











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Los ecosistemas de agua dulce ocupan menos del 1% de la superficie de la tierra y albergan aproximadamente el 10% del total de las especies conocidas (Balian et al., 2008). El manejo sustentable de los recursos dulceacuícolas ha ganado importancia a escala regional y global debido al avance y deterioro progresivo de estos sistemas (Hart & Calhoun, 2010; Domínguez et al., 2020). Estos ecosistemas son importantes por los bienes y servicios de valor cuantificable que proporcionan al hombre: agua para consumo, alimento, forraje, medicinas, además de servicios indirectos como el control de inundaciones, filtración de agua, reducción de contaminación, recreación, entre otros (Russi et al., 2013). Lo preocupante es que esos beneficios y la biota asociada al recurso agua están expuestos a amenazas constantes, dado que la expansión antrópica y los cambios en el uso de la tierra, constituyen uno de los principales factores que afectan la biodiversidad a nivel mundial (Collen et al., 2014; Newbold et al., 2015). Estos servicios ecosistémicos sustancian en gran parte el desarrollo social sobre nuestro planeta y resulta clave la tarea de monitorear los cambios de los sistemas (Díaz et al., 2005). Argentina se destaca por su riqueza en sistemas hídricos los cuales albergan una biota muy diversa. Las condiciones climáticas permiten el establecimiento de diversos tipos de vegetación que contribuyen a los altos porcentajes de materia orgánica de los suelos que evitan su erosión y ayudan a incrementar la captación y almacenamiento de agua. Sin embargo, muchos de estos cursos de agua son afectados y sufren cambios debido al aporte de sustancias contaminantes. La cuenca Salí-Dulce es una de las principales del Noroeste de Argentina (NOA), en su recorrido atraviesa cinco provincias y recibe el aporte de diversos ríos y arroyos de orden inferior (Pérez Miranda et al., 2001; Adler, 2014). En Tucumán, el río Salí marca un corredor casi lineal en el cual se concentra el área de mayor desarrollo económico de la provincia y están establecidas varias industrias (ingenios, citrícolas, papelera, entre otras) por lo que también es uno de los sistemas más afectados por la actividad antrópica (Adler, 2014).
En los ambientes acuáticos las diatomeas ocupan un amplio rango de hábitats y la complejidad en la estructura y dinámica de los ensambles de estos organismos se debe a la interacción entre variables geomorfológicas, hidrológicas y bióticas que operan a gran escala e influyen en la dinámica trófica, además se relacionan estrechamente con los ciclos bioquímicos (Luque & Martínez de Fabricius, 2005; Martínez de Fabricius et al., 2010; Andrada-Servin & Israde-Alcántara, 2021). La caracterización de los diferentes ensambles de diatomeas a lo largo de un sistema fluvial permite definir distintos estados de sucesión de las mismas y a partir de ello dar una idea aproximada de la calidad de agua (Martínez de Fabricius et al., 2005). La biología y taxonomía de este grupo son propicias para respaldar su uso exitoso en el biomonitoreo basado en sus respuestas específicas a los cambios ambientales, especialmente la contaminación orgánica. Presentan un amplio espectro de tolerancia, desde condiciones oligotróficas hasta eutróficas, estas microalgas han sido ampliamente utilizadas como bioindicadores de la calidad del agua (Bellinger & Sigee, 2010; Wu et al., 2017; Ballesteros et al., 2020).
En tal sentido, es reconocida la importancia de los estudios bioecológicos de las algas, particularmente de diatomeas, a través de evaluaciones acerca de la diversidad, métricas de saprobiedad e índices basados en ensambles diatomológicos (Peña et al., 2005). Considerando el término “bioindicador” es importante resaltar que no hay bioindicadores específicos para cada tipo de contaminación, ni indicadores que puedan aplicarse de una manera estandarizada y universal, ya que los organismos que se encuentran en una región determinada tienen, por lo general, una distribución limitada y sus tolerancias pueden ser especie-específicas (Domínguez et al., 2022), por lo cual cobra relevancia la implementación y conocimiento de los ensambles de diatomeas en un área determinada.
El manejo y planificación de cuencas debería ir acompañado de una adecuada explotación del recurso agua. Las diversas actividades agrícolas, ganaderas e industriales son la causa por la cual muchos de los ecosistemas acuáticos se hallen en un estado de notable deterioro. Por lo que surge la necesidad de establecer el “status” ecológico actual de los sistemas acuáticos e implementar técnicas de biomonitoreo, resaltando que el uso de los indicadores biológicos y la medición de los parámetros fisicoquímicos son métodos complementarios y no alternativos en la evaluación de los ecosistemas acuáticos (Domínguez et al., 2022). El objetivo del presente trabajo fue caracterizar la dinámica y la composición de las taxocenosis de diatomeas y su relación con variables ambientales de los arroyos Mista y Calimayo, pertenecientes a la cuenca del río Salí-Dulce (Tucumán-Argentina).
Materiales y Métodos
Áreas de estudio
La cuenca del río Salí-Dulce (Fig. 1) pertenece al sistema hidrográfico Mar Chiquita, en cuyos márgenes se establecieron y viven aproximadamente 2,5 millones de personas. Este sistema hídrico abarca a las provincias de Salta, Catamarca, Tucumán, Santiago del Estero y Córdoba (Domínguez et al., 2020). En Tucumán el río Salí recorre gran parte de su territorio, el cual presenta un clima subtropical con régimen de lluvias en primavera-verano y durante el invierno ocurren las menores precipitaciones. Para este trabajo se seleccionaron dos tributarios del río Salí, el arroyo Mista ubicado en el departamento Leales (ecoregión Chaco seco, subregión Chaco Semiárido) y el arroyo Calimayo que se encuentra en la localidad de Lules (ecorregión de Yungas) (Morello et al., 2012). Estos arroyos reciben en diferentes sectores de su recorrido, efluentes industriales provenientes de ingenios, papeleras, frigoríficos y lixiviados agroquímicos de los campos cercanos, entre otros. Para determinar si existen variaciones significativas en los ensambles diatomológicos y en los parámetros fisicoquímicos se establecieron tres estaciones de muestreo en cada arroyo, los sitios 1 se consideraron como sitios no impactados o de referencia: Calimayo: S1C (26º55´55´´S - 65º23´17´´W); S2C (26º57´05´´S - 65º21´30´´W); S3C (27º23´51´´S - 65º19´54´´W), Mista: S1M (27º01´02´´S - 65º06´36´´W); S2M (27º11´16´´S - 65º06´08´´W) y S3M (27º21´37´´S - 65º04´01´´W).
Trabajo de campo y laboratorio
Se realizaron muestreos durante primavera, verano, otoño e invierno desde agosto de 2012 a mayo de 2014 en los tres sitios seleccionados en cada arroyo (Fig 1). Es necesario resaltar que no se dispone de datos fisicoquímicos o biológicos de la primavera del 2013 en el sitio 3 del arroyo Mista, dado que su cauce fue desviado aguas arriba del sitio de muestreo para riego de campos de cultivos, por un evento de sequía en esos meses.
Los parámetros físicoquímicos que se midieron in situ fueron: temperatura del agua, pH y conductividad eléctrica para lo que se utilizó un analizador multiparamétrico digital portátil para agua, Water Quality Meter 850081, Sper Scientific. En frascos tipo winkler se colecto agua para la determinación del oxígeno disuelto (OD) para lo cual se fijó con dos reactivos: 1- Sulfato de Manganeso (MnSO-4) y el reactivo 2- Ioduro de Potasio (KI) más Hidróxido de Sodio (NaOH) en el campo. Asimismo, se extrajeron muestras con recipientes plásticos de 1,5 litros, los que fueron trasladados al laboratorio en frío y oscuridad para su posterior tratamiento para el análisis de iones mayoritarios, demanda bioquímica de oxígeno (DBO5), compuestos nitrogenados y fosfato.
La metodología que se siguió para los análisis físicoquímicos fue la recomendada en APHA (2005). El control y aseguramiento de la calidad de los procedimientos de toma de muestras de agua y las determinaciones analíticas en laboratorio, se efectuaron de acuerdo a normas del Instituto Argentino de Normalización y Certificación (IRAM, 29012-2, 1996; 29012-3, 1998; 301/ISO17025, 2005). Para la evaluación del tipo de agua, se emplearon los diagramas de Piper-Hill-Langelier.
Las muestras cualitativas se obtuvieron con una red de plancton de 20 µm de malla expuesta a la corriente durante 20 minutos, fueron fijadas in situ con formaldehído al 4%. Las muestras fueron tratadas según la metodología de Batarbee (1986) y se realizaron preparados permanentes utilizando Naphrax como medio de montaje, para poder realizar una correcta determinación taxonómica de las especies de diatomeas. Para la identificación de las especies se utilizó un microscopio binocular marca Zeiss, los taxones fueron fotografiados con un software incorporado al microscopio y la bibliografía empleada fue mencionada en diversas publicaciones (Taboada et al., 2015, 2017, 2018, 2021).
Las muestras cuantitativas se extrajeron en envases de 250 ml y se fijaron in situ con formaldehído al 4%. La cuantificación se realizó bajo microscopio invertido siguiendo la metodología de Utermöhl (1958) se contó un número fijo de campos elegidos al azar, aceptándose un error de conteo de hasta 20%. Se contabilizaron 200 campos al azar hasta completar un total de 400 valvas (frústulo entero=1 unidad/individuo por ml).
Análisis de datos
Para jerarquizar la dominancia de las especies se construyó el diagrama de Olmstead & Tukey (Sokal & Rohlf, 1981). Las especies dominantes, constantes, ocasionales y raras (o pocos frecuentes) se determinaron a partir de la relación entre las densidades de los individuos y sus frecuencias de aparición.
Para analizar la diversidad de diatomeas se calculó el índice de Shannon & Weaver (1963) H (Magurran, 2004). Este índice y sus intervalos de confianza (IC) fueron calculados con el software Infostat (Di Rienzo et al., 2013). También se estimó la riqueza puntual con sus intervalos de confianza por el mismo método mencionado anteriormente.
Para la categorización del estado ecológico de los arroyos se aplicó el método de los saprobios (IS) de Pantle & Buck (Schwoerbel, 1975). El grado sapróbico se determinó a partir de la densidad y valencia sapróbica de cada especie con datos obtenidos de investigaciones propias, de bibliografía regional, de otros lugares y los registros de DBO5 (Slàdeček, 1973; Seeligmann et al., 2001; Licursi & Gómez, 2003; entre otros). Estos datos se incorporaron a la fórmula propuesta (IS) y se obtuvo el valor de saprobiedad.
Se utilizó el Índice de Diatomeas Pampeano (IDP) que fue diseñado para evaluar la eutrofización y polución orgánica de ríos y arroyos (Gómez & Licursi, 2001), este considera la sensibilidad de cada taxón con tres variables estrechamente relacionadas a diversos impactos eutróficos y orgánicos (demanda bioquímica de oxígeno (DBO5), amonio y fósforo reactivo soluble). El IDP varía entre 0 y 4, rango que define cinco tipos de clases de calidad de agua según las concentraciones de DBO5, NH4+ y PO43-, que son definidas como muy buena (0-0,5), buena (>0,5-1,5), aceptable (>1,5-2), mala (>2-3) y muy mala (> 3-4) (Licursi & Gómez, 2003). En este caso se reemplazó una de las variables, el P-PO4, por el ortofosfato.
Se utilizó el método multivariado análisis de correlación canónica (ACC) para relacionar una matriz de abundancia de especies de diatomeas con una matriz de variables ambientales. Se empleó esta técnica porque en un examen preliminar de los datos se observó que las longitudes de los gradientes ambientales fueron mayores a 2 (dos) condición sugerida para el uso del método ACC (Ter Braak & Šmilauer, 1998; Legendre & Legendre, 2000). Las variables que componían la matriz ambiental fueron: año (2012, 2013, 2014), arroyo (Mista, Calimayo), estación de colecta (S1, S2, S3), estación del año (seca, de mayo a octubre y húmeda de noviembre a abril), temperatura, pH, oxígeno disuelto, Na+, Ca2+, NH4+, PO43-, CE y K+. Se utilizaron test de permutación de Monte Carlo para comprobar la significancia de los ejes canónicos y las variables abióticas relacionadas a los mismos junto con correlación intergrupal.
Resultados
En el arroyo Calimayo el mínimo valor de temperatura fue de 13 ºC en S1C en otoño/13 y el máximo se presentó en S2C con 30 ºC (verano/14). Los registros térmicos del arroyo Mista oscilaron entre 12-26 ºC en los sitios S3M (invierno/12) y S1M (verano/14), respectivamente.
El pH fue siempre superior a 7 en ambos arroyos, en el Calimayo osciló entre 7,2 y 8,7 en invierno/12 (S3C y S1C). En el arroyo Mista el menor y mayor registro se dieron en invierno/12 con 7,7 en S3M y 9,0 en S1M y S2M. Se observó una tendencia, en general, decreciente de los registros de esta variable hacia aguas abajo en ambos sistemas lóticos.
Los registros mínimos y máximos de conductividad eléctrica para el Calimayo fueron de 150 µS/cm en S1C (verano/14) y de 2572 µS/cm S2C (invierno/12). El menor valor en Mista fue de 1450 µS/cm en S2M (verano/14) y el máximo registro fue de 5435 µS/cm en S3M (invierno/13).
En el arroyo Calimayo los tenores de oxígeno variaron entre 0,06 y 10 mg/L (en S3C durante casi todo el período de estudio y S1C otoño/13, respectivamente). Mientras que en las aguas del arroyo Mista el OD fluctuó de 0,06 mg/L a 15 mg/L, durante el invierno/12 en S3M y S1M, respectivamente.
La menor concentración de DBO5 en el Calimayo se observó en el S1C durante verano/13 (0,30 mg/L) y la máxima en S2C para invierno/13 (738 mg/L). En el Mista esta variable fluctuó entre 0,40 y 95 mg/L en S1M (verano/13) y S3M (invierno/12), respectivamente. En la tabla 1 se presentan los valores obtenidos de las diferentes variables fisicoquímicas para ambos arroyos (Tabla 1).
Las aguas del arroyo Calimayo según el diagrama de Piper-Hill-Langelier fueron bicarbonatadas-cálcicas en el S1C, mientras que en S2C y S3C se clasificaron como cloruradas-sódicas. En el caso del Mista las aguas fueron cloruradas-sulfatadas-sódicas para todos los sitios y períodos estudiados.
La riqueza específica para ambos arroyos fue de 61 taxones, 15 fueron comunes a los dos sistemas, 10 se registraron exclusivamente en el Calimayo y 36 se observaron solamente para el Mista (Tabla 2).
En relación a la distribución temporal de la riqueza de especies del arroyo Calimayo se pudo apreciar que el menor registro se dio en el otoño/14 con 2 taxones para S2C y el mayor número se determinó para el invierno de 2012 en el S1C con 17. El género con más especies correspondió en el Calimayo a: Gomphonema (4). Mientras que para el arroyo Mista la menor riqueza se presentó en S2M con 14 taxones en el otoño/13 y el máximo fue de 39 en el invierno/12 en S3M. Los géneros que aportaron el mayor número de especies en este sistema fueron: Navicula y Tryblionella con 7 y Gomphonema y Nitzschia con 6.
El arroyo Mista presentó valores mayores de riqueza (49 taxones) comparado con el arroyo Calimayo (25 taxones) y los máximos registros no coincidieron ni en sitios ni temporalmente.
En el arroyo Calimayo la menor densidad correspondió al S3C en el otoño/14 con 19 ind/ml, mientras que la máxima se presentó en el S1C con 227 ind/ml durante el invierno/12. En el sistema lótico Mista la mínima abundancia fue de 6 ind/ml en el verano/14 en S2M, mientras que el máximo registro se observó en S3M con 228 ind/ml durante agosto/13.
Con respecto al índice de diversidad y sus intervalos de confianza asociados (Fig. 2), no se observaron diferencias de los valores máximos registrados entre los arroyos muestreados. Para el caso del arroyo Calimayo, los datos de diversidad más elevados y significativos comparados con sitios y fechas correspondieron al sitio 1 en agosto (invierno) de 2012 y 2013 junto con noviembre (primavera) de 2013. Mientras que para el arroyo Mista fue el sitio 3 en noviembre (primavera) de 2012 y mayo (otoño) de 2014.
En el arroyo Calimayo mediante el diagrama de Olmeasted Tuckey ocho especies resultaron ser dominantes por su alta densidad y frecuencia: Amphipleura lindheimerii, Cocconeis placentula var. euglypta, Cocconeis placetula var. lineata, Diatoma vulgare, Gomphonema parvulum, Nitzschia palea, Reimeria uniseriata y Ulnaria ulna. Las especies constantes fueron tres: Gomphonema truncatum, Pinnularia acrosphaeria y Surirella ovalis. Los taxones ocasionales fueron seis: Cymbella cymbiformis, Gomphonema pseudoaugur, Navicula tripunctata, Nitzschia recta, Rhoicosphaenia abbreviata y Surirella solea. Los taxones raros fueron ocho: Achnanthes brevipes, Cymbella affinis, Gomphonema affine, Luticola mutica, Melosira varians, Navicula radiosa, Nitzschia linearis y Sellaphora pupula (Fig. 3A).
En el arroyo Mista se reconocieron 12 especies dominantes: Amphora ovalis, Anomoeonoeis sphaerophora, Cocconeis placentula var. euglypta, Cyclotella meneghiniana, Entomoneis alata, Gomphonema affine, Nitzschia obtusa, Pleurosigma elongatum, Rhopalodia gibba, Surirella ovalis, Surirella rorata y Ulnaria ulna. Los taxones constantes fueron seis: Gomphonema parvulum, Cosmioneis brasiliana, Parlibellus crucicula, Navicula peregrina, Pleurosira laevis y Tryblionella constricta. Las ocasionales estuvieron representadas por 17 taxones: Campylodiscus clypeus, Cocconeis placentula var. lineata, Craticula cuspidata, Cymbella turgidula, Gomphonema pseudoagur, Halamphora coffeaeformis, Hippodonta capitata, Luticola mutica, Navicula cryptocephala, N. meniscus, N. radiosa, N. tripunctata, Nitzschia palea, Pinnularia acrosphaeria, Tryblionella levidensis var. levidensis, T. littoralis y T. salinarum. Mientras que los taxones raros fueron 14: Diatoma vulgaris, Diploneis boldtiana, D. subovalis, Encyonema silesiacum, Gomphonema affinopsis, G. mexicanum, G. truncatum, Halampora veneta, Nitzschia acicularis, N. linearis, N. reversa, Tryblionella calida, T. compressa y T. hungarica (Fig. 3B).
En el arroyo Calimayo considerando los valores del índice sapróbico (IS) (1,6-1,8) las aguas en S1C se caracterizaron como oligo-ß-mesosapróbica indicando una contaminación débil y según el IDPm este sitio presentó valores que oscilaron entre 1,1 a 1,5, marcando una buena calidad de agua. Mientras que en S2C y S3C con registros de 2,9-3,4 del IS y 2,4-3,5 en el IDPm, estos índices denotaron aguas dentro del rango α-polisapróbicas, con contaminación fuerte y aguas de mala calidad (Fig. 4).
En el arroyo Mista los datos del IS caracterizaron a S1M como oligo-ß-mesosapróbico, con valores de 1,6 a 1,9 y según el IDPm la calidad del agua en S1M fue aceptable (1,8-2). En el caso de los S2M y S3M el IS los determinó como ß-mesosapróbicos, con valores de 2-2,3 y 2-2,6, respectivamente. Asimismo, el IDPm los catalogó con valores que oscilaron entre 2-2,6, indicando una mala calidad del agua en esos puntos (Fig. 5).
En base al ACC los ejes 1 y 2 acumularon el 68% de la varianza explicada (F= 2.83, p <0.05, chi cuadrado=2.13, Fig. 6). La tabla 3 muestra las variables ambientales que resultaron significativas mediante pruebas de permutación de Monte Carlo y que habrían favorecido las mayores abundancias de 11 especies de diatomeas: Melosira varians (Meva); Achnanthes brevipes (Acbr); Luticola mutica (Lumu); Navicula radiosa (Nara); Cymbella affinis (Cyaf); Rhoicosphaenia abbreviata (Rhab); Sellaphora pupula (Sepu); Cymbella cymbiformis (Cycy); Surirella solea (Cyso); Nitzschia recta (Nire); Reimeria uniseriata (Reun). Así, en la estación de muestreo 2 del arroyo Calimayo a menores valores de pH, CE, OD, Na+ y Ca2+ y a mayores valores de temperatura y NH4+ se registraron las mayores abundancias de las especies mencionadas.
Discusión y Conclusiones
En los sitios de referencia (sitios 1) se observó una fluctuación de la temperatura relacionada con la variabilidad estacional, con valores mínimos en otoño-invierno y los más elevados en verano; sin embargo, es necesario destacar que en los sitios afectados por efluentes polutos la temperatura fue siempre más elevada que en los sitios 1 de ambos arroyos, independientemente de la estacionalidad. Esta situación también se observó en otros ambientes lóticos de Argentina como en el río Gastona (Tucumán, Mirande, 2001) y en el arroyo Don Gato, tributario del río de La Plata (Buenos Aires, Gómez et al., 2008).
La alcalinidad de los arroyos estudiados se mantuvo durante el período analizado lo que coincide con datos de la cuenca del río Juramento en Salta (Salusso, 2005), en los ríos Tapia y Vipos (Martínez De Marco, 2009) en Tucumán, entre otros. El pH presentó un decrecimiento marcado en los sitios ubicados aguas abajo, lo que podría estar asociado a la descomposición de la carga orgánica vertida por los efluentes polutos o a un incremento en la concentración de sulfatos o cloruros, situación concordante con lo encontrado por Plataroti (2010) en el río Luján que se encuentra afectado por descargas contaminantes.
El agua del sitio 1 del arroyo Calimayo presentó una mineralización media-acentuada, lo que concuerda con diversos sistemas lóticos como por ejemplo con lo encontrado por Martínez & Donato (2003) para el río Tota (Colombia), en Tucumán por Martínez De Marco & Tracanna (2012) en el río Vipos. Todos los sitios del Mista y S2C-S3C del Calimayo tuvieron mineralizaciones elevadas (más de 1.000 µS /cm), lo que estaría asociado a factores como las bajas precipitaciones en ciertas épocas del año, la litología y también a la descarga de contaminantes que alteran el contenido de iones y materiales disueltos provocando las discontinuidades registradas tanto a nivel espacial como temporal. Estos elevados registros de conductividad ya fueron determinados por González & Domínguez (1994), Valz-Gianinet et al. (1999) y Powell (2011) en el arroyo Calimayo al analizar el impacto de efluentes polutos sobre la biota. Este comportamiento también se observó en distintos ríos y arroyos del país sometidos a diferentes tipos de polución por ej. ríos Luján, Reconquista, Matanza-Riachuelo, entre otros (Castañe et al., 1998; Lombardo et al., 2009; Plataroti, 2010).
En los tres sitios del arroyo Mista (excepto en S3M en invierno/12) y el sitio 1 del Calimayo las aguas estuvieron oxigenadas con leves variaciones y las concentraciones de DBO5, fueron bajas. Esta conjunción de alto contenido de OD y bajo DBO5 coincidió con lo mencionado en Tucumán para algunos ríos de Tafí del Valle (Isasmendi et al., 2007), en Salta para los ríos Arenales y Arias (Saluso & Moraña, 2002), en Córdoba para las redes hídricas Piedra Blanca, Chocancharava y Chucul (Luque & Martínez de Fabricius, 2005; Novoa, 2014). Los tenores de oxígeno disuelto en los sitios 2 y 3 del arroyo Calimayo y el S3M (invierno/12) fueron muy bajos denotando condiciones de anoxia lo que podría indicar que la carga orgánica de origen industrial vertida en estos tramos excede la capacidad de autodepuración y los niveles de oxígeno responden principalmente a la DBO5, tal como fuera descripto por Arreghini (2008) para el río Reconquista. Probablemente, el vertido proveniente de la industria papelera en esos sectores del Calimayo generó un aporte de sustancias que no pudo ser biodegradado totalmente. Estos efluentes causan diferentes tipos de impacto ambiental, los cuales están asociados a su contenido de sólidos en suspensión y carga orgánica.
En el caso del arroyo Mista, particularmente en el sitio 3, su estado se podría deber a la producción del ingenio azucarero, que elimina residuos orgánicos (cachaza y vinaza), que ocasionan anoxia, elevada demanda bioquímica de oxígeno y disminución del pH entre otros disturbios, situación que concuerda con lo encontrado en los ríos Salí y Gastona (Mirande, 2001; Seeligmann et al., 2001).
Mientras que el sitio 1 del Calimayo se corresponde por su litología a aguas bicarbonatadas, en los otros sitios de este arroyo, el agua se clasificó como clorurado-sódicas, lo que fue observado también en la década del 90 por Padilla et al. (1996) y podría relacionarse con la utilización de hipoclorito de sodio en la producción del papel (Villalonga, 2006).
En el arroyo Mista el agua fue clorurado-sulfatado-sódica en todos los sitios y épocas estudiadas lo que coincide con lo propuesto por Zuccardi & Fadda (1985) quienes mencionan que la naturaleza salina del suelo sobre la que discurre el arroyo estaría aportando gran cantidad de sodio (entre 350-20.000 mg/L) datos que coinciden con los encontrados en este trabajo.
Las concentraciones de nitrato en los sitios 2 y 3 del Calimayo fueron elevadas cercanas al valor límite, registros similares se encontraron en distintos ríos: Santa Lucía, Uruguay (Mendes et al., 1988), Matanza-Riachuelo (Buenos Aires) (Goméz et al., 2008), Salí (Seeligmann et al., 2001). En el caso de los sitios del arroyo Mista cabe mencionar que se encuentran cercanos a cultivos de caña de azúcar y que es probable que el uso de agroquímicos podría incrementar los niveles de nitrato, como así también se debe considerar los efluentes del ingenio azucarero.
La riqueza de diatomeas fue, en general elevada, similar a lo encontrado en otros cursos del país, por ejemplo en los ríos: Piedra Blanca, Chucul, Santa Catalina, Río Cuarto (Córdoba), alta cuenca del Bermejo (Salta), en el Gastona, Vipos y Tapia (Tucumán) (Mirande, 2001; Luque & Martínez de Fabricius, 2005; Martínez De Marco, 2009; Martínez de Fabricius et al., 2010; Martínez De Marco et al., 2011; Novoa, 2014). El menor número de especies se observó en el arroyo Calimayo y el mayor en el Mista. Hubo ciertas diferencias estacionales entre los arroyos, la máxima riqueza en el Calimayo se obtuvo en invierno y la mínima en el verano, mientras que en el Mista fue más elevada en verano y menor en otoño. Las variedades, Cocconeis plancetula var. euglypta, C. placentula var. lineata y las especies Gomphonema parvulum, G. pseudoaugur, Melosira varians, Surirella solea y Ulnaria ulna fueron dominantes en las muestras analizadas y han sido previamente citadas en ambientes eutrofizados a hipereutrofizados, lo que da un indicio de un alto grado de deterioro en los sistemas considerados (Pizarro & Almanni, 2005; Esquiús et al., 2008; Mengo, 2017).
Se pudo observar que la diversidad varió siguiendo una dinámica anual-interanual y según el sitio de descarga de los efluentes contaminantes. En el Calimayo la mayor diversidad, en general, se observó en el sitio aguas arriba y en el Mista preferente en aguas debajo de la descarga (S3M).
La abundancia de diatomeas fue máxima durante el invierno y la primavera en ambos arroyos, pero en diferentes años y los menores valores se registraron en verano-otoño, coincidiendo con lo observado en otros cuerpos de agua (Mirande, 2001; Salusso, 2005). En los sitios impactados las mayores densidades observadas en invierno coincidieron con elevados registros de DBO5 lo que denota una mala calidad en los puntos de descarga o aguas abajo. Es necesario destacar que en la época invernal ocurre la mayor actividad agroazucarera y agroindustrial de la zona (Medwid, 2008).
En base al diagrama de Olmeasted & Tukey se pudo observar que la composición de diatomeas para cada arroyo fue diferente. Las especies dominantes fueron 8 y las constantes solo 3 en el Calimayo, mientras que el Mista fueron 12 dominantes y constantes 6, situación semejante a lo citado para ríos de México por Varona Cordero & Gutiérrez Mendieta (2003). Sólo dos taxones coincidieron en ambos arroyos como dominantes: Cocconeis placentula var. euglypta y Ulnaria ulna. Entre las otras especies dominantes algunas fueron propias de cada arroyo, mientras que otras cambiaron su status ya sea a constante, a ocasional o a rara sin apreciarse una diferenciación por estación de muestreo, por año o por estación seca versus época húmeda.
Se pudo apreciar que el tipo de agua en los sitios 1 de los dos sistemas fueron determinadas como oligo-ß-mesosapróbica denotando una buena calidad, esta tipificación fue respaldada por el índice de diatomeas pampeano modificado (IDPm), según el cual estos lugares presentaron un buen estado, con polución y eutrofización mínimas y un leve grado de disturbio, datos que coinciden con las características mencionadas para ríos serranos de Córdoba (Martínez de Fabricius et al., 2010), para el arroyo La Tapera en Buenos Aires (Esquiús et al., 2008) entre otros. Esta condición se modificó en el resto de los sitios, tanto de descarga (S2) como aguas abajo (S3) donde presentan el rango α-polisaprobicas para el arroyo Calimayo, mientras que ß-mesosapróbico para el Mista. El mal estado de los sitios más poluidos se vio también afectado por la estacionalidad de la región, incrementando sus malas condiciones en las épocas de sequía. Los valores elevados del IS e IDPm en los sitios más perturbados concuerdan con lo citado para los ríos de la cuenca Matanza-Riachuelo en Buenos Aires y La Plata por Licursi & Gómez (2003). Cabe resaltar que el arroyo Calimayo en la estación de descarga (S2) a menores valores de pH, CE, OD, Na+ y Ca2+ y a mayores valores de temperatura y NH4+ se observaron incrementos en las abundancias de 11 especies que en otras condiciones se registraron como ocasionales o raras (resultado del método multivariado ACC). Se destaca que Reimeria uniseriata especie propia del Calimayo, independientemente de condiciones anteriores o posteriores a las descargas de efluentes contaminantes, estuvo en condición dominante lo que determinaría que se comportó como tolerante para esas condiciones ambientales. El ACC realizado permitió observar una relación entre las variables fisicoquímicas y el ensamble de diatomeas, que favoreció la presencia y desarrollo de diversos taxones los cuales fueron mencionados en varios ambientes con un amplio rango de polución.
De los resultados obtenidos se detectaron dos patrones de fluctuación: uno espacial asociado a la acción antropogénica y uno temporal relacionado con el clima y la hidrología de cada arroyo. La existencia de patrones de variación temporal y estacional también se asocian con las condiciones físicoquímicas que se modifican debido al incremento de actividad agroindustrial que provocan el deterioro de la calidad del agua y el aumento del estado trófico en los cursos hídricos estudiados, lo que se refleja en la variación de la composición y abundancia diatomológica.
El ensamble de diatomeas resultó ser una taxocenosis bioindicadora sensible del estado de los arroyos, coincidiendo con numerosos trabajos realizados en diversas partes de Argentina y del mundo. Es necesario destacar que estos arroyos aportan gran cantidad de materia orgánica y efluentes polutos a la cuenca Salí-Dulce de manera que incrementan la contaminación y el deterioro de esta importante red hídrica del NOA.
En este trabajo se registraron un total de 61 especies de las cuales 11 en su dinámica pasaron de ser ocasionales o raras a dominantes según sus abundancias, lo que se observó en el ACC por lo que serían buenas indicadoras del estado de contaminación.
Esta contribución es de relevancia si se considera, coincidiendo con Domínguez et al. (2022), que en el territorio argentino, tan amplio y diverso no se puede utilizar un único tipo de índice en bioindicación, sino que es necesario generar y ajustar los índices desarrollados en cada una de las regiones. El empleo del índice de diatomeas pampeano modificado, creado en base a datos propios, resultó más sensible y eficaz en la determinación del estado ecológico. De manera que podría ser una herramienta diagnóstica, de seguimiento de la calidad del agua y planificación que serviría para brindar información válida y de fácil aplicación en cuencas del Noroeste de Argentina.
Agradecimientos
A las Dras. Beatriz Tracanna y Silvia Martínez De Marco por su dirección, guía, acompañamiento y enseñanzas en la temática de la ficología.

Tabla 1 Variables fisicoquímicas y biológicas medidas en ambos arroyos: CE: Conductividad eléctrica (µS/cm), Tem: Temperatura (°C), OD: Oxígeno disuelto (mg/L), DBO5: Demanda bioquímica de oxígeno (mg/L), las restantes unidades se midieron como mg/L. N/D: No detectado.
| pH | CE | Te | OD | DBO5 | HCO3 - | CO2- 3 | SO2 4 | Cl- | Na+ | K+ | Ca2+ | Mg2+ | NO3 - | NO2 - | NH4 + | PO4 3 | |
| S1C lnv-12 | 8,7 | 631 | 14 | 9,2 | 0,6 | 213 | 108 | 5,7 | 18 | 23 | 3,9 | 80 | 15 | 2,2 | 0,01 | 0,05 | 0,05 |
| S2C lnv-12 | 7,3 | 2572 | 23 | 0,1 | 338 | 793 | N/D | 192 | 294 | 368 | 46,8 | 134 | 15,6 | 1,53 | 0,03 | 0,44 | 0,62 |
| S3C lnv-12 | 7,2 | 2000 | 19 | 0,1 | 134 | 579 | N/D | 144 | 265 | 299 | 35,8 | 80 | 28,2 | 1,12 | 0,1 | 0,55 | 0,75 |
| S1M lnv-12 | 9 | 2620 | 17 | 15 | 3,5 | 439 | 168 | 408 | 276 | 506 | 19,5 | 40 | 20 | 0,01 | 0,02 | 0,05 | 0,1 |
| S2M lnv-12 | 9 | 3440 | 13 | 12 | 2,1 | 500 | 168 | 672 | 336 | 667 | 21 | 40 | 27,9 | 0,02 | 0,01 | 0,05 | 0,15 |
| S3M lnv-12 | 7,7 | 4820 | 12 | 0,1 | 94,8 | 640 | 510 | 1128 | 510 | 851 | 35,8 | 140 | 32 | 0,93 | 0,01 | 0,31 | 0,4 |
| S1C Prim-12 | 8,3 | 470 | 20 | 7,1 | 0,6 | 195 | 48 | 4 | 24,8 | 17 | 5,07 | 39,9 | 20 | 2,24 | 0,02 | 0,06 | 0,19 |
| S2C Prim-12 | 7,8 | 2030 | 27 | 0,6 | 116,1 | 610 | N/D | 100,8 | 291 | 330 | 65,1 | 33,4 | 15 | 1,25 | 0,02 | 0,67 | 0,29 |
| S3C Prim-12 | 7,4 | 1933 | 24 | 0,1 | 248 | 579,5 | N/D | 78 | 291 | 399 | 33,9 | 60 | 36 | 8,75 | 0,03 | 1,44 | 0,85 |
| S1M Prim-12 | 8,4 | 2640 | 23 | 9,2 | 0,8 | 497 | 138 | 401 | 269 | 469 | 23 | 33 | 29,6 | 1,12 | 0,01 | 0,07 | 0,2 |
| S2M Prim-12 | 8,6 | 3160 | 21 | 12 | 2,6 | 378 | 264 | 557 | 333 | 540 | 24 | 46,6 | 18 | 0,37 | 0,01 | 0,03 | 0,2 |
| S3M Prim-12 | 8,3 | 4650 | 22 | 5,8 | 9,6 | 408 | 126 | 1037 | 576 | 845 | 30,8 | 113 | 43 | 0,8 | 0,01 | 0,22 | 0,2 |
| S1C Ver-13 | 8,4 | 280 | 20 | 7,9 | 0,3 | 103,7 | 24 | 9,61 | 17,7 | 8,9 | 3,9 | 26,6 | 12 | 1,34 | 0,07 | 0,07 | 0,12 |
| S2C Ver-13 | 7,9 | 1620 | 25 | 1 | 119,1 | 433,1 | N/D | 110 | 241 | 230 | 12,9 | 73,4 | 12 | 1,08 | 0,3 | 0,54 | 0,84 |
| S3C Ver-13 | 7,9 | 1400 | 24 | 0,1 | 94,12 | 433 | N/D | 72 | 191 | 190 | 12,9 | 66,6 | 15,96 | 2,17 | 0,24 | 0,48 | 0,82 |
| S1M Ver-13 | 8,6 | 2520 | 23 | 8,6 | 0,4 | 439 | 84 | 456 | 251 | 460 | 15,9 | 36 | 23 | 0,01 | 0,01 | 0,05 | 0,2 |
| S2M Ver-13 | 8,7 | 3380 | 23 | 7,1 | 1 | 439 | 108 | 710 | 354 | 600 | 17,9 | 40 | 24 | 0,02 | 0,01 | 0,21 | 0,4 |
| S3M Ver-13 | 8,5 | 4880 | 24 | 5,9 | 3,3 | 305 | 66 | 1219 | 609 | 719 | 23 | 146 | 27,9 | 1,2 | 0,02 | 0,49 | 0,5 |
| S1C Oto-13 | 8,6 | 476 | 13 | 9,5 | 0,9 | 183 | 72 | 2,8 | 17,7 | 169 | 3,9 | 46,6 | 20 | 0,53 | 0,02 | 0,05 | 0,17 |
| S2C Oto-13 | 7,6 | 2250 | 22 | 0,5 | 263 | 707 | N/D | 96 | 315 | 310 | 23 | 113 | 20,04 | 0,63 | 0,07 | 0,42 | 0,2 |
| S3C Oto-13 | 7,3 | 1880 | 19 | 0,1 | 275,9 | 610 | N/D | 67 | 262 | 270 | 19,8 | 100 | 20 | 0,59 | 0,06 | 0,75 | 0,51 |
| S1M Oto-13 | 8,5 | 2580 | 14 | 12 | 2 | 445 | 192 | 350 | 283 | 480 | 15,9 | 42 | 24 | 0,33 | 0,02 | 0,05 | 0,4 |
| S2M Oto-13 | 8,7 | 3280 | 13 | 7,7 | 1 | 402 | 240 | 585 | 354 | 609 | 17,9 | 40 | 20 | 0,02 | 0,01 | 0,05 | 0,3 |
| S3M Oto-13 | 8,4 | 5400 | 14 | 9,4 | 0,7 | 219 | 192 | 1344 | 684 | 700 | 21,8 | 126 | 31,2 | 0,24 | 0,02 | 0,4 | 0,2 |
| S1C lnv-13 | 8,5 | 600 | 14 | 10 | 1,04 | 73 | 204 | 144 | 39 | 23 | 3 | 60 | 27,6 | 2,19 | 0,01 | 0,3 | 0,85 |
| S2C lnv-13 | 7,3 | 2300 | 22 | 0,1 | 738,3 | 701,5 | 36 | 168 | 262 | 339 | 37 | 86 | 3,9 | 1,37 | 0,04 | 1,6 | 1,5 |
| S3C lnv-13 | 7,4 | 1900 | 19 | 0,1 | 578,3 | 530 | 72 | 120 | 234 | 270 | 28 | 93 | 15,9 | 0,8 | 0,05 | 1,6 | 1,34 |
| S1M lnv-13 | 8,2 | 3660 | 15 | 12 | 1,2 | 6,1 | 354 | 835 | 468 | 675 | 18 | 66 | 15,9 | 3,9 | 0,01 | 0,62 | 0,3 |
| S2M lnv-13 | 8,4 | 3465 | 18 | 8,7 | 2,2 | 390 | 216 | 655 | 390 | 670 | 19 | 26 | 15,9 | 0,6 | 0,01 | 0,19 | 0,4 |
| S3M lnv-13 | 8,2 | 5435 | 14 | 11 | 5,8 | 213 | 192 | 1392 | 640 | 1100 | 23 | 133 | 42 | 1,14 | 0,02 | 0,6 | 0,3 |
| S1C Pri-13 | 8,2 | 670 | 21 | 7,6 | 1,07 | 280 | 72 | 20 | 28 | 28 | 3 | 90 | 11 | 3,07 | 0,02 | 0,05 | 0,25 |
| S2C Pri-13 | 7,4 | 2036 | 28 | 0,3 | 438,5 | 457 | 72 | 350 | 372 | 350 | 19 | 85 | 20 | 0,54 | 0,04 | 0,3 | 1,01 |
| S3C Pri-13 | 7,5 | 1865 | 26 | 0,1 | 163,5 | 366 | 72 | 379 | 322 | 379 | 20 | 86,6 | 10,4 | 0,64 | 0,01 | 1,9 | 1,03 |
| S1M Pri-13 | 8,1 | 2634 | 22 | 8,7 | 2,7 | 585 | 84 | 480 | 393 | 480 | 15 | 50 | 9,6 | 0,17 | 0,03 | 0,1 | 0,3 |
| S2M Pri-13 | 7,8 | 3731 | 25 | 7,6 | 2,4 | 457 | 24 | 609 | 620 | 609 | 16 | 78 | 11 | 0,41 | 0,02 | 0,05 | 0,5 |
| S1C Ver-14 | 8,1 | 150 | 21 | 7,3 | 1,6 | 36 | 12,4 | 5 | 17,7 | 6,9 | 3 | 13,4 | 4,8 | 1,63 | 0,02 | 0,05 | 0,24 |
| S2C Ver-14 | 7,7 | 1040 | 30 | 0,7 | 223 | 506 | N/D | 95 | 74,5 | 97,9 | 14 | 80 | 14 | 0,32 | 0,03 | 0,1 | 0,32 |
| S3C Ver-14 | 7,6 | 1080 | 25 | 0,1 | 153 | 445 | N/D | 65 | 103 | 95,9 | 13 | 80 | 12,8 | 4,7 | 0,03 | 1,14 | 0,19 |
| S1M Ver-14 | 8,1 | 2150 | 25 | 5 | 2,8 | 341 | 68 | 244 | 305 | 410 | 21 | 46 | 14 | 4,2 | 0,02 | 0,05 | 0,5 |
| S2M Ver-14 | 8,2 | 1450 | 23 | 5 | 2,6 | 274 | 37 | 124 | 220 | 290 | 17 | 33 | 6,3 | 1,7 | 0,02 | 0,05 | 0,7 |
| S3M Ver-14 | 8 | 2700 | 24 | 4,3 | 5,9 | 311 | 37 | 417 | 426 | 439 | 19 | 66 | 15,9 | 4,4 | 0,02 | 0,05 | 0,5 |
| S1C Oto-14 | 7,9 | 200 | 17 | 8,4 | 1,7 | 79,3 | N/D | 4,8 | 21 | 5,9 | 3,9 | 21,4 | 10,4 | 2,23 | 0,02 | 0,05 | 0,05 |
| S2C Oto-14 | 7,9 | 1245 | 21 | 2,3 | 344 | 378 | N/D | 12 | 213 | 184 | 13 | 60 | 12 | 2,84 | 0,02 | 0,14 | 0,19 |
| S3C Oto-14 | 7,9 | 1170 | 20 | 2,1 | 254 | 353,8 | N/D | 24 | 192 | 160 | 12 | 66,6 | 15,2 | 1,8 | 0,02 | 1,1 | 0,3 |
| S1M Oto-14 | 8,5 | 2480 | 18 | 8,2 | 2,1 | 408 | 61 | 271 | 372 | 439 | 21 | 60 | 15,9 | 2,7 | 0,04 | 0,05 | 0,2 |
| S2M Oto-14 | 8,4 | 2820 | 18 | 7 | 1 | 500 | 55 | 307 | 418 | 520 | 19 | 46 | 22 | 1,4 | 0,05 | 0,05 | 0,3 |
| S3M Oto-14 | 8,3 | 5000 | 19 | 6,2 | 4,5 | 317 | 62 | 816 | 915 | 871 | 24 | 126 | 39 | 3,8 | 0,03 | 0,05 | 0,2 |
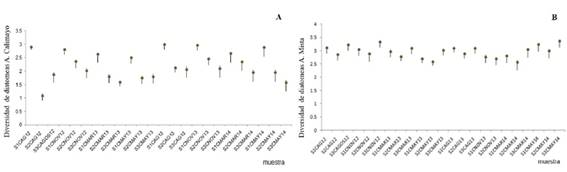
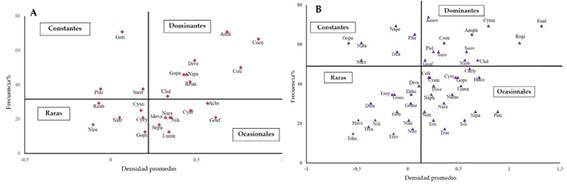
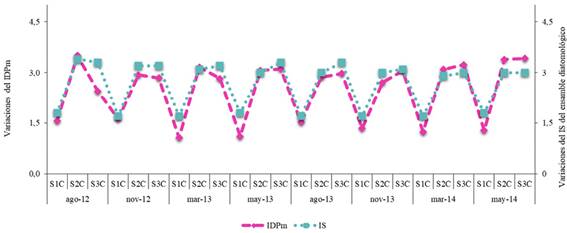
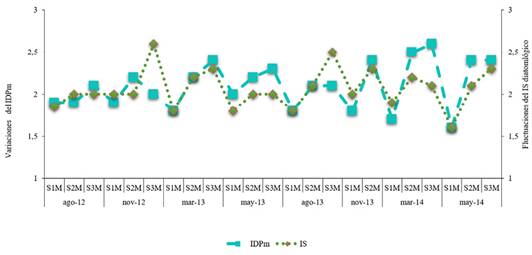

Fig. 6 Biplot ACC entre variables ambientales y ensamble de diatomeas: sitio 1 (antes de descarga), sitios 2 y 3 (después de descarga) durante cuatro estaciones: agos (invierno-agosto/2012), nov (primavera-noviembre/2012), mar (verano-marzo/2013) y may (otoño-mayo/2013).
Tabla 2 Riqueza diatomológica de ambos arroyos. S1C: ▲; S2C: ■; S3C: ●; S1M: ♠; S2M: ♦; S3M: ♣
| Taxones | Cod. | Presencia Sitios/Arroyos |
|---|---|---|
| Achnanthes brevipes C. Agardh | Acbr | ▲ |
| Amphipleura lindheimerii Kützing | Amp | ▲ |
| Amphora ovalis (Kützing) Kützing | Amov | ♠♦♣ |
| Anomoeoneis sphaerophora (Ehrenberg) E. Pfitzer | Ansph | ♠♦♣ |
| Campylodiscus clypeus (Ehrenberg) Ehrenberg ex Kützing | Cacly | ♠♦♣ |
| Cocconeis placentula var. euglypta (Ehrenberg) Grunow | Coeu | ▲■●♠♦♣ |
| Cocconeis placentula var. lineata (Ehrenberg) Van Heurck | Coli | ▲■●♠♦♣ |
| Cosmioneis brasiliana (Cleve) C. E. Wetzel & Ector | Nabr | ♠♦♣ |
| Craticula ambigua (Ehrenberg) D. G. Mann | Cram | ♠♣ |
| Cyclotella meneghiniana Kützing | Cyme | ♠♦♣ |
| Cymbella affinis Kützing | Cyaf | ▲ |
| Cymbella cymbiformis C. Agardh | Cycy | ▲■● |
| Cymbella turgidula Grunow | Cytu | ♠♦♣ |
| Diatoma vulgaris Bory | Divu | ▲■●♠♦♣ |
| Diploneis boldtiana Cleve | Dibo | ♠♦♣ |
| Diploneis subovalis Cleve | Disu | ♠♦♣ |
| Encyonema silesiacum (Bleisch in Rabenhorst) D. G. Mann | Ensi | ♦♣ |
| Entomoneis alata (Ehrenberg) Ehrenberg | Enal | ♠♦♣ |
| Gomphonema affine Kützing | Goaff | ▲■●♠♦♣ |
| Gomphonema affinopsis Metzeltin, H. Lange-Bertalot & F. García-Rodríguez | Gono | ♠♦♣ |
| Gomphonema gracile Ehrenberg | Gogr | ♠♦♣ |
| Gomphonema mexicanum Grunow | Gome | ♠♦♣ |
| Gomphonema parvulum (Kützing) Kützing | Gopa | ▲■●♠♦♣ |
| Gomphonema pseudoaugur Lange-Bertalot | Gops | ▲■●♠♦♣ |
| Gomphonema truncatum Ehrenberg | Gotr | ▲■●♠♦♣ |
| Halamphora coffeaeformis (C. Agardh) Levkov | Haco | ♦♣ |
| Halamphora veneta Kützing (Kützing) Levkov | Have | ♠♦♣ |
| Hippodonta capitata Patrick (Ehrenberg) Lange-Bertalot, Metzeltin & Witkowski | Hive | ♦♣ |
| Luticola mutica (Kützing) D. G. Mann | Lumu | ▲■●♠♦♣ |
| Melosira varians C. Agardh | Meva | ▲■● |
| Navicula cryptocephala Kützing | Naph | ♣ |
| Navicula meniscus Schumann | Name | ♠♦♣ |
| Navicula peregrina (Ehrenberg) Kützing | Nape | ♠♦♣ |
| Navicula radiosa Kützing | Nara | ▲■●♠♦♣ |
| Navicula tripunctata (O. F. Müller) Bory | Natr | ▲■●♠♦♣ |
| Nitzschia acicularis (Kützing) W. Smith | Niac | ♠♣ |
| Nitzschia linearis (C. Agardh) W. Smith | Nili | ▲■●♠♦♣ |
| Nitzschia obtusa W. Smith | Niob | ♠♦♣ |
| Nitzschia palea (Kützing) W. Smith | Nipa | ▲■●♠♦♣ |
| Nitzschia recta Hantzsch ex Rabenhorst | Nire | ▲ |
| Nitzschia reversa W. Smith | Nirv | ♠♦♣ |
| Parlibellus crucicula (W. Smith) Witkowski, Lange-Bertalot & Metzeltin | Nacr | ♠♦♣ |
| Pinnularia acrosphaeria W. Smith | Piac | ▲■●♠♦♣ |
| Pinnularia borealis Ehrenberg | Pibo | ♠♣ |
| Pleurosigma elongatum W. Smith | Plel | ♠♦♣ |
| Pleurosira laevis (Ehrenberg) Compére | Plla | ♠♦♣ |
| Reimeria uniseriata Sala, Guerrero & Ferrario | Reun | ▲■● |
| Rhoicosphenia abbreviata (C. Agardh) Lange-Bertalot | Rhab | ▲■ |
| Rhopalodia gibba (Ehrenberg) O. Müller | Rogi | ♠♦♣ |
| Sellaphora pupula (Kützing) Mereschkowksy | Sepu | ▲ |
| Surirella ovalis Brébisson | Suov | ▲■●♠♦♣ |
| Surirella solea (Brébisson) Brébisson | Cyso | ■● |
| Surirella rorata Frenguelli | Suro | ♠♦♣ |
| Tryblionella calida (Grunow) D. G. Mann | Trca | ♠♦♣ |
| Tryblionella compressa (J. W. Bailey) M. Poulin | Trco | ♠♦♣ |
| Tryblionella constricta (Kützing) Poulin | Trtr | ♠♦♣ |
| Tryblionella hungarica (Grunow) Frenguelli | Truh | ♠♦♣ |
| Tryblionella levidensis var. levidensis (W. Smith) Grunow | Trle | ♠♦♣ |
| Tryblionella littoralis (Grunow) D. G. Mann | Trli | ♠♦♣ |
| Tryblionella salinarum (Grunow) Pelletan | Trsa | ♠♦♣ |
| Ulnaria ulna (Nitzsch) Compère | Ulul | ▲■●♠♦♣ |
Tabla 3 Coeficientes de correlación inter conjunto de los ejes canónicos (ACC1, ACC2) y variables ambientales.
| Códigos de las variables ambientales | Coeficiente de correlación interset ACC1 | Coeficiente de correlación interset ACC2 |
|---|---|---|
| Temperatura (T) | 0,074 | -0,385 * |
| pH | -0,369 | 0,562 * |
| Oxígeno disuelto (OD) | -0,335 | 0,610 * |
| CE | -0,777* | -0,243 |
| Na+ | -0,812* | -0,211 |
| K | -0,356 | -0,573 |
| Ca2+ | -0,144 | -0,269 * |
| NH4+ | 0,275 | -0,583 * |
| PO43- | 0,265 | -0,433 |
| Arroyo Mista vs Calimayo | -0,989* | 0,051 |
| Estación 2 | -0,024 | -0,352* |
| Estación 3 | -0,203 | -0,293 |
| Estación húmeda vs estación seca | -0,089 | -0,105 |
| 2013 | -0,017 | -0,074 |
| 2014 | -0,025 | 0,016 |

