











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
La soja, Glycine max L., es uno de los cultivos que más sufre ataques de plagas animales, principalmente insectos (Aragón, 2002). Dentro de las plagas insectiles, este cultivo es atacado por una gran diversidad de especies de larvas de lepidópteros conocidos como orugas defoliadoras tanto durante el período vegetativo como reproductivo. El principal daño que causan es la pérdida del área foliar lo cual incide en el rendimiento debido a la menor intercepción de luz, menor capacidad fotosintética, pérdida del material almacenado en la hoja y acortamiento del periodo de llenado de granos (Board et al., 2011). Esta situación ubica a la soja como el cultivo que requiere más insecticidas para su protección dentro de los cereales (maíz, Zea mays L. y trigo, Triticum aestivum L.) y las oleaginosas (girasol, Helianthus annus L. y colza, Brassica napus L.).
Como consecuencia, el desarrollo tecnológico para el control de plagas en este cultivo se ha profundizado. Un avance importante en los últimos años es el empleo de soja genéticamente modificada para expresar proteínas de la bacteria Bacillus thuringiensis (Berliner, 1915) para controlar algunas especies plaga. La soja Bt es una variedad que combina los eventos de transformación MON 87701 (que expresa la proteína Cry1Ac) y MON 89788 (tolerancia al glifosato) (Monsanto, 2019). En la actualidad, este evento, controla plagas primarias como la oruga medidora (Rachiplusia nu Guenée), la falsa medidora (Chrysodeixis includens Walker), la oruga bolillera (Helicoverpa gelotopoeon Dyar), la oruga de las leguminosas (Anticarsia gemmatalis H.) y otras de menor importancia (Urretabizkaya et al., 2010, Bernardi et al., 2012; 2014; Pomari et al., 2012). En Argentina, esta tecnología es de reciente incorporación en el mercado y su uso se está extendiendo año a año (Ybran y Lacelli, 2016). Es así como en la campaña 2018-2019 según datos del Sistema de Información Simplificado Agrícola (SISA) y el Catálogo de INASE (2019), más del 15 % de la superficie sembrada con soja en Argentina correspondió a variedades con el evento Bt.
Las plantas Bt son muy eficaces para evitar daños de sus plagas objetivo (dominantes en el nicho agro-ecológico), produciendo la disminución poblacional de las mismas. Este hecho, sumado a la reducción en el uso de insecticidas consecuencia de la adopción de esta tecnología y al posible efecto de estas plantas sobre los controladores biológicos de ciertas plagas, podría favorecer la ocurrencia de plagas no blanco de la tecnología Bt. Estas especies podrían aumentar su nivel poblacional y convertirse en plagas clave del cultivo (Bernardi et al., 2014).
En los últimos años, la ocurrencia de orugas del género Spodoptera está en aumento en el cultivo de soja, coincidiendo este incremento con el uso de la tecnología Bt (Vera et al., 2018). Entre las especies más frecuentes, podemos mencionar Spodoptera cosmioides (Walker), vulgarmente denominada como oruga del yuyo colorado u oruga militar grande. Esta especie es muy polífaga, afecta plantas cultivadas y malezas, y consume tanto estructuras vegetativas como reproductivas, lo que contribuye a su rápida adaptación a diferentes agroecosistemas causando daños en cultivos de importancia económica (Silva et al., 2016; Vera et al., 2018). Se la puede encontrar a menudo en cultivos de interés económico como: soja, maíz, arroz (Oryza sativa L.), girasol, sorgo (Sorghum bicolor (L.) Moench), algodón (Gossypium spp.), maní (Arachis hypogaea L.), alfalfa (Medicago sativa L.), avena (Avena sativa L.), hortícolas, ornamentales y otras. En Argentina, ha sido citada en las provincias de Chaco, Tucumán, Santa Fe, Entre Ríos y Buenos Aires (Kahl et al., 2016). En Brasil, fue recién luego de la adopción de la tecnología Bt que se observaron brotes poblacionales importantes de esta especie tanto en soja como en algodón (Luz et al., 2018; Conte et al., 2019).
La calidad nutricional de los alimentos es muy importante en los insectos fitófagos para su desarrollo y reproducción. Es por ello que su comportamiento y movilidad entre hospedantes estará determinada por la búsqueda y obtención de la cantidad adecuada de los nutrientes necesarios para su desarrollo y supervivencia (da Silva et al., 2017). Para el caso de larvas con poco movilidad, el sustrato en donde la hembra coloque los huevos será clave para el desarrollo de la progenie (da Silva et al., 2017). Cuando el sustrato no es óptimo a nivel nutricional, lo que se observa es un mayor número de estadios larvales. Es así como la planta hospedante utilizada como sustrato alimenticio en la etapa larval afecta los atributos biológicos de los adultos (Pashley et al., 1995; Awmack y Leather, 2002) y puede influir, por lo tanto, en la dinámica poblacional de las especies. Si bien este efecto en el desarrollo es más visible en insectos monófagos está demostrado en muchas ocasiones en insectos polífagos como S. cosmioides (Bavaresco et al., 2002, 2003, 2004; Cabezas et al., 2013).
Es así como resulta necesario conocer aspectos básicos de la biología de esta especie, tales como el tiempo de desarrollo, la fecundidad y la mortalidad asociados a la planta huésped y cultivos de interés, a fin de determinar el impacto que puede generar en los niveles poblacionales de esta especie de lepidóptero la adopción masiva de soja Bt y contribuir a implementar estrategias de control eficientes en el marco de un manejo integrado de plagas en el cultivo. En este contexto, el objetivo de este trabajo fue determinar parámetros de desarrollo de estados inmaduros de S. cosmioides criada en dos sustratos diferentes: Amaranthus spp. y soja Bt de una primera generación de individuos obtenidos del campo.
Materiales y métodos
Recolección de insectos
Para establecer una cría experimental de S. cosmioides, se realizaron recolecciones en campos comerciales de soja Bt en Tucumán, Argentina. Se ubicaron lotes con presencia de S. cosmioides y se procedió a recolectar larvas con la ayuda de paños verticales (más de 300 larvas). Las larvas se colocaron en cajas de Petri con hojas del cultivo como sustrato temporal, las cuales fueron acondicionadas en conservadoras refrigeradas y se trasladaron para su posterior cría en el laboratorio.
Cría en laboratorio
La cría se mantuvo en los Laboratorios de Terapéutica Vegetal (Facultad de Agronomía y Zootecnia-Universidad Nacional de Tucumán) con temperatura (25 ºC ± 2 ºC), humedad relativa (80 % ± 10 %) y fotoperíodo (12 h luz y 12 h oscuridad) controlados. Las larvas traídas del campo fueron acondicionadas en forma individual en cajas de Petri con hojas de soja Bt hasta que completaron el desarrollo y se formó la pupa. Las pupas obtenidas fueron colocadas en una jaula de cría revestida con papel sulfito como sustrato para la oviposición. Los adultos fueron alimentados con una solución de miel al 10 % embebida en algodón y colocada sobre un recipiente de plástico. Una vez que se produjeron las cópulas, se recolectaron las posturas, que fueron colocadas en cajas de Petri hasta la obtención de neonatas, las cuales fueron utilizadas para los bioensayos.
Obtención de plantas para los bioensayos
Para la alimentación de las larvas se utilizaron dos sustratos: hojas de soja Bt y hojas de Amaranthus spp. Las hojas de soja Bt se obtuvieron de plantas de la variedad M6410 IPRO sembradas en macetas de plástico negro de 1 L de capacidad. Una vez que las plantas alcanzaron un tamaño de 30-40 cm o más de 4 hojas, se cortaron las hojas en trozos de aproximadamente 1 cm x 3 cm para luego ser utilizadas para alimentar a las larvas. Las hojas de Amaranthus spp. se recolectaron de plantas que crecían en el predio de la Facultad de Agronomía y Zootecnia, Finca El Manantial, sin ningún tipo de manejo agronómico. Al igual que en soja, se cortaron las hojas en trozos de aproximadamente 1 cm x 3 cm para alimentar a las larvas.
Determinación de la duración de los estadios larvales
Para determinar la duración de los estadios larvales, se utilizaron neonatas de S. cosmioides a partir de huevos de los adultos obtenidos de las larvas recolectadas a campo. Se trabajó con una cohorte de 200 larvas, 100 para cada sustrato, las cuales se acondicionaron de forma individual en tubos de ensayos con tapón de algodón y el sustrato correspondiente durante los primeros estadios larvales. Para los últimos estadios larvales se las colocó en cajas de Petri debido a su mayor tamaño. La ocurrencia de mudas y mortalidad larval fue registrada diariamente en cada tubo o caja de Petri. Mediante microscopio estereoscópico se buscó la presencia de la cápsula cefálica en el tubo de ensayos o caja de Petri como indicio que la larva había mudado (Zenker et al., 2007). Una submuestra de las cápsulas cefálicas para cada uno de los estadios larvales y sustratos se conservó para posterior medición. Las hojas se cambiaron cada 48 horas. Se consideró finalizado el estado de larva cuando la misma dejó de alimentarse y de moverse y entró en un estado de aletargamiento, definido a veces como pre-pupa (Specht y Roque-Specht, 2016).
Determinación de la masa pupal y duración del estado de pupa
La duración del estado de pupa se determinó como el tiempo transcurrido desde el inicio de formación de la pupa hasta la emergencia de adultos. Las pupas obtenidas de los bionesayos fueron pesadas con una balanza de precisión a las 48 h y se determinó el sexo mediante observación de las aperturas genitales. Esto fue posible debido al dimorfismo sexual que presentan las pupas (Specht y Roque-Specht, 2016). Una vez determinado el sexo se las colocaron en forma individual en recipientes plásticos (1 L) tapados con voile sujetado con bandas elásticas hasta la emergencia del adulto.
Determinación del porcentaje de emergencia de adultos y relación de sexos
El número de adultos emergidos se utilizó para determinar el porcentaje de emergencia de adultos (número de adultos / número de pupas x 100), se confirmó el sexo asignado a cada individuo y se calculó la relación de sexos (número de hembras / número de hembras + número de machos).
Determinación de cambios en el tamaño corporal
Para la determinación de cambios en el tamaño corporal de la larva se midió el ancho de la cápsula cefálica (Bernardi et al., 2012). Para cada sustrato se realizaron montajes de una muestra de las cápsulas cefálicas conservadas de cada estadio larval en portaobjetos y se fijaron las cápsulas con esmalte de uñas transparente. La medición se realizó a partir de fotos tomadas en un microscopio y un programa específico para mediciones (ImageJ).
Supervivencia durante el desarrollo larval
Se determinó la supervivencia al estado larval mediante el registro de mortalidad diaria durante el desarrollo larval y el porcentaje de larvas que llegaron al estado adulto (número de adultos / número de larvas x 100).
Análisis de los datos
Para todas las variables medidas se procedió a verificar el cumplimiento de los supuestos de normalidad y homogeneidad de varianza con el fin de hacer un análisis paramétrico de los datos. Para duración de los estadios larvales no se verificó el cumplimiento de los supuestos de normalidad y homogeneidad de varianza aun cuando se procedió al modelado de varianza, por lo que esta variable se evaluó mediante un análisis no paramétrico de Mann-Whitney. Para duración del estado de pupa y masa pupal se verificó el cumplimiento de los supuestos de normalidad y homogeneidad de varianza, por lo que el análisis se realizó mediante ajuste a un modelo lineal general donde el sustrato brindado fue el factor fijo. Para la variable tamaño de cápsula cefálica también se verificó el cumplimiento de los supuestos de normalidad y homogeneidad de varianza, por lo que se realizó un ajuste a modelo lineal general. En todos los casos la significancia fue del 5 %. La relación de sexos se analizó dentro de cada sustrato mediante una bondad de ajuste a una relación 1:1 y mediante la prueba de independencia (Prueba exacta de Fisher) para determinar diferencias entre sustratos. Asimismo, se realizó un análisis de curvas de sobrevida de Kaplan-Meier.
Resultados
Duración de los estadios larvales
La duración de los estadios larvales presentó diferencias significativas entre los distintos sustratos para L2, L6 y L7 (Figura 1, Tabla 1). Para Amaranthus spp. y soja Bt, el número de individuos que completaron el estado larval fue de 20 y 19 respectivamente. Asimismo, el desarrollo en soja Bt involucró estadios larvales adicionales en algunos individuos, lo que llevó a que en soja Bt la duración del estado larval duplicara a la de Amaranthus spp. (Tabla 1, Figura 2).
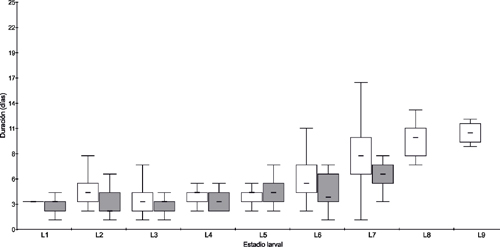
Tabla 1. Medianas (N) de la duración de los estadios larvales para los dos sustratos evaluados y valores de U para el test de Mann-Whitney.
/
*Solo para los individuos que completaron el desarrollo larvario y mudaron a pre-pupa.

Masa pupal y duración del estado de pupa
De los 20 individuos criados en Amaranthus spp. que completaron su desarrollo larval, solo 9 mudaron a pupa, mientras que la totalidad de los individuos criados en soja Bt mudaron a pupa. El resultado del modelo lineal para masa pupal mostró diferencias significativas entre soja Bt y Amaranthus spp. (F1,21 = 12,86; p = 0,0017), siendo las primeras más livianas que las segundas. La media en Amaranthus spp. fue 312,54 ± 15,49 mg (N = 9) y en soja Bt fue 239,86 ± 12,83 mg (N = 14). Para duración del estado de pupa las diferencias fueron no significativas (F1,22 = 4,25; p = 0,0514). La duración promedio en Amaranthus spp. fue de 14,14 ± 0,51 días (N = 7) y en soja Bt fue de 12,18 ± 0,57 días (N = 17).
Emergencia de adultos y relación de sexos
El porcentaje de emergencia de adultos fue de 77,7 % en Amaranthus spp. y de 94,44 % en soja Bt. En ambos casos, se observó predominio de hembras sobre machos (Chi2 = 12,5; p = 0,0004 en Amaranthus spp y Chi2 = 4,5; p = 0,0338 en soja Bt). Por su parte, la prueba exacta de Fisher mos-tró que no hay diferencias para la proporción de sexos atribuibles al sustrato (F = 0,27; p = 0,3521).
Cambios en el tamaño corporal
Las cápsulas cefálicas presentaron diferencias significativas atribuibles al sustrato donde se de-sarrollaron las larvas solo en L2 y L5 (Tabla 2).
Tabla 2. Medias del ancho de cápsula cefálica (mm) ± error estándar (N) para los distintos estadios larvales (L1-L8) para los dos sustratos evaluados y valores de T para la prueba de Student.
/
Supervivencia durante el desarrollo larval
El porcentaje de individuos que llegaron a adultos fue 7 % para Amaranthus spp. y 17 % para soja Bt. El análisis de Sobrevida de Kaplan Meier no mostró diferencias significativas entre los sustratos (Chi2 = 0,609; p = 0,4353, Figura 3).

Discusión
Este trabajo evaluó el desarrollo de estados inmaduros de S. cosmioides, una oruga con potencial de convertirse en plaga primaria de la soja Bt por ser plaga no blanco de los eventos de esta tecnología que se están cultivando en Argentina desde 2012, sobre su planta hospedante original Amaranthus spp., o yuyo colorado, y sobre soja Bt. Las variables evaluadas evidenciaron diferencias en el desarrollo entre ambos sustratos. La duración de los estadios larvales fue mayor en soja Bt en comparación con Amaranthus spp. Por su parte la masa pupal fue mayor en los individuos criados en Amaranthus spp. Para duración del estado de pupa y relación de sexos en los adultos emergidos, no se encontraron diferencias estadísticamente significativas entre los sustratos. El tamaño de las cápsulas cefálicas presentó diferencias estadísticamente significativas atribuibles al sustrato en algunos estadios. Los resultados muestran que la alimentación tiene una marcada influencia en la duración del ciclo de vida del insecto, acentuando la necesidad de conocer en mayor profundidad este impacto a fin de determinar el rol del yuyo colorado y la soja Bt en el crecimiento poblacional de esta especie de lepidóptero.
La duración de los estadios larvales fue más prolongada en soja. Bavaresco et al. (2003) observaron que S. cosmioides presenta un periodo larval más largo cuando es criada en soja no Bt, comparado con otros sustratos como cebolla, ricino y poroto, mientras que da Silva et al. (2017) obtuvieron resultados similares en soja comparado con algodón, avena, trigo, maíz y dieta artificial. En este trabajo, la duración del estado larva para Amaranthus spp. y soja Bt fueron de 23 y 40 días respectivamente, mostrando una diferencia de casi el doble. Asimismo, en soja Bt se observaron larvas que completaron el desarrollo presentando más estadios que Amaranthus spp. llegando incluso a L9. Bavaresco et al. (2003) obtuvieron resultados similares en soja no Bt con un 86 % de larvas con siete estadios y las restantes con ocho estadios; observando incluso diferencias notables con los otros sustratos evaluados (cebolla, ricino poroto). Por su parte, Cabezas et al. (2013) obtuvo mayor cantidad de estadios larvales en larvas alimentadas con hojas del árbol de aceite (Aleurites fortii Hemsl), sustrato que además presentó el ciclo larval más largo entre todos los sustratos evaluados en dicho estudio (Ricinus communis L. y Jatropha curcas L.). Claramente la ocurrencia de más estadios larvales y la mayor duración del ciclo larval en larvas alimentadas con soja, tanto Bt (nuestros resultados) como no Bt (otros autores), podría deberse a una menor adecuación nutricional de este alimento para su desarrollo. Este hecho parece ser independiente de la presencia de cristales Bt ya que Bernardi et al. (2014) no encontraron diferencias en distintos parámetros del desarrollo en larvas de S. cosmioides criadas en soja Bt versus soja no Bt (MON 87701 x MON 89788 y una línea genéticamente cercana respectivamente). En este sentido, es necesario realizar estudios con soja Bt y no Bt a fin de determinar si la presencia de los cristales Bt imparte un impedimento adicional para el desarrollo de poblaciones de Argentina de esta especie de lepidóptero o si se confirman los resultados obtenidos por Bernardi et al. (2014) para poblaciones del sur de Brasil. Asimismo, es interesante mencionar en este contexto el hecho que esta especie también se alimenta de las vainas, pudiendo compensar así posibles carencias nutricionales de las hojas de soja con lo cual un estudio completo que determine la adecuación de la soja como sustrato para el desarrollo de S. cosmioides debería incluir vainas.
Las pupas provenientes de larvas alimentadas con Amaranthus spp. fueron más pesadas que las pupas provenientes de larvas alimentadas con soja Bt. Un estudio donde se evaluó el desarrollo de S. cosmioides y S. eridania en diferentes sustratos mostró que en ambas especies los individuos alimentados con soja presentaron las pupas más livianas dentro de los sustratos evaluados (da Silva et al., 2017), reforzando lo expuesto anteriormente sobre la posible deficiencia nutricional de las hojas de soja.
El porcentaje de emergencia de adultos fue superior en soja Bt con un 94.44 %, valor similar al reportado por Specht y Roque-Specht (2016) en dieta artificial. Por su parte, las pupas criadas en Amaranthus spp. también presentaron un valor alto indicando que los individuos que completaron el desarrollo larvario y llegaron a pupa tenían las condiciones necesarias para completar esa etapa. Sin embargo, hay que destacar la alta mortalidad en pre-pupa y la baja viabilidad durante el desarrollo larvario.
En cuanto a la relación de sexos, para ambos sustratos se observó una mayor cantidad de emergencia de hembras que de machos. Este resultado difiere de lo obtenido por otros autores (Bavaresco et al., 2003, 2004; Cabezas et al., 2013; da Silva et al., 2017) donde la proporción siempre fue cercana a 1:1. A pesar de esta discrepancia, la prueba de Fisher mostró que no existen diferencias atribuibles al sustrato, coincidiendo con lo expresado por Bavaresco et al. (2003, 2004); Cabezas et al. (2013); da Silva et al. (2017).
La medición del ancho de cápsula cefálica permitió observar el crecimiento de S. cosmioides en los sustratos evaluados. Para ambos casos, se verificó un mayor crecimiento en los últimos estadios larvales, en concordancia con Specht y Roque-Specht (2016) en dieta artificial. Asimismo, en L5 las cápsulas de las larvas criadas en Amaranthus spp. fueron más grandes que las criadas en soja, coincidiendo con la mayor masa de la pupa registrada en dicho sustrato.
En ambos sustratos la supervivencia de larva a adulto fue muy baja. En contraposición, Bernardi et al. (2014) obtuvieron valores superiores al 90 % en su trabajo con soja Bt y soja no Bt y Bavaresco et al. (2004) valores superiores al 75 % en su trabajo con dietas artificiales. En este sentido es posible que los bajos valores encontrados se puedan deber a algún tipo de estrés ambiental sufrido por las larvas en los primeros estadios del desarrollo.
Conclusiones
Este estudio muestra que la soja Bt resulta un sustrato menos apropiado para el desarrollo de S. cosmioides que Amaranthus spp. El menor peso de pupa registrado en soja Bt confirma la baja calidad nutricional de la soja Bt. El hecho que Amaranthus spp. haya resultado un sustrato más apropiado nutricionalmente es explicable debido a que se trata del hospedante natural donde existe una historia evolutiva común. Por su parte, el mayor tiempo para el desarrollo de las larvas en soja Bt supone un efecto negativo para la soja Bt, ya que al pasar S. cosmioides más tiempo en el estado de larva, puede causar daños significativos por defoliación en el cultivo. Este alargamiento en el desarrollo podría significar que esta plaga al ser polivoltina, presente menos generaciones por año cuando se desarrolla en soja, lo que determinaría un menor crecimiento poblacional.
En su conjunto, este estudio brinda información relevante sobre la biología de esta especie que junto con estudios que analicen la tasa de consumo, el nivel de daño ocasionado y el potencial reproductivo de S. cosmioides desarrollada en soja Bt permitirán estimar el impacto de este cultivo en el crecimiento poblacional de la especie y su potencial de convertirse en plaga primaria; permitiendo de esta forma contribuir a la estabilidad en el sistema de la tecnología Bt.

