











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkKEY POINTS
• The developmental and epileptic encephalopathy are a clinically and genetically heterogeneous group and it is often challenging to determine their genetic cause.
• Genetic testing is essential to prevent unnecessary and potentially harmful diagnostic procedures and manage ment, and to determine precision medicine strategies.
• A genetic diagnosis was made in 39% of the patients and approximately 42% of the detected variants were novel.
• We emphasize the importance of considering mosaic variants, copy number variants and hereditary forms when designing and interpreting molecular studies, in order to optimize diagnosis and management of these patients.
The International League Against Epilepsy (ILAE) defines epileptic encephalopathy as a condition in which the epileptic form activity itself contributes to severe cogni tive and behavioral impairments beyond what expected from the underlying pathology alone (such as a cortical malformation)1.
In 2017, the new concept of “developmental and epi leptic encephalopathy” (DEE) was introduced for those cases in which the developmental consequences arise directly from the effect of a genetic variant, in addition to the effect of the frequent epileptic activity on development2. The recognition of DEE is critical, as there is a treatable component with the potential to improve the developmen tal outcome and cognition. Optimal management depends on a number of factors, such as the age of the patient, comorbidities, current medications and, most importantly, the underlying etiology, which seems to be mainly genetic3.
The DEEs are a clinically and genetically heteroge neous group and it is often challenging to determine their genetic cause. Major advances in molecular diagnosis over the past decades have contributed to our increased knowledge on the underlying causes of DEEs and genotype-phenotype correlations and have led to a rapid increase in the discovery of genes across the human disorders4-6. However, most genomic data is reported from developed countries, whereas data from developing countries is underrepresented.
The search term “developmental and epileptic en cephalopathy” led to more than 250 entries associations in the Online Mendelian Inheritance in Man database (OMIM) demonstrating the wide genetic heterogeneity underlying the DEEs.
Next generation sequencing (NGS) is a relatively new technique, which allows the simultaneous study of hundreds of genes. It is being applied to genetic testing of diseases or syndromes that are caused by a single or by many different genes, and has the potential to detect causal variants, including de novo and familial variants associated with DEEs and, due to the variable phenotypic presentations of the latter disorders, to greatly improve molecular diagnosis7.
Genetic testing is essential to determine precision medicine strategies and to prevent unnecessary and po tentially harmful diagnostic procedures and management. A genetic diagnosis can also provide useful information on the natural history and prognosis of a disease and facilitate better targeted genetic counseling. Furthermore, it allows the subject and their family to enter gene-specific networks of families with the same condition3,8,9.
The aim of this study was to extend our knowledge of the genetic background of Argentine children with DEE seen at a public tertiary pediatric hospital, applying a targeted NGS panel designed to analyze 52 epilepsy candidate genes in a group of 31 DEE patients.
Materials and methods
Study population
We retrospectively analyzed 31 patients with DEE diagnosed and followed at the Department of Neurology of Hospital de Pediatría J. P. Garrahan, a public tertiary-care pediatric center, between 2019 and 2020. We included patients with DEE with age at onset younger than 12 months and an unknown etiol ogy after ruling out structural abnormalities. The occurrence of metabolic disorders and/or large chromosomal abnormalities were investigated according to the epileptic syndrome. The fol lowing study variables were analyzed: sex, age at onset, family history, seizure semiology and neurodevelopment. Standard electroencephalography (EEG) and/or video EEG recordings and magnetic resonance imaging (MRI) were reviewed. Epi leptic syndromes were diagnosed and classified according to the ILAE classification2,10.
This study was approved by our Institutional Ethics Com mittee. Written informed consent for the study was obtained from patients, parents and/or tutors.
Molecular studies
Genomic DNA was extracted from peripheral blood using Chemagic 360 (PerkinElmer, USA). A panel of the fifty-two genes (Table 1) most frequently associated with DEEs was de signed based on bibliographic references using a customized enrichment strategy (SureSelect XT Low Input, Agilent Tech nologies, USA) which was run following the manufacturer’s protocols. The obtained enriched libraries were sequenced for paired-end reads of 150 bp in a MiSeq sequencer using the MiSeq Reagent Kit v2 (300-cycles) (Illumina, USA) with a mean coverage depth of 250X per sample.

Table 1 Gene panel related to developmental and epileptic encephalopathies tested by next generation sequencing
After sequencing, raw data were processed using an in-house bioinformatic pipeline to detect single nucleotide vari ants (SNV) and copy number variants (CNV) in all 31 cases according to international guidelines, as previously published by our group11.
The high coverage-depth obtained allowed us to analyze the presence of mosaic variants. Alternate allele fraction (AAF) (mosaic variant reads/total reads) was calculated using the data generated by NGS. For autosomal variants and X-linked variants in females, a variant was considered possibly mosaic if the AAF was less than 36% or greater than 64% by NGS analysis, while AAF higher than 10% was used as a threshold to identify mosaic variants in X-linked genes in males12.
The presence of the genetic variants was confirmed by Sanger sequencing and/or array comparative genomic hy bridization (CGH) (SurePrint G3 8x60K, Agilent Technologies, USA), as necessary. Variant classification was performed according to the American College of Medical Genetics and Genomics (ACMG) Standards and Guidelines13. When patho genic or likely pathogenic variants were found, samples from the patient’s presumed biological parents were tested by Sanger sequencing to classify the case as being hereditary (present in one or both parents) or de novo variants (pres ent only in the patient). All novel variants were submitted to ClinVar NCBI database.
In patients with Dravet syndrome (DS) and Dravet like syndrome (DLS) without relevant single nucleotide variants, multiplex ligation-dependent probe amplification (MLPA) for the SCN1A gene was performed using SALSA MLPA probemix P137 (MRC-Holland, Amsterdam) according to the manufac turers’ instructions.
Results
This report includes genomic results obtained from 31 pediatric patients with DEEs with the following clinical phenotypes: DS (n:7), DLS (n:3), West syndrome (WS) (n:6), WS that evolved to Lennox-Gastaut syndrome (WS/LGS) (n:4), epilepsy of infancy with migrating focal seizures (EIMFS) (n:2), continuous spikes and waves during slow sleep (CSWSS) that evolved to LGS (n:1), LGS (n:1), myoclonic status in non-progressive encepha lopathy (MSNE) (n:1), myoclonic atonic epilepsy (n:1), EE with multifocal spikes (n:1) and unclassified epileptic encephalopathy (UEE) (n:4). Eighteen of the patients were female. Median age at symptoms onset was 4.8 months (range 0-12 months).
Relevant variants were detected in 12 cases (39%) and the main clinical characteristics of these patients are shown in Table 2. Of these 12 cases, five showed novel pathogenic or likely pathogenic variants, six were previ ously reported as pathogenic and one was classified as variant of unknown significance (VUS) (Table 3).


Table 3 Relevant variants identified by a multigenic next generation sequencing- panel for developmental and epileptic encephalopathies
Relevant single-nucleotide heterozygous variants were identified in the SCN1A (sodium voltage-gated channel alpha subunit 1), GABRG2 (gamma-amino butyric acid type A receptor subunit gamma2) and STXBP1 (syntaxin binding protein 1) genes, a mo saic variant in SCN2A (sodium voltage-gated chan nel alpha subunit 2) and one homozygous variant in SCN1B (sodium voltage-gated channel beta subunit 1). Additionally, a heterozygous deletion involving the SCN1A, SCN2A and SCN3A (sodium voltage-gated channel alpha subunit 3) genes was detected in one patient, and the most frequent triplet repeat expansion in the ARX (aristaless related homeobox) gene was observed in another one in hemizygous state (Table 3 and Fig. 1).
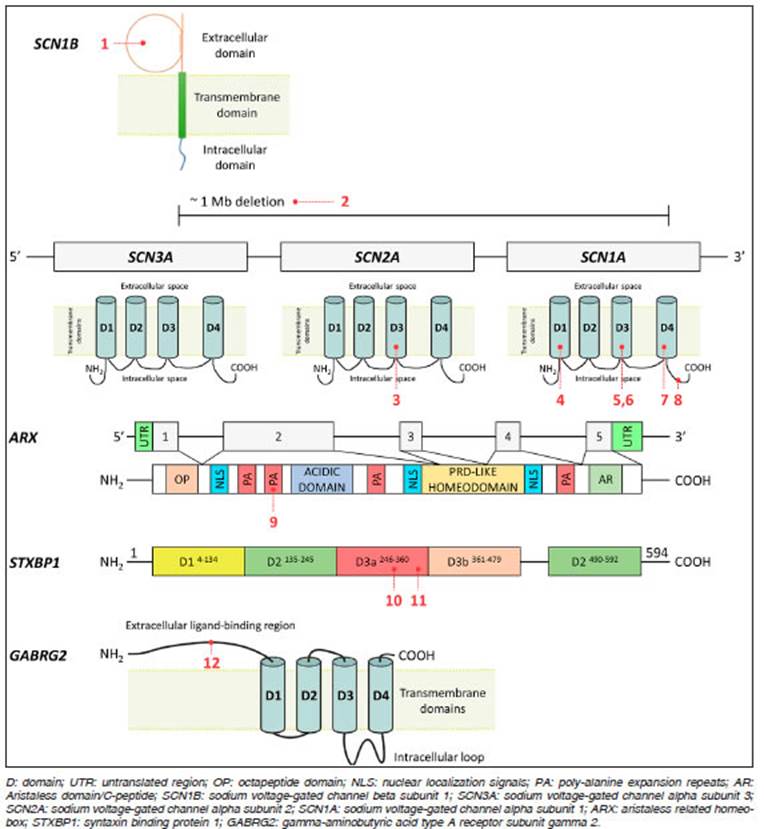
Fig. 1 Schematic representation of the genes, protein domains and associated variants detected (indicated with patient numbers)
Dravet and Dravet like syndrome
We identified pathogenic variants in the SCN1A gene in four cases with DS; one was missense, one was non sense and two were frameshift variants. One of them was novel while the remaining three had been previously reported14-16. All alterations were de novo.
Three of the four variants identified in the SCN1A gene generate a premature stop codon and are predicted to give rise to truncated proteins that affect normal function of the alpha subunit of the sodium channel Nav1.1. The variants detected in patient #4 (p.Cys336ValfsTer16) and patient #7 (p.Arg1636Ter) have been previously reported as associated with DS while the frameshift vari ant identified in patient #8 (p.Phe1831fsTer6) was novel. This latter variant was classified as pathogenic (ACMG criteria: PVS1, PM2, PM6, PP4). The variant c.4178 A>C p.(His1393Pro) detected in patient #6 is located within the extracellular loop, connecting the fifth and sixth segment in the third domain of the sodium channel protein Nav1.1, that involves the gate and voltage-sensing region of the channel. This pathogenic variant has been previously reported in a patient with DS14-16.
Finally, in patient #1 with DS we identified a novel homozygous missense variant in the SCN1B gene (p.Glu56Lys) affecting a highly conserved amino acid in the extracellular domain of the beta-1 subunit of the sodium channel. The variant was classified as likely patho genic (ACMG criteria: PM1, PM2, PP2, PP3, PP4). This variant is predicted to be probably damaging and disease-causing by PolyPhen-2 and Mutation Taster, respectively, suggesting that this novel variant might affect protein activity and function. Both parents are heterozygous for the variant and declared that there is no consanguinity within the family.
In the 3 patients with DLS no relevant variants were detected.
MLPA for CNVs in the SCN1A gene was performed in five patients without relevant variants and resulted nega tive in all of them.
Epilepsy of infancy with migrating focal seizures
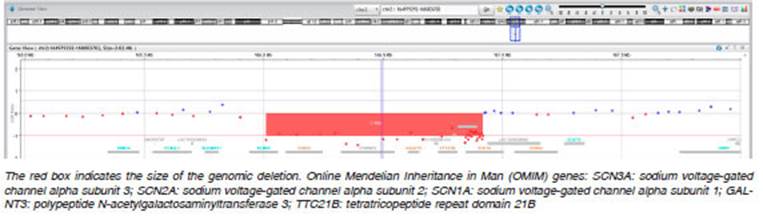
Two relevant novel variants were identified associated with sodium channels. In patient #2, we found a mosaic vari ant in the SCN2A gene (p.Val1340Ile) with approximately 17% of the AAF in leucocytes. This variant affects the fifth transmembrane segment of the third domain of the sodium channel protein type 2 subunit alpha, that involves the gate and voltage-sensing region. In silico tools suggest that this novel variant could be deleterious for protein activity and function. Secondly, in patient #3, we identified a CNV that affects the SCN3A, SCN2A and SCN1A genes. This variant was detected through a bioinformatics pipeline applied to the NGS panel and was confirmed by array- CGH (Figure 2). Both novel alterations were classified as pathogenic and their presence is in agreement with the clinical phenotype of the EIMFS patients9.
West syndrome/Lennox-Gastaut syndrome
We identified two previously reported variants in patients with WS that evolved to LGS, in the ARX and STXBP1 genes. In patient #9 we detected one of the most com mon polyalanine expansion mutations in the ARX gene c.315_335dup; p.(Ala109_Ala115dup), which is predicted to expand the original 16 alanine residues to 27 alanine residues in the first polyalanine tract of the homeobox protein ARX, a transcription factor required for normal brain development17. Since NGS studies have limita tions to detect triplet repeat expansions, this variant was confirmed by Sanger sequencing due to strong clinical suspicion. The variant was classified as pathogenic and was also present in the patient’s mother.
In patient #10 we identified a recurrent de novo vari ant in the STXBP1 gene (p.Arg292Cys) associated with WS18. The variant affects a highly conserved amino acid in the domain 3a of the syntaxin-binding protein and was classified as pathogenic.
Besides, we identified a previously reported missense variant (p.Leu135Phe) in the GABRG2 gene in a patient with LGS (#12). In silico tools strongly suggest that this variant could be deleterious for protein activity and func tion; however, we consider that there is not enough clinical evidence to confirm the deleterious impact of the variant and therefore we have classified it as VUS (ACMG criteria: PM2, PP2, PP3). At the same time, the variant was identi fied in his healthy father, indicating possible incomplete penetrance, as previously reported by other authors19,20.
Developmental and epileptic encephalopathy with a particular electroclinical pattern
We identified a novel variant in the SCN1A gene in patient #5 who had CSWSS that evolved to LGS. The variant (p.Ser1346Pro) detected in the SCN1A gene is predicted to affect a topological domain in the intracellular loop, between the fourth and fifth segment in the third domain of the Nav1.1 protein. It is predicted to be deleterious, prob ably damaging and disease-causing by SIFT/PROVEAN, PolyPhen-2 and Mutation Taster, respectively, suggesting that this novel variant might affect normal protein activity and function. It was absent in the mother’s sample while the father’s sample was not available. The variant was classified as likely pathogenic (ACMG criteria: PM1, PM2, PP2, PP3, PP5).
In patient #11, who presented with a myoclonic status, a presumably de novo missense variant (p.Cys354Arg) in the STXBP1 gene was identified. This variant has been previously reported as pathogenic and was associated with WS and Ohtahara syndrome21.
Discussion
Here we present a group of 31 pediatric patients with DEE studied at a public reference center in Argentina and analyzed with a customized NGS panel of 52 epilepsy-related genes. The incorporation of NGS analysis into our diagnostic approach led to a molecular diagnosis in 39% of the cases. These results are similar to those previously reported by our group, even though, with a different mo lecular strategy and a different group of patients, and are in agreement with those published in the literature, report ing diagnostic yields ranging from 10% to 48.5%3,11,22-26.
Approximately 66% of the patients in whom a molecular diagnosis was made showed a variant in genes that encode for ion channels, which agrees with previously reported data3,6,27,28. It is well known that pathogenic variants in sodium channel genes, including SCN1A, SCN1B, SCN2A, SCN3A, SCN8A and SCN9A, are responsible for a large proportion of cases with pediatric genetic epilepsies23, the three most common being SCN1A, SCN2A and SCN8A. This is especially true for DS, EIMFS and other DEE where the impact of the variants in the ion channels is relevant for selection of the most appropriate treatment.
In four of the seven patients with DS identified here, SCN1A variants were found. Although the functional ef fect of the variants is unknown, it is well established that the mechanism underlying epileptogenesis in SCN1A-associated DS is a loss-of-function (LoF) in sodium channels affecting inhibitory interneurons. Therefore, sodium-channel-blocking drugs should be avoided in DS29.
In addition, elucidation of the mechanisms responsible for SCN1A-related DS has stimulated innovative research into precision treatments to correct the underlying molecular or functional defect.
As expected, all the DS patients with a SCN1A muta tion presented with de novo variants. Nevertheless, we observed a very infrequent pattern of inheritance in a DS patient who had a homozygous variant in the SCN1B gene. To our knowledge, this alteration is the tenth homo zygous variation in the sodium channel accessory subunit gene reported in DEE patients30-32. Both parents of the patient harbor the heterozygous p.(Glu56Lys) variation and have no history of seizures.
No relevant variants were identified in 28% of the patients with DS, as was to be expected based on the reports in the literature, since SCN1A variants or deletions are found in 70-80% of tested DS cases14,33. A substantial minority of patients has no identified cause, and a smaller minority has a confirmed pathogenic variant involving a different gene, similar to our patient with the homozygous SCN1B variant.
In the patients with DLS no relevant variants were detected. In recent years, a large number of genes as sociated with DLS have been reported, but data are still insufficient to establish a genotype-phenotype relation ship; therefore, in this analysis we decided to include in our panel only those genes with a proven relationship with DS or DLS (SCN1A, SCN2A, SCN8A, SCN9A, SCN1B, PCDH19, GABRA1, GABRG2, STXBP1, HCN1, CHD2 and KCNA2)19.
In the EIMFS cases we identified a mosaic SCN2A variant and a CNV that involved the SCN1A, SCN2A and SCN3A genes, in agreement with previous studies describing SCN2A as the second-most frequent gene after KCNT1 involved in this syndrome9. In contrast to patients with SCN1A variants, neonatal and early infantile epilepsies related to SCN2A variants may show a good response to sodium-channel blockers, as did our patient with the mosaic SCN2A variant. This effect was explained by other authors through functional studies that showed that SCN2A-associated early-onset epilepsies are due to gain-of-function (GoF) variants with increased sodium channel activity and show a good response to sodium channel blockers, whereas variants found in late-onset forms have a LoF effect without a meaningful response to sodium-channel blockers34.
The mosaic SCN2A variant identified here supports the increasing number of patients with DEE associated with variants with somatic mosaicism9. It is important to be aware that a proportion of patients with DEE may have somatic mosaic variants that can easily be missed by exome sequencing, a technical approach that usually shows a lower depth of coverage than customized gene panels. Currently, it has been widely established that NGS is able to detect germline mutations present in all of the patients’ cells; however, somatic and mosaic variants can be difficult to identify. High-sequence depth, which is more commonly reached with targeted gene panels, aids in the detection of mosaic mutations with studies more commonly associated with somatic mutations relying on coverage >100X for accurate variant calling7,12.
Importantly, using an in house specific CNV-detection pipeline, we identified a CNV in a patient with EIMFS studied using the NGS panel, which seemed to involve the SCN1A and SCN2A genes. The application of array CGH allowed us to describe a 1Mb heterozygous deletion in patient #2 that involved not only SCN1A and SCN2A, but also the SCN3A gene, which was not included in our customized NGS panel (Table 1). The deleted region also includes the OMIM genes GALTN3 and TTC21B, associ ated with recessive diseases and thus cannot be related with the patient’s phenotype. NGS is increasingly being employed in the detection of CNVs using bioinformatic approaches, although a second method is necessary to confirm them and precisely determine the size of the in volved region. The development of effective bioinformatics pipelines employed in the detection of CNVs has added another layer of complexity to the applications of NGS for diagnostic testing7.
In the group of patients with WS that evolved to LGS, one had a heterozygous variant in the GABRB2 gene. Genes related to inhibitory GABA neurotransmitters play a key role in genetic epilepsies, including DEEs and genetic generalized epilepsies35. Genes that encode subunits of GABA(A) receptors, including GABRG2, GABRA1, GABRB3 and GABRD are part of the long list of epilepsy genes. A patient with LGS carrying a GABRB3 mutation was successfully supplemented with vinpocetine, an alka loid with pharmacological effects that include vasodilata tion, antioxidation, anti-inflammation, synaptic modulation and antithrombotic properties, leading to reduction of epileptiform activity on EEG recordings and improvement of cognitive, behavioral and language functions36.
The other patient with WS/LGS had several relatives with intellectual disability and epilepsy with a genealogy showing an X-linked pattern; therefore, an alteration in the ARX gene was suspected. The most common polyalanine expansion mutation in the ARX gene c.315_335dup; p.(Ala109_Ala115dup), was confirmed by Sanger se quencing.
A specific epilepsy syndrome was observed in one of the patients with a variant in the STXBP1 gene who developed myoclonic status epilepticus. Pathogenic variants in this gene are most commonly associated with Ohtahara syndrome, WS and non-syndromic intellectual disability with or without epilepsy, while their association with myoclonic status had not been previously reported37.
It should be noted that the number of negative results in patients with DEE in this cohort is in agreement with data reported by other authors3,6,7. Nevertheless, we emphasize the need of performing additional studies in depth to these patients, for example by exome sequenc ing, to reach a precise diagnosis and hopefully treatment optimization.
Furthermore, the diagnostic yield in our study was similar to that reported in the literature, although only the 52 genes most frequently associated with DEE were included. This demonstrates that a small group of genes are responsible for the majority of DEE, which is relevant when searching for an efficient diagnostic approach in countries with limited access to NGS panels or exome sequencing, such as Argentina.
In conclusion, using an NGS panel, a genetic diagnosis could be made in 39% of patients, including in those with rare genetic variants, such as one CNV and one mosaic alteration, and establishing new genotype-phenotype as sociations that have not been previously described. We emphasize the importance of considering mosaic variants, CNV and hereditary forms when designing and interpreting molecular studies in order to optimize the diagnosis and management of patients with DEE. Approximately 42% of the detected variants were novel, expanding the knowl edge of the molecular basis of DEEs in Latin-American patients. Molecular characterization is becoming the new diagnostic paradigm in DEE patients. Knowing the genetic cause of DEE will improve the treatment tailoring of some genetically determined forms of epilepsy.

