











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Los humedales son sitios de alta biodiversidad (Ramsar, 2006), que debido tanto a los disturbios naturales (fluctuaciones hídricas) como a los antrópicos (ganado, urbanización) son ambientes muy susceptibles a ser invadidos por especies introducidas (Raffaele, 2004; Zedler & Kercher, 2004; Gallardo et al., 2016). Esta susceptibilidad se refleja en el hecho de que un alto porcentaje (24%) de las plantas más invasoras del mundo son invasoras de humedales (Zedler & Kercher, 2004). Sin embargo, no todas las especies exóticas (introducidas) se convierten en especies naturalizadas o invasoras (Richardson et al., 2000) y es necesario entender la diferencia entre estos términos cuando se realizan inventarios de especies identificando nativas e introducidas.
El impacto que introducen diferentes especies varía enormemente y depende de la susceptibilidad de los ecosistemas a la invasión y de las características de la especie. Según Pysek et al. (2020), las especies invasoras afectan la riqueza y abundancia de las especies nativas, y aumentan su riesgo de extinción; además, alteran las redes tróficas y cambian los ciclos de nutrientes. También se ve afectada la hidrología, la estructura del hábitat y el régimen de disturbio, con lo cual todo el funcionamiento de los ecosistemas puede quedar transformado por la incorporación de especies invasoras. Sin embrago, no toda especie invasora tiene un efecto negativo sobre la comunidad en la que se naturaliza. Richardson et al. (2000) sugirieron el término transformadoras para las plantas invasoras que introducen cambios en el ecosistema en un área significativa dentro del ambiente invadido.
En la Patagonia andina existen numerosos cuerpos de agua temporales o semitemporales denominados localmente mallines o vegas. Estos humedales presentan vegetación en prácticamente toda su superficie, ya que suelen secarse durante los meses de verano. Son ambientes altamente productivos (Buono et al., 2010) y de mucha biodiversidad (Perotti et al., 2005; Jara et al., 2013). Se caracterizan por la presencia de especies vegetales adaptadas a las fluctuaciones del nivel del agua (hidrófitas), y la ausencia de aquellas no tolerantes a la condición de anegamiento producto de los pulsos de inundación. Suelen presentar una cobertura vegetal densa, dominada principalmente por plantas herbáceas de las familias Juncaceae, Cyperaceae y Poaceae (Iriondo et al., 1974; Raffaele, 1999). En este sentido, la zona litoral de los lagos, tanto profundos como someros que también abundan en la Patagonia, son ambientes con un régimen hídrico fluctuante. Esto le confiere características semejante a los humedales en cuanto a una vegetación hidrófita, con una producción primaria que puede ser alta en lugares reparados de la costa (Neiff, 1973). Por lo tanto, en este estudio llamaremos humedales tanto a los mallines como a la zona litoral de lagos.
El número de especies introducidas en humedales es alta (Hauenstein et al., 2008; Kutschker et al., 2014; Cuassolo & Diaz Villanueva, 2019; Manzo et al., 2020). Kutschker et al. (2014) encontraron un aumento de la cobertura de especies introducidas en humedales con mayor presión de ganadería, mientras que Cuassolo & Díaz Villanueva (2019) encontraron una relación positiva entre el porcentaje de especies introducidas y la distancia al centro de la ciudad de San Carlos de Bariloche. En el Parque Nacional Nahuel Huapi, el 25% de la flora terrestre ha sido identificada como especies introducidas (Ezcurra & Brion, 2005) y Speziale & Ezcurra (2011) hallaron que el proceso de invasión en la región está influenciado tanto por el gradiente de precipitaciones como la distancia a los centros urbanos y los disturbios antrópicos. Entre las especies introducidas encontradas por Cuassolo & Díaz Villanueva (2019) en mallines, Potentilla anserina L. (Rosaceae) (Fig. 1A) fue registrada en todos ellos (cuatro ambientes), alcanzando una cobertura casi del 50% en uno de los mallines (Fig. 1B) y se la identificó como posible invasora.
En la Argentina se han hallado cinco especies del género Potentilla; dos son nativas (P chiloensis L. Mabb. y P. tucumanensis Castagnaro & M. Arias) y tres introducidas (P anserina, P vesca (L.) Scop. y P recta (L.)) (Puntieri & Brion, 2005). En particular, P. anserina es una planta herbácea, hemicriptofita y estolonífera que fue hallada en Argentina por primera vez en 1877 en la provincia de Buenos Aires (Berg, 1877) y más tarde se registró en toda la Patagonia (Grondona, 1984). Manzo et al. (2020) encontraron que su abundancia en humedales de Chubut es mayor en sitios con concentración alta de fósforo total. Por otro lado, en Chile fue encontrada en lagunas costeras con alta conductividad (Hauenstein et al., 2008) y en mallines de Torres del Paine (Clausen et al. 2006).

Fig. 1: A: Foto de una planta de Potentilla anserina. B: Foto de la Laguna Fantasma durante su fase sin agua (verano), mostrando la distribución de las tres especies dominantes (c: Carex aematorhyncha, e: Eleocharis pachycarpa y p: Potentilla anserina).

Fig. 2: Mapa del Parque Nacional Nahuel Huapi mostrando los 28 sitios de relevamiento. Círculos verdes: ausencia; círculos rojos: presencia de Potentilla anserina. En el mapa de la República Argentina se detalla con círculo azul la ubicación del área de estudio.
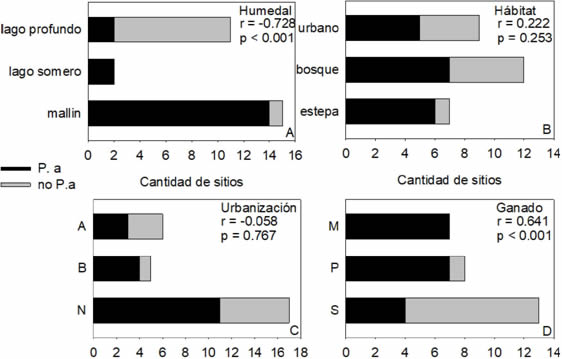
Fig. 3: Cantidad de sitios con presencia/ausencia de Potentilla anserina (P. a) según A) el tipo de humedal, B) el hábitat, C) el grado de urbanización (N: nula, B: baja, A: alta) y C) la presencia de ganado (M: mucho ganado, P: poco ganado, S: sin ganado). Se detalla en cada panel el número de r y el valor p de las correlaciones de Spearman para cada una de las categorías y la presencia de P anserina.
La presencia de P. anserina en los humedales puede tener impactos en el funcionamiento de los ecosistemas, como por ejemplo en el reciclado de nutrientes. Por un lado, por su capacidad de liberar más cantidad de materia orgánica disuelta (MOD) que las especies nativas con las que convive (Cuassolo et al., 2012; Cuassolo et al., 2016) y, por otro lado, porque su tasa de descomposición es mayor que las especies endémicas (Cuassolo et al., 2020). La presencia de micorrizas fue propuesta como una característica que le confiere ventajas adaptativas para crecer también en suelos con bajo contenido de fósforo, ya que se halló que concentra cantidades altas de este elemento en sus raíces (Cuassolo et al., 2012).
Los objetivos del presente trabajo fueron: a) analizar la distribución de P. anserina cubriendo un rango de precipitación media, ambientes e impacto antrópico y en diferentes tipos de humedal, desde las costas de lagos profundos y someros hasta mallines de bosque y estepa; b) determinar si la especie representa un riesgo para los humedales del Parque Nacional Nahuel Huapi. Para ello se identificaronallines con presencia de P. anserina y se midió su cobertura y la riqueza de especies (diferenciando nativas e introducidas) con el fin de determinar si existe una relación negativa entre estas variables.
Materiales y Métodos
Area de estudio
El estudio se llevó a cabo en el Parque Nacional Nahuel Huapi (PNNH), en el noroeste de la Patagonia Argentina (entre 40°58’ S - 41°21’ S y 71°08’ O - 71°49’ O, Fig. 2) en ambientes acuáticos temporales o semitemporales (mallines) y en la zona litoral vegetada de lagos someros y profundos. El clima de la región es templado frío, con precipitaciones medias anuales que van desde los 3000 mm en el Oeste hasta los 200 mm en el Este (Paruelo et al., 1998), concentradas en el otoño en forma de lluvias y en el invierno como lluvias y nevadas. Este gradiente de precipitaciones medias genera un gradiente de vegetación marcado. En el Oeste se desarrolla el bosque húmedo con un predominio de especies perennifolias, principalmente Nothofagus dombeyi (Mirb.) Oerst. (coihue), y arbustos como Luma apiculata (DC.) Burret (arrayán) y una bambusoidea nativa de America del Sur, Chusquea culeou É. Desv. (caña coligüe) principal componente del sotobosque Templado Húmedo. Por encima de los 1000 m snm domina la especie caducifolia N. pumilio Krasser (lenga). En el Este del gradiente estudiado se encuentra la estepa, con vegetación dominada por Festuca pallescens (St.-Yves) Parodi y una baja abundancia de arbustos, como Discaria chacaye (G. Don) Tortosa y la especie invasora Salix fragilis L. (Ezcurra & Brion, 2005). Entre la zona de bosque húmedo y la estepa se encuentra la ciudad de San Carlos de Bariloche, que corresponde a una vegetación de bosque mixto, con presencia de especies perennifolias como Maytenus boaria Molina (maitén), Lomatia hirsuta (Lam.) (radal) y la conífera Austrocedrus chilensis (D. Don) (ciprés de la cordillera).
Relevamiento
Durante los meses de verano (estación de crecimiento) de 2020 se realizó un relevamiento en 28 sitios a lo largo de una transecta Este-Oeste de 55,4 km y abarcando 37,1 km en sentido Norte-Sur (Tabla 1, Fig. 2). El relevamiento incluyó:
Tres tipos de humedal: a) costas vegetadas de lagos profundos b) costas vegetadas de lagos someros, y c) humedales temporales o semitemporales (mallines).
Tres tipos de hábitats: a) bosque, b) estepa y c) urbanos. Los sitios urbanos correspondieron a los incluidos en el ejido urbano de la Municipalidad de San Carlos de Bariloche con una densidad de viviendas (contabilizando el número de casas a una distancia de 100 metros, desde la zona periférica del humedal) mayor a 5 por ha (Cuassolo & Diaz Villanueva, 2019).
3) Tres categorías según el grado de urbanización: a) alta: son aquellos sitios dentro del hábitat urbano que presentan mayor grado de urbanización, sin protección. Estos ambientes se caracterizan además por el alto porcentaje de especies introducidas, b) baja: sitios en zonas con densidad de viviendas menor a 5 por ha, que conservan la vegetación nativa, (dentro del hábitat de bosque) y sitios dentro del hábitat urbano pero eclarados reservas naturales urbanas (RNU), con bajo impacto antrópico, y c) nula: sitios tanto de bosque como de estepa sin asentamientos urbanos.
4) Tres categorías según la presencia de ganado: a) sin ganado, b) poco ganado: menos de 20 animales a una distancia de 100 metros desde la zona periférica del humedal, mayormente equino, y c) mucho ganado, más de 20 animales a 100 metros desde la zona periférica del humedal, son sitios de pastoreo de ganado bovino y equino (observaciones personales).
Cobertura y riqueza
Además, se seleccionaron 8 mallines en donde se había verificado la presencia de P. anserina, cubriendo un rango de hábitat que abarcó bosque (3), urbano (3) y estepa (2), incluyendo en los de bosque el mallín de altura. En dichos mallines se realizó un muestreo de la flora para medir la riqueza de especies (tanto nativas como introducidas) y se midió la cobertura de P. anserina. En primer lugar, se realizaron muestreos de toda la flora encontrada en transectas (entre 5 y 9, dependiendo del tamaño del mallín) para determinar el número de especies (riqueza). En los mallines que presentaron ojos de agua permanentes (Teleférico y Los Patos) se trazaron transectas que atravesaron la zona de orilla, con lo cual en el listado se incluyeron las especies netamente hidrófitas. Para la determinación de las especies se realizaron herbarios de cada mallín los cuales se encuentran en el laboratorio de Limnología del Centro Regional Universitario Bariloche. La clasificación, nomenclatura y el origen geográfico de las especies se determinó según (Correa 1971-1998, Dimitri 1977, Zuloaga et al. 1994, Ezcurra & Brion 2005). Además, todas las especies fueron confirmadas con la base de datos actualizada de la Flora del Cono Sur del Instituto de Botánica Darwinion (http://www.darwin. edu.ar) y los autores de los taxones fueron corroborados de acuerdo a The International Plant Names (https://www.ipni.org.).
Luego de las identificaciones se realizó un muestreo en el cual se estimó la cobertura de P. anserina utilizando un cuadrado de 50 x 50 cm (0,25 m2) en transectas (entre 5 y 10, dependiendo del tamaño del mallín), que incluían las zonas centrales e intermedias de los ambientes. En cada cuadrado se estimó la cobertura de P. anserina y se extrapoló el área muestreada al área del mallín. Para calcular el área total de cada mallín se utilizaron imágenes satelitales (Google Earth).
Análisis de datos
La presencia de P. anserina fue analizada en relación con las variables categóricas (tipo de humedal, habitat, urbanización y ganado). Para analizar si existe una relación entre la presencia de P. anserina y las variables categóricas mencionadas se realizó un test no paramétrico (correlación de Spearman), que además relaciona a las variables categóricas entre sí. Para probar si existe correlación entre el porcentaje de cobertura de P. anserina y la riqueza de especies total, de introducidas y de nativas de los 8 mallines muestreados se realizó un test de correlación de Pearson. Para ambos análisis se utilizó el programa SigmaPlot versión 11.0.
Resultados
La presencia de P. anserina fue verificada en 18 de los 28 sitios relevados (Tabla 1, Fig. 2). Estuvo significativamente relacionada con el tipo de humedal (correlación de Spearman, r = 0,728, p < 0,001), ya que estuvo presente en casi todos los mallines analizados, tanto en ambientes de bosque como de estepa, con excepción del mallín Los Moscos, y en la zona litoral de los dos lagos someros. En los lagos profundos se observó en el lago Hess y en un sitio del lago Nahuel Huapi (Fig. 2 y 3A). El sitio en donde se halló del lago Hess corresponde a la boca del río Manso, en una zona con acceso al público, donde se realizan prácticas recreativas. La presencia en la costa del lago Nahuel Huapi corresponde al centro de la ciudad de San Carlos de Bariloche, el sitio con mayor impacto registrado en todo el perímetro del lago.
Con relación a los habitats, los sitios muestreados se encontraron mayormente en el bosque (12 sitios), mientras que 7 estuvieron en la estepa y 9 correspondieron a sitios urbanos (Tabla 1). En casi todos los sitios de estepa (86%) se encontró P. anserina, mientras que sólo se halló en la mitad de los de bosque y urbanos (Fig. 3B). Los sitios de estepa en los cuales no estuvo presente correspondieron a sitios en la costa del lago Nahuel Huapi. Sin embrago, el índice de Spearman indicó que no hay una relación significativa entre la presencia de P. anserina y el habitat (r = 0,222, p = 0,253).
De los 28 sitios analizados, 11 correspondieron a áreas con algún tipo de urbanización (6 urbanas y 5 periurbanas) y 17 a zonas sin urbanización (Tabla 1). Si bien la proporción de sitios con presencia de P. anserina fue mayor en las zonas periurbanas (80%), que en sitios sin urbanización (60%) (Fig. 3C), la relación entre estas variables no fue significativa (r = 0,058, p = 0,767).
Sin embargo, la presencia de ganado estuvo relacionada significativamente con la presencia de P. anserina (r = 0.641, p < 0,001). El ganado estuvo presente en 15 de los 28 sitios relevados (Tabla 1), de los cuales 14 (93%) tuvieron P. anserina, mientras que sólo se halló en el 30% de los sitios sin ganado (Fig. 3D). Esta variable (presencia de ganado) resultó significativamente relacionada con el tipo de ambiente (r = 0,743, p < 0,001), coincidiendo la presencia de ganado con el tipo de humedal, mallín y ausencia en costas de lagos profundos.
Relación con la riqueza
Se determinó un total de 101 especies, lo cual representa casi al 10% de la flora del PNNH, 60 de las cuales fueron nativas y 39 introducidas (4 especies quedaron sin identificar) (Tabla 2). El sitio con menor número de especies fue Laguna Fantasma (19 taxa) mientras que el mayor número de especies fue hallado en el mallín Frizón (31 taxa). La única especie cuya presencia se confirmó en todos los mallines fue P anserina. Otras especies halladas en más de la mitad de los sitios fueron las nativas Eleocharis pachycarpaÉ.Desv. (7 sitios), Carex aematorhyncha É.Desv. (6 sitios), Schoenoplectus californicus (C.A.Mey.) Soják (6 sitios) y la introducida Taraxacum officinale (diente de león) F.H.Wigg (5 sitios). Hubo 62 especies que fueron halladas en
El porcentaje de especies introducidas fue mayor en el mallín Teleférico (80%) mientras que el menor porcentaje se encontró en el mallín Llao-Llao (19%) (Tabla 2). La cobertura de P anserina fue menor al 10% en todos los sitios, con la excepción de Laguna Fantasma, donde la cobertura alcanzó el 47%, y del mallín del lago Morenito, con un 10% (Tabla 2). No se halló una relación entre la cobertura de P. anserina y el número total de especies de cada mallín, ni con el número de especies nativas ni con el número ni el porcentaje de introducidas (Tabla 3).
Discusión y conclusión
La distribución de P. anserina registrada sugiere que la especie se halla ampliamente distribuida en humedales temporarios (mallines) y en la zona litoral de lagos someros. De las variables categóricas analizadas, el tipo de humedal y la presencia de ganado fueron las más relacionadas con la distribución de esta especie. La presencia de ganado ha sido relacionada por otros autores, en particular para la provincia de Chubut, con una mayor cantidad y cobertura de especies introducidas (Kutschker et al., 2014). Sin embargo, hay que tener en cuenta que en nuestro estudio el tipo de humedal y la presencia de ganado fueron variables altamente correlacionadas.
Y por otro lado, en los dos mallines que no hubo ganado (mallín Llao-Llao y Bernal), P anserina igualmente estuvo presente. En particular en el mallín Llao-Llao, que se encuentra inmerso en el bosque del Parque Municipal Llao-Llao, su presencia coincide con la entrada al mallín, lo cual sugiere que el ingreso de personas al mallín podría haber actuado como un vector de dispersión. A su vez, el mallín Bernal, junto al Aeropuerto Internacional de Bariloche es un área altamente impactada por actividades recreativas (como motocrós).
De este modo, el tipo de hábitat no parece influir en la distribución de esta especie, ya que fue encontrada a lo largo de toda la transecta, tanto en el bosque como en la estepa. Incluso se registró su presencia en un mallín de alta montaña a 1504 m s.n.m. (mallín Los Patos). Su distribución tampoco estuvo asociada al grado de urbanización, lo cual indicaría que tiene un rango amplio de tolerancia a diferentes condiciones ambientales y disturbios antrópicos. La amplia tolerancia de esta especie a factores ambientales se puede concluir también a partir de haberse registrado tanto en zonas costeras del Pacífico (Hauenstein et al., 2008), en dunas de la provincia de Buenos Aires (Faggi & Dadon, 2010), como en mallines de Torre del Paine (Clausen et al., 2006).
A pesar de la falta de registros históricos que indiquen la distribución de P. anserina en función del tiempo en el PNNH, la amplia distribución hallada indicaría que esta especie es capaz de reproducirse en áreas distantes a >100 m de las plantas parentales en menos de 50 años, característica que demostraría que se trata de una especie invasora sensu Richardson et al. (2000). Sin embrago, no se halló una relación entre la cobertura de P. anserina y la riqueza total de especies, lo cual coincide con lo hallado por Stohlgren et al. (2003), que no encuentra una relación significativa entre las plantas nativas y las introducidas. En este sentido, Rafaele (2004) halló que en un mallín de altura sin disturbio antrópico, las especies introducidas fallaron en su establecimiento. Esto coincide con la mayor presencia de P. anserina en sitios con impacto antrópico (ganado). Sin embargo, hay que destacar que el único sitio con una cobertura de esta especie de casi el 50% (laguna Fantasma) fue también el que presentó la menor riqueza de especies. Este ultimo resultado indicaría que la cobertura podría estar negativamente asociada a la riqueza de especies (Fig. 4), pero se requiere más estudio para poder afirmar esto.
En conclusión, en este trabajo se determinó que P. anserina tiene una distribución amplia en el PNNH, que abarca desde humedales de bosque, de altura, y de estepa, tanto rurales como urbanos, y que si bien es característica de mallines (en particular aquellos con presencia de ganado), también puede estar presente en la zona litoral de lagos someros y profundos con impacto antrópico. Si bien no se demostró que su cobertura afecte a la riqueza de especies totales ni nativas, ni que facilite la instalación de nuevas especies introducidas, es necesario establecer cuáles son los factores que facilitan su expansión para intentar evitar que se exprese su potencialidad como invasora. Sobre todo teniendo en cuenta que trabajos previos indicaron que sí es una planta transformadora, ya que se demostró que introduce cambios en el ecosistema, como una alta liberación de materia orgánica disuelta coloreada (Cuassolo et al., 2011) y tasas de descomposición mayores a las de las especies nativas que coexisten con ella (Cuassolo et al., 2020). En cuanto a las tramas tróficas, se ha demostrado que es utilizada como alimento por larvas de insectos detritívoras (Diaz Villanueva & Trochine 2005), pero aún falta comparar cómo afecta a la comunidad de consumidores el reemplazo de una dieta de especies nativas a una dieta basada en esta especie introducida.
Contribución de las autoras
Ambas autoras han realizado conjuntamente y en partes iguales la colecta de datos, su interpretación y la redacción del manuscrito. F.C, realizó la identificación de las especies.
Agradecimientos
Agradecemos a las instituciones que han hecho posible este estudio. Agencia Nacional de Promoción Científica y Técnica, Proyectos PICT 2018- 4385. FC y VDV son investigadoras de CONICET. También, un especial agradecimiento a la Dra. Cecilia Ezcurra por su invaluable ayuda en la determinación de las especies. Por último, agradecemos a la Estancia Fortín Chacabuco por permitir realizar los muestreos en el mallín Frizón.

