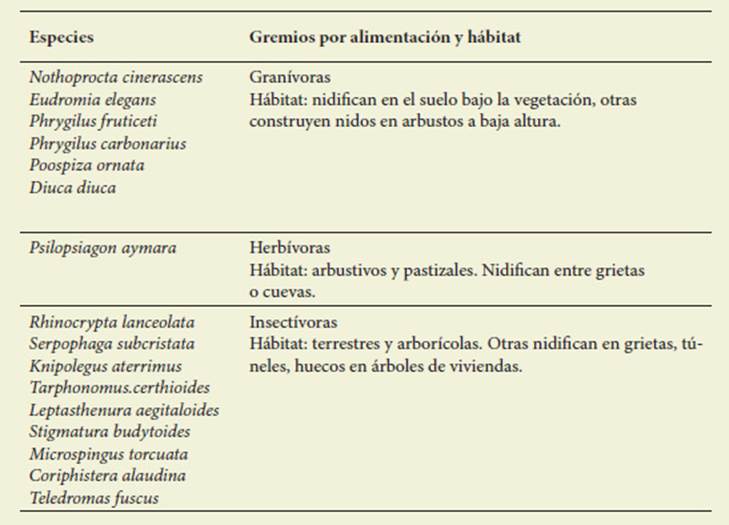
En el Piedemonte no consolidado se observaron 16 especies granívoras, her bívoras e insectívoras que por su sensi bilidad a los cambios se proponen como indicadoras ambientales ( Tabla 4).
Discusión
Los resultados de riqueza, abundancia, hábitat y alimentación, contrastados con la existencia de infraestructura verde, gris y potenciales amenazas permiten, a través de la respuesta de las aves, identifi car 16 aves exclusivas del piedemonte no consolidado y sensibles al impacto ge nerado por el avance urbano. Estas aves pertenecen a los gremios de granívoros que nidifican en el suelo, insectívoros que cavan túneles y nidifican en parte bajas de arbustos y perchan en árboles y arbustos ( Fraga & Narosky, 1985; Cane vari et al., 1991; Mezquida, 2002; De La Peña, 2013). Se comprueba así la hipóte sis de que un grupo de aves nativas pro pias del piedemonte puedan ser usadas como indicadores de efectos producidos por la urbanización.
En la ciudad de Mendoza, el avance de la urbanización implicó la introducción de especies vegetales exóticas (riqueza florística urbana 90 vs. 21 en el piede monte), lo cual explica el aumento de la riqueza y abundancia de aves urbanas, como ocurre en otras ciudades ( Chace & Walsh, 2006; Ortega Álvarez & MacGre gor-Fors, 2009) ( Tabla 1). Hay mayor cantidad de parques y plazas, arbolado denso, arboles maduros y todos los es tratos de vegetación están presentes. La variedad de oferta de paneo verde y fru tos repercute en la presencia llamativa de granívoras que puedan pastar en los céspedes de las distintas tipologías de espacios verdes, también en las frugívoras urbanas, las cuales quintuplican el nú mero del sector periurbano consolidado y fueron 9 veces más abundantes que en el no consolidado.
Otro indicador de impacto urbano es la presencia de infraestructura gris con valores mayores en zonas urbana que en piedemonte. En el piedemonte todas las construcciones son bajas y cubren las su báreas del este que lindan con ruta Pana mericana. Hacia el oeste la urbanización está en plena expansión, sin embargo, queda una zona importante sin edificar que garantiza hábitat silvestre para man tener la biodiversidad.
Los resultados del relevamiento de aves en el sector urbano indican una riqueza específica que casi triplicó el número de especies del piedemonte no consolidado con altos valores de las ge neralistas. También favoreció, aunque de manera leve, a las arborícolas, como Turdus amaurochalinus y T. rufiventris, Passer dometicus, Pseidoseisura lophotes, Columba livia, Patagioenas maculosa, con detrimento de las típicas de monte como Nothoprocta cinerascens, Eudro mia elegans, Zonotrichia capensis, Lep tasthenura aegitaloides, Phrygilus frutice ti, Microspingus torcuatus.
En cuanto al hábitat, la avifauna urba na en la ciudad de Mendoza es mayor mente arborícola y utiliza los árboles de los espacios verdes y de alineación para perchado, nidificación y alimentación, tal como ocurre en otras ciudades la tinoamericanas ( Faggi & Caula, 2017; MacGregor-Fors et al., 2017). Entre las arborícolas se observan aves genera listas; son los “explotadores” urbanos ( MacGregor-Fors & Schondube, 2011) y pueden llegar a expulsar a las aves autóc tonas, como ocurre con Passer domesticus, Columba livia y Storninus vulgaris. Otro grupo de aves son las “adaptadas” urbanas, que en general son omnívoras y nidifican en árboles. También utilizan elementos de la infraestructura gris que brinda espacios para posarse, dormir y nidificar ( Faggi & Caula, 2017). En contraposición, como era esperable en zonas urbanas, las aves de pastizal se registraron con bajos valores (UR 773), porque no encuentran en la ciudad de masiado sustrato para nidificar, ni sufi ciente material para la construcción de sus nidos.
Otros indicadores de impacto urba no son las amenazas que pueden afectar a la avifauna. En general, se producen muchas muertes de aves, como el caso de Columba livia, por diversas razones como ataques por parásitos de Passer do mesticus ( Gondim et al., 2010). En las zo nas urbanas de Mendoza se observa pre dación por animales domésticos, como gatos y perros, de Passer domesticus y Turdus amurochalinus. Otra amenaza la constituyen las colisiones contra edifi cios y atropellos por el tránsito vehicular ( Agudelo-Álvarez et al., 2010; Oviedo, 2014).
Coincidentemente con Mc Gregors Fors & Escobar-Alvarez (2017), para aves urbanas, la presencia de infraestruc tura gris no es problemática; por el con trario, la utilizan como lugares para nidi ficar. La avifauna urbana pertenece a las aves adaptadas por su gran plasticidad a los cambios atribuibles a su sistema nervioso ( Gil & Brumm, 2014), pudien do utilizar edificios para nidificar; en respuesta a las experiencias adquiridas, permanecen las que son más tolerantes a la perturbación humana o vehicular que les permite ser más flexibles para inva dir sitios urbanos (Sol & Lefebvre, 2008; Carrete & Tella, 2011). Las palomas, con cerebros más grandes, son tolerantes a la presencia humana y se adaptan a cam bios ambientales, mientras que las aves del piedemonte, de cerebro más peque ños, son más esquivas.
Las aves de pastizal predominaron en el piedemonte no consolidado PNC (826) debido al hábitat que ofrece mato rrales y pastizales nativos (Codecido & Bilenca, 2007). Las aves típicas de mon te fueron también más abundantes en el piedemonte no consolidado (PNC 2751) por la presencia de un hábitat cercano al natural; mientras que en PC fue menor (1477) porque el avance de las construc ciones hace que el piedemonte consoli dado se asemeje al hábitat urbano.
Conclusiones
El piedemonte sufre cambios en el há bitat a medida que la urbanización se extiende, especialmente por transforma ción de la cubierta vegetal nativa lo cual repercute en la desaparición de muchas especies de aves indicadoras del ambien te de la ecorregión Monte. Se identifican 16 especies de aves que pueden ser utili zadas como indicadoras de la condición ambiental. Ante el avance urbano en el piedemonte, y para impedir la pérdida de las aves sensibles, se debe fomentar el uso de plantas nativas en jardinería, don de la práctica de la xerojardinería es re comendable ( Dalmasso et al., 2009). Se aconseja implantar Schinus areira, Proso pis spp., Geoffroea decorticans, Cercidium praecox, Larrea spp ., Zuccagnia punctata, Atamisquea emarginata, Ximenia ameri cana, Condalia microphylla, Lycium spp ., Atriplex lampa y pastos como Trichloris crinita, Paspalum spp. , Stipa ichu, Aris tida mendocina, que además de brindar semillas pueden ser utilizadas en la cons trucción de nidos (Mezquida, 2003).

