











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
Los afloramientos triásicos de la República Argentina albergan numerosos registros pa leoxilológicos. En la provincia de San Juan me recen mencionarse las formaciones Ischigualasto (Archangelsky & Brett, 1963; Bonetti, 1966; Bodnar & Artabe, 2007), Cortaderita (Menéndez, 1956; Lutz & Herbst, 1992; Bodnar et al., 2015), Carrizal (Herbst, 1994; Lutz, 2006), Barreal (Bodnar, 2008, 2019), Hilario (Vallejos Leiz et al., 2019) y Los Rastros (Crisafulli & Lutz, 2008). En la provincia de Río Negro, fueron examinadas las maderas de la Formación Paso Flores (Zamuner & Artabe, 1994), mientras que en la provincia de Santa Cruz fue estudiada la lignoflora de las formaciones Laguna Colorada y Cañadón Largo que conforman el Grupo El Tranquilo (Crisafulli & Herbst, 2011). En Mendoza, se analizaron las paleoxilofloras de la Formación Paramillo (Brea, 1997; Artabe et al., 1999; Artabe & Brea, 2003).
Menéndez (1951) inició la descripción de maderas de los niveles de la Formación Llantenes. Crisafulli & Herbst (2010) y Gnaedinger & Zavattieri (2015, 2020) describieron leños de gimnospermas de Ginkgoales y Pinales colecta dos en diversas campañas a la localidad de Cerro Chihuido. Por ello, el propósito de este trabajo es continuar con la observación y determinación de los mismos para enriquecer la composición xilotafoflorística de esta formación, analizar los aspectos paleoclimáticos, paleoecológicos y esta blecer comparaciones con otras paleoxilofloras coetáneas.
Marco geológico
Las maderas proceden de la Formación Llantenes (Triásico Superior) que aflora en el Cerro Chihuido, considerado como la localidad tipo; ubicada a los 35° 38´S y 69° 35´O en las cer canías del arroyo Llantenes a 20 km de Malargüe al SO de la provincia de Mendoza (Fig. 1).

La cuenca de Malargüe está representada por el Grupo Tronquimalal (Stipanicic, 1979) conformado por las formaciones Chihuido y Llantenes, unidad inferior y unidad superior, respectivamente (Stipanicic, 1949). Este Grupo se apoya en discordancia erosiva y angular so bre los depósitos volcaniclásticos del Grupo Choiyoi (Groeber, 1929, 1946) y es cubierto en discordancia erosiva por las sedimentitas de la Formación Remoredo (Stipanicic, 1966).
La Formación Chihuido está compuesta pre dominantemente por facies clásticas gruesas y su modelo de depositación ha sido interpretada por Artabe et al. (1998) como depósitos de abani co aluvial, planicie entrelazada y eventualmente delta entrelazado desarrollados en una región húmeda con influencia volcánica. Estos depósi tos constituyen el primer estadio de relleno de la cuenca (sinrift inicial), el cual se caracterizó por un fuerte control de la subsidencia tectónica (Spalletti, 1997).
La Formación Llantenes, portadora de los restos leñosos, está integrada por una sucesión de 180 m de espesor (Menéndez, 1951) en la que predominan pelitas gris oscuras a negras a las que se asocian fangolitas, areniscas muy finas a finas y algunos conjuntos de areniscas me dianas a gruesas y conglomerados finos. En base a los atributos de las facies, Artabe et al. (1998) establecen el modelo deposicional de esta forma ción como un lago de tipo meromíctico, relativa mente profundo, hacia el que progradan sistemas deltaicos con dominio fluvial y desembocaduras de condición hipopicnal. Esta unidad represen ta la segunda etapa de relleno de la cuenca (sinrift tardío a postrift inicial), caracterizada por una disminución de la subsidencia tectónica y una posible incidencia de la subsidencia termal (Spalletti, 1997).
MATERIALES Y MÉTODOS
Los materiales estudiados corresponden a 16 fragmentos de leños fósiles permineralizados y silicificados con buena preservación de los teji dos, colectados de niveles de la sección media a superior de la Formación Llantenes. Los ejem plares están decorticados y con leño picnoxílico preservado. En algunos ejemplares se conserva la médula. Los materiales pertenecen a la Colección Paleobotánica del Museo de Ciencias Naturales y Antropológicas “Juan Cornelio Moyano” de la ciudad de Mendoza bajo el acrónimo MCNAM-PB con los números 1515-1519 y 1534-1544.
Para el procesamiento de las maderas se realizaron cortes petrográficos en las secciones transversal, longitudinal radial y longitudinal tangencial siguiendo el método de Hass y Rowe (1999) y luego pulidas hasta lograr el punto ópti mo de delgadez (aproximadamente 40 μm).
Las secciones planas y las láminas delgadas de los leños fósiles fueron observadas y fotogra fiadas con microscopio óptico Leica CDM 500 y lupa estereoscópica Leitz en el Laboratorio de Xilología (CECOAL-UNNE-CONICET).
Se obtuvieron un mínimo de 30 mediciones de cada caracter, registrándose en las descripcio nes primero la media y entre paréntesis los valo res mínimos y máximos.
La terminología utilizada corresponde a la lista de caracteres microscópicos para la iden tificación de maderas blandas de IAWA (2004), Philippe & Bamford (2008) y los elementos ana tómicos se midieron con los valores biométricos establecidos por García Esteban et al. (2002, 2003).
En las descripciones se siguió el estilo de re dacción usado por Pujana et al. (2017).
Para la clasificación supragenérica se siguie ron las normas de Cole et al., 2017.
RESULTADOS
Paleontología Sistemática
Euphyllophyta Kenrick & Crane, 1997
Spermatophyta Willkomm, 1854
Pinales (Dumortier) Gorozhankin, 190483
ChapmanoxylonPant & Singh, 1987
Especie tipo. Chapmanoxylon raniganjensisPant & Singh, 1987
Chapmanoxylon jamuriense (Maheshwari) Pant & Singh, 1987

Fig. 2 A-F. Chapmanoxylon jamuriense Pant & Singh. A. CT del xilema secundario (MCNAM-PB 1518 a), detalle de las traqueidas. B. CLRd (MCNAM-PB 1518 b) Detalle de las punteaduras uniseriadas circulares en las paredes traqueidales. C-F. CLTg. (MCNAM-PB 1518 c) C. Aspecto general de una traza con médula (M) en el centro rodeada de xilema (X). D. Distorsión en el trayecto de los radios (R) en la cercanía de la traza (Tr). E. Distribución y frecuencia de los radios uniseriados. F. Detalle de los radios uniseriados y biseriados (flechas). G. Megaporoxylon kaokense Kräusel. CT del leño (MCNAM-PB 1517 a). Detalle de la médula heterogénea con células esclerenquimáticas (flechas). Escala: A, D, E, G: 200 μm; B: 150 μm; F: 100 μm; C: 1,4 mm.

Fig. 3 CLRa A. Chapmanoxylon jamuriense Pant & Singh. Esquema de detalle de los campos de cruzamiento. B. Agathoxylon sp. Esquema de las punteaduras en las traqueidas, campos de cruzamiento, placas de resina e indentures (ver Figs. 6 A, B).
1966. Dadoxylon jamuriense Maheshwari, The Palaeo botanist, 13:148-152, lám.1, figs. 1-5, lám. 2, fig. 6.
1972. Damudoxylon jamuriense (Maheshwari) Ma heshwari, Palaeontographica B, 138:11.
1987. Chapmanoxylon jamuriense (Maheshwari) Pant & Singh, Palaeontographica B, 203: 4-21.
2008. Chapmanoxylon jamuriense (Maheshwari) Pant & Singh, 1987 en Crisafulli & Herbst, Ameghinia na, 45 :739-740, figs. 3. 1-4 y 4.1-2.
2009. Chapmanoxylon jamuriense (Maheshwari) Pant & Singh, 1987 en Crisafulli, Herbst & Manza Stort ti, Gaea, Journal of Geoscience, 5: 7-8, figs. 5. D-G y 6. A-D.
2012. Chapmanoxylon jamuriense (Maheshwari) Pant & Singh, 1987 en Crisafulli, Herbst & Dutra, Resu men XV Simposio Argentino de Paleobotánica, 36.
2012. Chapmanoxylon jamuriense (Maheshwari) Pant & Singh, 1987 en Leiva Verón, Crisafulli, Herbst, Filippi & Molina, Gaea, Journal of Geoscience, 8: 71, figs 3. C-D. Tabla 2.
2016. Chapmanoxylon jamuriense (Maheshwari) Pant & Singh, 1987 en Crisafulli, Herbst & Dutra, Gaea, Journal of Geoscience, 9: 42-44, figs. 5. E-F y 6. C.
Material estudiado. MCNAM-PB 1518 y pre parados microscópicos a, b y c.
Descripción. Fragmento de leño silicificado, de corticado, de color castaño claro, de 8 cm de lon gitud, 4 cm de diámetro menor por 7,5 cm de diá metro mayor. Externamente con marcas de cica trices foliares de forma romboidal dispuestas en forma espiralada. Xilema secundario picnoxílico, anillos de crecimiento indistintos. Traqueidas rectangulares, cuadrangulares a hexagonales en sección transversal con paredes gruesas; diáme tro radial y tangencial de 57 y 63 μm, respecti vamente. Radios con trayectos discontinuos y separados entre sí por 7 (4-11) traqueidas (Fig. 2. A). Paredes radiales de las traqueidas con punteaduras araucarioides uniseriadas, escasas biseriadas, circulares y contiguas que miden 7,5 μm de diámetro. Traqueidas con extremos ro mos (Fig. 2. B). Campos de cruzamiento con 2-5 punteaduras circulares, areoladas, generalmente contiguas en dos hileras, a veces aplastadas (Fig. 3. A). En sección longitudinal tangencial, pun teaduras araucarioides uniseriadas y contiguas. Traza rameal con médula homogénea, compacta, no tabicada y de contorno irregular aguzada en los extremos (Fig. 2. C). Células parenquimáticas grandes, redondeadas o globosas; diámetros ra dial y tangencial de 90 y 70 μm, respectivamente. Xilema primario obliterado. Presentes en este corte otras trazas rameales-foliares de contor no ovoidal con un diámetro promedio de 4 mm de ancho y 6 mm de alto y con un característico patrón en espiral (Bodnar & Coturel, 2012) ro deadas por traqueidas y radios leñosos curvos formando un remolino; los radios que rodean la parte superior de la traza son muy bajos, de 1 a 4 células de alto (Fig. 2. D). Los restantes ra dios leñosos muy bajos y medianos (1-14 células), registrándose con mayor frecuencia los de 4 cé lulas de alto. Homocelulares, uniseriados y uni seriados parcialmente biseriados. Células centra les redondeadas y en los extremos elipsoidales. Densidad de los radios 28 r/mm2 y frecuencia de 6 r/mm (Fig. 2. E-F).
Comentarios. La presencia de una médula ho mogénea en la traza concuerda con el género ChapmanoxylonPant & Singh (1987) por poseer este mismo caracter en la médula del eje prin cipal (Mussa, 1982; Crisafulli & Herbst, 2008). Este patrón se repite en otros géneros como en Kaokoxylon Kraüsel (1956) con células escleren quimáticas en la médula del tronco y en la traza rameal. Pant y Singh (1987) crearon este taxón para reunir a los leños con médula homogénea asignados a DadoxylonEndlicher (1847) que, des de 1978, de acuerdo con el Código Internacional de Nomenclatura Botánica se considera inválido. La problemática sobre la complejidad de la apli cación del nombre Dadoxylon está exhaustiva mente explicada en Mussa (1982).
Crisafulli & Herbst (2008) brindaron una de tallada información sobre las características de Chapmanoxylon; principalmente por el carácter peculiar de la médula homogénea siendo un ta xón del Neopaleozoico y Triásico del Gondwana en los que predominan las médulas heterogé neas. En ese trabajo se mencionan otros taxo nes fósiles con este carácter: BageopitysDohms (1976); AbietopitysKräusel (1928) en Kraüsel & Range (1928); ArauspiropitysPant & Sing (1987) y GinkgophytoxylonTidwell & Munzing (1995), realizándose una minuciosa comparación.
Consecuentemente, este ejemplar puede ser albergado en ese taxón fósil que está distribui do en Gondwana desde el Pérmico al Triásico Superior con numerosas especies (Crisafulli & Herbst, 2008; Crisafulli et al., 2018).
En la sección longitudinal radial presenta punteaduras uniseriadas, circulares contiguas tí picamente araucarioides. Las punteaduras en los campos de cruzamiento de una a cinco permiten acercarnos a Chapmanoxylon jamuriense. Este taxón es citado para el Pérmico de las formacio nes Raniganj de la India (Pant & Singh, 1987), Solca de Argentina (Crisafulli & Herbst, 2008), Tacuary de Paraguay (Leiva Verón et al., 2012) y Tres Islas de Uruguay (Crisafulli et al., 2009), y para el Triásico de la Formación Caturrita, Brasil (Crisafulli et al., 2016, 2018). La mayor diferencia es el número de punteaduras en los campos de cruzamiento que varía de 1 a 5 en el ejemplar analizado y de 1 a 3 en el holotipo. Otra distinción es la altura de los radios, compuestos por 45 células parenquimáticas, mientras que en el material de Llantenes varía de 1 a 14 células asemejándose más al ejemplar de la Formación Caturrita de Brasil, aunque el promedio es simi lar para todas las formaciones mencionadas. La presencia de radios uniseriados parcialmente bi seriados también se asemeja al ejemplar hallado en la Formación Caturrita (Tabla 1).
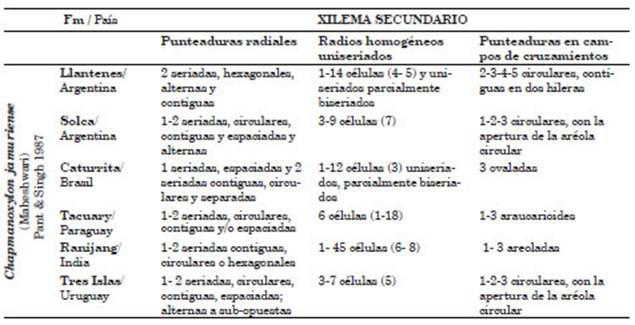
Tabla 1 Variaciones en las características anatómicas de Chapmanoxylon jamuriense (Maheshwari) Pant & Singh.
Megaporoxylon (Kräusel, 1956) emend. Maheshwari, 1972
Especie tipo. Megaporoxylon kaokenseKräusel, 1956
Megaporoxylon kaokenseKräusel, 1956
Figs. 2 (G), 4 (A-F) y 5 (A-C)

Fig. 4 A-F. Megaporoxylon kaokense Kräusel. A-D. CT del leño (MCNAM-PB 1517 a). A-B. Aspecto general de la médula (M), xilema primario (X1) y secundario (X2), en B traza migrando (Tr). C. Detalle de la traza en el xilema. D. Detalle del xilema secundario con anillos de crecimiento. E. CLRd. (MCNAM-PB 1517b). Aspecto de las traqueidas con punteaduras uniseriadas contiguas y espaciadas y campos de cruzamiento en el xilema secundario. F. Detalle de las punteaduras en los campos de cruzamiento. Escala: A: 0,7 cm; B, D, E: 200 μm; C: 120 μm; F: 50 μm.

Fig. 5 A-C. Megaporoxylon kaokense Kräusel. A. CLRd. (MCNAM-PB 1517b). Detalle de las punteaduras uniseriadas espaciadas en las traqueidas del xilema secundario. B-C. CLTg. (MCNAM- PB 1517 c). B. Aspecto de la traza (Tr) rodeada por los radios (R). C. Frecuencia y distribución de los radios leñosos uniseriados. D. Agathoxylon sp. CT del leño. (MCNAM-PB 1536 a). Detalle de las traqueidas cuadrangulares y de los radios rectilíneos distorsionados por alteración tafonómica. Escala: A: 80 μm; B: 0, 2 mm; C- D: 100 μm.
1956. Megaporoxylon kaokense Kräusel en Senckenber giana Lethaea, 37:421, pl. 6, figs. 22-24.
1961. Megaporoxylon kaokenseKräusel, 1956 en Kräu sel, Maithy & Maheshwari, The Paleobotanist, 10: 102, figs. 21a, 21d.
1972. Megaporoxylon kaokenseKräusel, 1956 en Ma heshwari, Paleontographica B, 138:12.
2016. Megaporoxylon kaokenseKräusel, 1956 en Cri safulli, Herbst & Dutra, Gaea, Journal de Geocien ce, 9: 40- 42. Figs. 4. F; 5. A- C y 6. A.
Material estudiado. MCNAM- PB 1517 y pre parados microscópicos a, b y c.
Descripción. Fragmento de leño silicificado, decorticado, de color castaño claro, de 7,4 cm de longitud, de 3,8 cm diámetro menor y 5,4 cm de diámetro mayor; médula preservada con proyec ciones cuneiformes del xilema primario y xilema secundario con anillos de crecimiento marcados (Figs. 2. G y 4. A-B). Médula heterogénea, com pacta, no tabicada y de contorno irregular en corte transversal. Células parenquimáticas poli gonales. Células esclerenquimáticas de contorno pentagonal, dispersas en la médula (Fig. 2. G). Xilema primario endarco, no bien preservado, dispuesto en fascículos cuneiformes rodeando a la médula (Fig. 4. A). Xilema secundario picnoxí lico con anillos de crecimiento verdaderos y falsos (Fig. 4. B). Trazas rameales de contorno aguzado en los extremos con promedio de 0,9 mm x 1,3 mm (Fig. 4. C). Traqueidas rectangulares, cua drangulares a hexagonales, con paredes gruesas y diámetro radial y tangencial de 24 (12-32) μm y 23 (18-28) μm, respectivamente. Un promedio de 7 y un rango de 4-11 traqueidas separan los radios entre sí. Leño tardío con una media de 2 células de ancho. Transición del leño temprano al tardío abrupta (Fig. 4. D). Paredes radiales de las traqueidas con punteaduras araucariodes, unise riada, contiguas y espaciadas, raramente biseria das. Campos de cruzamiento con una punteadura simple, oval en su mayoría y otros con dos o tres más pequeñas y en dos hileras (Fig. 4. E-F y 5. A). En CLTg, presenta algunas trazas rodeadas por radios uniseriados y unos pocos uniseriados parcialmente biseriadas (Fig. 5. B). Altura de 6 (4-11) células. Frecuencia de los radios 10 r/mm (Fig. 5. C).
Comentarios. La presencia de una médu la heterogénea con células esclerenquimáticas asemeja este ejemplar a géneros fósiles como KaokoxylonKräusel (1956); ScleromedulloxylonMussa (1982) y Megaporoxylon Kräusel (1956). Sin embargo, la distribución dispersa de dichas células en la médula lo aparta de Kaokoxylon. De Scleromedulloxylon, se diferencia porque las células esclerenquimáticas del material de la Formación Llantenes no se encuentran en tabi ques o diafragmas medulares.
En cuanto a los rasgos del xilema secun dario puede compararse con MegaporoxylonKräusel (1956); Phyllocladopitys Kräusel en Kräusel & Range (1928); Circoporoxylon Kraüsel (1949) y PhyllocladoxylonGothan (1904), prin cipalmente por similitudes en las punteaduras en los campos de cruzamiento caracterizados por Lepekhina & Yatsenko (1966) como tipo de Protophyllocladoxylon. Sin embargo, solo Megaporoxylon y Phyllocladopitys conservan la médula, pero se diferencian en que Megaporoxylon tiene una médula heterogénea como el material estudiado y Phyllocladopitys una médula hete rogénea /homogénea y xilema primario mesarco. No obstante, este rasgo y la combinación de las punteaduras en las paredes de las traqueidas jus tifican la asignación a Megaporoxylon, que posee punteaduras araucarioides uniseriadas, rara mente biseriadas y campos de cruzamiento con punteaduras simples, ovales y en dos hileras, que son los rasgos de la especie M. kaokense Kräusel (1956). Este taxón ha sido hallado en los sedi mentos del Permo-Triásico de Namibia, África (Kräusel, 1956), en la Formación Alan Nunatak, Antártica (Maheshwari, 1972) y en el Triásico de la Formación Caturrita, Brasil (Crisafulli et al. 2016, 2018).
AgathoxylonHartig, 1848sensu Philippe, 1995
Especie tipo. Agathoxylon cordaianumHartig, 1848
Agathoxylon sp.

Fig. 6 A-D. Agathoxylon sp. A-C. CLRd. (MCNAM-PB 1536 b). A. Detalle de punteaduras uniseriadas. B. Punteaduras uniseriadas en trayectoria oblícua, en grupo y campos de cruzamiento con punteaduras areoladas (las dos flechas de la derecha); placas de resina en las paredes traqueidales (Pl. r) e “indentures” (las dos flechas de la izquierda). C. Punteaduras uniseriadas contiguas (flechas a la izquierda) y engrosamientos helicoidales (flechas a la derecha). D. CLTg. (MCNAM-PB 1536 c). Frecuencia y distribución de los radios uniseriados y uniseriados parcialmente biseriados. E. Agathoxylon protoaraucana (Brea) Gnaedinger & Herbst. CT. (MCNAM-PB 1535 a) Shearing zones (Sh. z) en el leño secundario. Escala: A: 100 μm; B: 50 μm; C: 100 μm; D: 120 μm; E: 250 μm.
Material estudiado. MCNAM- PB 1534 y 1536 y preparados microscópicos: a, b y c.
Descripción. Fragmento de leño silicificado, decorticado. picnoxílico con anillos de crecimien to no marcados. En CT, shearing zones sensuErasmus (1976). Traqueidas cuadrangulares con paredes muy gruesas. Diámetro radial de 60 (50-70) μm y diámetro tangencial de 60 (53-74) μm. Radios parenquimáticos continuos y separa dos por 2 (1-5) traqueidas entre sí (Fig. 5. D). Forma y seriación de las punteaduras radiales en las traqueidas: circulares, pocas hexagonales, típicamente araucarioides, uniseriadas y unise riadas parcialmente biseriadas con aréolas cir culares (Fig. 6. A). Variedad en la disposición de las punteaduras: contiguas, en grupo de manera oblicua y separadas unas de otras; contiguas y espaciadas en la misma traqueida. Algunas tra queidas con extremos en bisel, otras aguzados y también romos. Placas bicóncavas de resina. Campos de cruzamiento con 2-4 punteaduras, apretadas unas con otras en dos columnas, algu nas con aréola preservada, hendiduras radiales (indentures) en los campos de cruzamientos, sen suGarcía Esteban et al. (2003), ubicadas sobre las paredes horizontales en las proximidades de las paredes axilares (Figs. 3. B; 6. B). Finos y es casos engrosamientos helicoidales en la pared de la traqueida (Fig. 6. C). En CLTg, radios homo celulares, uniseriados parcialmente biseriados. Densidad 36 r/mm2. Bajos a medianos, con un rango de 5-21 células de altura y un promedio de 11 células (Fig. 7). Células del centro de contorno rectangular y las de los extremos triangular. En las centrales, punteaduras circulares en número de 2 a 4. Punteaduras uniseriadas dispuestas en forma contigua en las traqueidas (Fig. 6. D).

Comentarios. La diversidad de punteadu ras en las paredes radiales de las traqueidas en CLRd principalmente, acerca este ejemplar a AgathoxylonHartig (1848), AustraloxylonMarguerier (1973) y TaeniopitysKräusel (1956). Fundamentalmente, la presencia de punteadu ras en grupos en las paredes de las traqueidas cortas lo aproxima a Australoxylon. Sin embar go, no se han observado punteaduras simples en los campos de cruzamiento como las que posee este taxón, al cual no se lo ha encontrado en sedimentos más jóvenes que los del Pérmico en el Gondwana. Puede compararse asimismo con Taeniopitys, hallado en los sedimentos pérmicos de las montañas transantárticas, con una única especie T. scotti Kräusel (1956), pues tiene mar cadas semejanzas en cuanto al juego de puntea duras en sección longitudinal radial y comparte la presencia de punteaduras en grupos. No obs tante, en el material en estudio están ausentes las punteaduras triseriadas, hexagonales, la mé dula y el xilema primario, por lo que sería im prudente albergarlo en este género ya que no es posible establecer comparaciones.
La presencia de indentures o hendiduras ra diales es un rasgo de alto valor diagnóstico, que caracteriza a Planoxylon Stopes; ya que se ob serva en varias de sus especies: P. gnaedingeriae Crisafulli del Pérmico Superior de la Formación Yaguarí y P. australe (Salard) Vozenin Serra & Salard Cheboldae hallado por Gnaedinger (2007) en la Formación La Matilde. Sin embar go, Planoxylon posee parénquima axial y un plan leñoso mixto que lo apartan del ejemplar aquí descripto (Stopes, 1916; Crisafulli, 2003; Gnaedinger, 2007).
La combinación del plan araucarioide de este especimen y la presencia de indentures o hendiduras no compatibles con otra especie de Agathoxylon (Crisafulli & Herbst, 2011; Cabral Kloster & Gnaedinger, 2018; Vera et al., 2019) no permite asignarlo a una especie conocida, razón por la cual se lo asigna a Agathoxylon sp.
Agathoxylon protoaraucana (Brea) Gnaedinger & Herbst, 2009
Figs. 6. (E), 8 (A-G) y 9

Fig. 8 A-G. Agathoxylon protoaraucana (Brea) Gnaedinger & Herbst. CT. del leño (MCNAM-PB 1535 a). A. Detalle de las traqueidas y radios. B-E. CLRd. (MCNAM-PB 1535 b). B-D. Detalle de las punteaduras araucarioides areoladas, uniseriadas en las paredes traqueidales. E. A la izquierda de la figura, punteaduras uniseriadas y a la derecha punteaduras biseriadas dispuestas en los márgenes de las traqueidas (flecha). F-G. CLTg. del xilema secundario (MCNAM-PB 1535 c). F. Frecuencia y distribución de los radios uniseriados y punteaduras circulares areoladas uniseriadas en la pared de la traqueida (flecha). G. Radio uniseriado parcialmente biseriado (flecha). Escala: A: 160 μm; B, F, G: 120 μm; C: 80 μm, D: 45 μm; E: 30 μm.

Fig. 9 Agathoxylon protoaraucana (Brea) Gnaedinger & Herbst. CLRa. Esquema de los campos de cruzamiento.
1997. Araucarioxylon protoaraucana Brea, Ameghinia na 34: 486-488, Lámina 1-3; fig.3.
2009. Agathoxylon protoaraucana (Brea) Gnaedinger & Herbst, Ameghiniana 46: 61-63, figs. 3. 1- 2, 4-5. Cuadro 2.
Descripción. Leño picnoxílico silicificado, sin límite de anillos de crecimiento y shearing zo nes (Erasmus, 1976). Contorno cuadrangular a rectangular de las traqueidas del leño temprano en corte transversal, con un diámetro medio ra dial y tangencial de 45 (39-59) μm y 38 (23-52) μm, respectivamente. Transición del leño tem prano al tardío, gradual. Radios separados entre sí por unas 4 (1-7) traqueidas. Radios disconti nuos sin trayecto rectilíneo (Figs. 6. E y 8. A). Punteaduras radiales de 17 (14-20) μm x 14 (13- 15) μm de diámetro, con la apertura de la aréola circular o elíptica. Coeficiente de achatamiento: e = d (diámetro menor) / D (diámetro mayor) 0,65 (13/20). Uniseriadas, circulares, contiguas, de tipo araucarioide ó espaciadas con tendencia abietinoide, biseriadas, circulares, ocasionalmen te opuestas (Fig. 8. B-D). Campos de cruzamien to con punteaduras circulares, contiguas de 4 a 6 y dispuestas en dos hileras (Fig. 9). En CLTg, radios leñosos homocelulares, uniseriados y uni seriados parcialmente biseriados los bajos (3-20 células), registrándose con mayor frecuencia los radios de 11 células de altura. Células centrales de los radios ovoides y elípticas las de los extre mos. Punteaduras areoladas uniseriadas con apertura circular en las paredes de las traqueidas (Fig. 8. F-G).
Comentarios. La presencia de xilema secunda rio picnoxílico, punteaduras radiales y campos de cruzamiento de tipo araucarioides permiten determinar este ejemplar como Agathoxylon. De entre las numerosas especies de este taxón, este material comparte similitudes con los ejem plares hallados por Brea (1997) en la Formación Paramillo del Triásico Medio y Superior de la lo calidad de Agua de la Zorra, Mendoza y con el de Gnaedinger & Herbst (2009) de la Formación Roca Blanca, perteneciente al Jurásico Inferior situada en la provincia de Santa Cruz. El tipo y la seriación de las punteaduras en las paredes radiales y tangenciales de las traqueidas y los campos de cruzamiento del material analizado muestran una mayor concordancia con el ejem plar descripto por Brea (1997), al presentar por campo de 3 a 9 punteaduras desordenadas o a ve ces alineadas en hileras verticales. Sin embargo, el ejemplar estudiado por Gnaedinger & Herbst (2009) presenta de 1 a 8 punteaduras dispuestas irregularmente, a veces alineadas de a dos o tres. Un rasgo sobresaliente del espécimen estudiado y no compartido, o al menos ausente en los ejem plares anteriormente mencionados, es la ubica ción de las punteaduras biseriadas opuestas en los márgenes de la pared radial de una traqueida, pero al no repetirse en otras células traqueales, no amerita diferenciarlo en otra especie. Cabe mencionar también que en este material el límite de anillos de crecimiento está ausente, posible mente porque no hubo una estacionalidad mar cada. Se lo ha comparado con A. lamaibandianusCrisafulli & Herbst (2011), pero la diferencia principal con esta especie se debe a la ausencia de punteaduras en las paredes tangenciales de las traqueidas (Tabla 2).

CONSIDERACIONES FINALES
Consideraciones paleobotánicas
En este trabajo se determinaron maderas gimnospérmicas con médulas preservadas de afinidad incierta: Chapmanoxylon jamuriense y Megaporoxylon kaokense además de otras que han conservado solo el xilema secundario cuyos caracteres se asemejan a los de las Araucariaceae: Agathoxylon protoaraucana y Agathoxylon sp. Todas ellas incluidas en el orden Pinales.
Estos taxones se suman a los que Crisafulli & Herbst (2010) describieron para la mis ma formación: una afín a las Araucariaceae, Agathoxylon dallonii (Boureau) Crisafulli & Herbst (2010); un representante afín a las Podocarpaceae, Podocarpoxylon tikiense Ram- Awata & Rajanikanth (2007) y otro con caracte rísticas anatómicas semejante a las formas más antiguas de las Taxaceae, Prototaxoxylon inter trappeumPrakash & Srivastava (1961) (Philippe et al., 2019); así como un leño perteneciente al Orden Ginkgoales, Baieroxylon chilensisTorres & Philippe (2002). Además, este acervo xilológico se vio incrementado por Gnaedinger & Zavattieri (2020) quienes estudiaron las xilofloras del Grupo Tronquimalal determinando Agathoxylon lamaibandianus Crisafulli & Herbst (2011) y dos especies correspondientes a las Cupressaceae: Cupressinoxylon llantenesense Gnaedinger & Zavattieri (2020) y Protojuniperoxylon ischigua lastense (Bonetti) Bodnar & Artabe (2007) para la Formación Llantenes. A. lamaibandianus se re pite en la infrayacente Formación Chihuido jun to a Agathoxylon cozzoi Gnaedinger & Zavattieri (2020) (Tabla 3).

Tabla 3 Distribución estratigráfica y geográfica de las especies de la Formación Llantenes en el Gondwana. Los taxones escritos en negrita corresponden a los hallados en este trabajo y los escritos en naranja a los encontrados en trabajos anteriores. Abreviaturas: Pm=Pérmico, Tr= Triásico, Jr= Jurásico.
Consideraciones bioestratigráficas
Esta asociación de maderas coincide en parte con varias formaciones coetáneas de Sudamérica, como las de La Coipa, La Ternera y las Breas en el norte de Chile (Lutz et al., 1999, 2001, 2003), la Formación Caturrita en el Sudeste de Brasil y la Formación Laguna Colorada en el sur de Argentina (Crisafulli & Herbst, 2011).
Agathoxylon protoaraucana fue registra do en las formaciones Paramillo en el Triásico Medio-Superior de Mendoza y Roca Blanca en el Jurásico Inferior de Santa Cruz. Gnaedinger & Herbst (2009) encontraron elementos arbó reos afines a las Araucariaceae, Cupressaceae, Taxaceae y Ginkgoales como en las asociaciones mencionadas más arriba.
Sin embargo, es éste el primer registro de Chapmanoxylon jamuriense y Megaporoxylon kaokense para el Triásico Superior de Argentina, estando presentes en la coetánea Formación Caturrita de Brasil constituyendo la importante flora de “bosques de altura” (upland forests) del que es parte asimismo la lignoflora del Norte de Chile (Lutz et al., 2001; Crisafulli & Herbst, 2008 y Crisafulli et al., 2016).
A su vez, los componentes de esta xilotafoflora son comunes en los bosques pérmicos de las for maciones Solca (La Rioja), Tres Islas (Uruguay) y Tacuary (Paraguay) donde están presentes Chapmanoxylon, Baieroxylon y Agathoxylon, lo que indica que son taxones fósiles muy frecuen tes.
Megaporoxylon kaokense fue hallado en el Pérmico de Namibia (África), en la Formación Alan Nunatak de Antártida (Maheshwari, 1972) y con su registro en la Formación Llantenes am plía su distribución geográfica.
Todas las especies aquí descriptas son exclu sivamente gondwánicas (Fig. 10). Sin embargo, cabe señalar que Podocarpoxylon, Prototaxoxylon y Agathoxylon son cosmopolitas y géneros de am plio biocrón pues se extendieron desde el Pérmico hasta el Neógeno.

Consideraciones paleoambientales
Los anillos de crecimiento aportan informa ción para caracterizar las condiciones climáticas por las que atravesaron estos leños. La iden tificación en estas maderas de anillos Tipo D y Tipo 0 de acuerdo a la clasificación de Creber & Chaloner (1984) en Brisson et al. (2001) in dican condiciones climáticas favorables para el crecimiento, con buena disponibilidad hídrica. Chapmanoxylon jamuriense, Agathoxylon sp. y Agathoxylon protoaraucana, poseen anillos Tipo 0 que reflejan estaciones de crecimiento relativa mente uniformes, sin variaciones marcadas como Agathoxylon dallonii y Podocarpoxylon tikiense, descriptos ya para esta xiloflora por Crisafulli & Herbst (2010).
Agathoxylon lamaibandianus, Cupressi noxylon llantenesense, Protojuniperoxylon ischigualastense (Gnaedinger & Zavattieri, 2020), Baieroxylon chilensis, Prototaxoxylon intertrapeum (Crisafulli & Herbst op. cit.) y Megaporoxylon kaokense desarrollaron anillos Tipo D, lo que significa que el límite entre el leño temprano y tardío es más marcado. Se observan en promedio 4 hileras de traqueidas correspon dientes al leño tardío.
Los resultados obtenidos en este trabajo rati fican lo expresado por Volkheimer & Papú (1993) al estudiar la composición de la microflora que refleja “condiciones de un clima por lo menos localmente húmedo” y que fuera enunciado por Crisafulli & Herbst (2010).
La composición xilotafoflorística de la Formación Llantenes, que señala condiciones de clima templado cálidas y húmedas, permite deducir que habría existido en la región estudia da una paleocomunidad que representaría una asociación higromesofítica a mesoxerofítica se mejante a las otras formaciones donde estaban citadas estas especies, en opinión de Agashe & Prasad (1989).
Consideraciones tafonómicas
A causa de los procesos de transporte y ente rramiento sufridos se visualizan atributos mor fo-anatómicos de importancia tafonómica en es tas maderas (Fernández López, 2000). Se obser va en corte transversal compresión en diferentes direcciones, ocasionando deformación del xilema secundario. Agathoxylon sp. y A. protoaraucana presentan shearing zones, que son sectores del leño temprano que sufrieron, al tiempo de la fosi lización, deformación o deslizamiento por acción de una fuerza cortante a consecuencia de que allí las traqueidas son débiles y menos resistentes (Erasmus, 1976). Estos elementos xilemáticos pueden ser fácilmente infiltrados por diversas sustancias como óxido de hierro, siguiendo un típico patrón oblicuo, por lo que son observables a simple vista como una banda tangencial de di ferente tonalidad.
Otro caracter tafonómico es la producción de S-shaped linear rows (Maheshwari, 1972). Es otra manera particular de disposición de los ele mentos xilemáticos en zig-zag (Fig. 11. A-F).

Fig. 11 A-D. CT. del leño. A-B. Shearing zones A. Agathoxylon sp. (MCNAM-PB 1536 a). B. Agathoxylon protoaraucana (Brea) Gnaedinger & Herbst. (MCNAM-PB 1535 a). C-D. Agathoxylon sp. (MCNAM-PB 1534 a), infiltraciones de óxido de hierro en las traqueidas de una madera indeterminada del mismo sitio fosilífero. E-F. S-shaped linear rows. E. Megaporoxylon kaokense Kräusel. (MCNAM- PB 1517 a) F. Agathoxylon sp. (MCNAM- PB 1534 a).
Consideraciones evolutivas
Desde el punto de vista evolutivo los rasgos anatómicos de las maderas estudiadas muestran un modelo araucariode-mixto-abietinoide, carac terístico de las maderas coetáneas, lo cual con tribuye a ratificar la edad Triásico Superior para esta formación. Estas consideraciones también se arriban a través de la flora de impresiones, y por los elementos de la microflora con especies tí picas como la presencia del polen de Classopollis que llega hasta el Daniano (Barreda et al., 2012).
CONCLUSIONES
El estudio de las maderas procedentes de la Formación Llantenes aflorante en la locali dad Cerro Chihuido del Grupo Tronquimalal (Mendoza) permitió arribar a la conclusión de que la xilotafoflora del Triásico de la Formación Llantenes fue más diversa que la conocida hasta el presente.
La identificación de los rasgos anatómicos diagnósticos condujo a la determinación sistemá tica de cuatro taxones fósiles de gimnospermas sensu lato.
Asimismo, los componentes de esta asociación de Araucariaceae, Podocarpaceae, Cupressaceae, Ginkgoales y maderas afines a las Taxaceae cuyos rasgos anatómicos muestran un modelo araucariode-mixto-abietinoide, han sido registra dos en yacimientos de India, Brasil, Argentina y Chile, ratificando por otra parte la edad Triásico superior de esta unidad estratigráfica.
Las condiciones templado cálidas y húmedas inferidas por la información sedimentológica y por la paleoflora permiten enunciar que en la Formación Llantenes se habría desarrollado una asociación higromesofítica a mesoxerofítica simi lar a los otros yacimentos donde se registaron las especies aquí preservadas.

