











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Las especies forrajeras perennes constituyen uno de los recursos más eficientes para la protección del suelo, ya que mejoran su fertilidad, disminuyen los costos por no requerir labranzas e incrementan los rendimientos agrícolas a través de la re- circulación de nutrientes (Pagliaricci et al., 2002). Por estas razones, es importante buscar nuevas alternativas forrajeras para desarrollar sistemas más productivos y sostenibles de producción animal. El gran desafío de los productores que practican una ganadería moderna consiste en incrementar la producción de carne y leche en forma acelerada y sostenible, de tal manera que permita garantizar la demanda de la población y la conservación de los recursos naturales y del ambiente (Giraldo, 1999).
En Argentina, la investigación en forrajes ha generado y producido gramíneas y leguminosas con potencial para aumentar la producción animal en sistemas de pastoreo (Lascano et al., 1996).
Los cultivos perennes utilizados en sistemas bovinos para carne y leche en el centro de Argentina son principalmente alfalfa y gramíneas de ciclo otoño-in- vierno-primavera, los que pueden sembrarse como cultivos puros o constituyendo praderas perennes (Pagliaricci et al., 1990). En la actualidad, las asociaciones de especies en una pastura son cada vez más frecuentes y, generalmente, comprenden la interrelación armónica y equilibrada entre dos o más especies de gramíneas y leguminosas. Estas asociaciones se pueden realizar con leguminosas nativas que se encuentran en el pastizal o con especies introducidas ya probadas (Sánchez, 1998). En la pampa húmeda y subhúmeda de Argentina, las gramíneas perennes de crecimiento otoño-invierno-primavera más cultivadas son la festuca alta (Festuca arundinacea), agropiro (Tynophiron ponticum), pasto ovillo (Dactylis glomerata), entre otras. La festuca alta es considerada la principal, debido a que es la mejor adaptada y de mayor producción forrajera en esta zona (Rimieri y Wolff, 2010). Esta especie se destaca por su resistencia a las condiciones invernales húmedas, así como su tolerancia a la sequía, haciendo de ella la gramínea por excelencia para suelos con limitaciones edáficas (Cullen, 1997). Es una especie C3, rústica y productiva, que ocupa un lugar muy significativo en las praderas y establecimientos ganaderos de
numerosos países (Rolhauser et al., 2007).
En la actualidad, las leguminosas asociadas a los pastizales nativos y el esta- blecimiento de praderas mixtas de gramíneas y leguminosas han sido ampliamente adoptadas para superar el déficit de Nitrógeno (N) del forraje en sistemas de pastoreo. La introducción de leguminosas en las praderas conduce a una mayor acumulación de N del suelo y una mayor productividad (Mortenson et al., 2004, Rao et al., 2007). Al evaluar la fijación biológica de Nitrógeno (FBN) en asociaciones de gramíneas y leguminosas, Reynolds (1982) determinó que la mayor parte del N cedido, alrededor de un 80%, se transfiere al suelo a través de la descomposición de raíces y nódulos o deyecciones de animales hasta ser asimilados por las plantas asociadas, mientras que otra porción significativa del N fijado se libera directamente al suelo por las exudaciones de las raíces hacia la rizósfera (Trannin et al., 2000; Spehn et al., 2002; Paynel et al., 2008).
De esta manera, las leguminosas no sólo incrementan el valor nutritivo de la gramínea asociada, particularmente en lo que se refiere a los contenidos de proteína total y de minerales, sino que además son capaces de aumentar la producción de materia seca en las praderas con una distribución estacional más uniforme y menor variabilidad interanual. Esta mayor disponibilidad de forraje permite elevar la carga animal por unidad de superficie (Rojas Hernández et al., 2005; Scheneiter, 2005), debido a que la consociación de gramíneas y leguminosas mejora el rendimiento de proteína y materia seca (Frankow-Lindberg et al., 2009; Nyfeler et al., 2011) y aumenta la fertilidad del suelo mediante el incremento de la materia orgánica y el contenido de N (Nguyen, 2003; Wichern et al., 2007). Al crecer de manera conjunta, las leguminosas y las gramíneas no compiten directamente por el N sino, por el contrario, la consociación de ambas puede llevar a un aumento significativo de la producción y el rendimiento de proteína (Cardinale et al., 2007; Kirwan et al., 2007; Frankow-Lindberg et al., 2009; Nyfeler et al., 2011). Por lo tanto, el uso de legumi- nosas puede disminuir la necesidad de utilizar fertilizantes nitrogenados, reducir los costos de producción y la contaminación del medio ambiente (Tekeli y Ateş, 2005). La búsqueda, introducción y persistencia de nuevas especies leguminosas nativas con potencial forrajero es posible cuando se conoce la biología (Izaguirre y Beyhaut, 1999), las estrategias de persistencia y las asociaciones con microorganismos del suelo (Bemhaja y Risso, 2006). Hace más de diez años que el equipo de Morfología Vegetal de la Facultad de Agronomía y Veterinaria (Universidad Nacional de Río Cuarto) estudia a Adesmia bicolor, debido a que es una leguminosa nativa, perenne, herbácea, de ciclo indefinido preferentemente invernal (Veneciano et al., 2005; Pe- reira Machín, 2011; Basconsuelo et al., 2013), tolerante a bajos niveles de Fósforo
(P) en suelo (Dodd y Orr, 1995) y con potencial forrajero y capacidad para la FBN
asociada con los rizobios nativos del suelo (Bianco et al., 2012; Basconsuelo et al., 2013; Bianco, 2014). Si bien durante estos últimos años se avanzó en los estudios del efecto de la competencia, de la consociación y dinámica de movilización de car- bohidratos de F. arundinacea y A. bicolor (Grassi et al., 2023) es interesante evaluar la capacidad de FBN que presenta A. bicolor creciendo consociada. Por ello, se propuso como objetivo de trabajo: cuantificar la biomasa seca total, el contenido de N y el número de nódulos producidos por A. bicolor consociada con F. arundinacea durante las fases vegetativa, de floración y de fructificación.
MATERIALES Y MÉTODOS
El ensayo se llevó a cabo en el Campo Experimental de la Facultad de Agronomía y Veterinaria (33º06’23.50’’ S, 64º17’54.41’’ O, 452 m snm), Universidad Nacional de Río Cuarto (UNRC), Río Cuarto, Córdoba, Argentina.
El suelo es un haplustol típico de textura franca arenosa muy fina y el clima es templado subhúmedo con régimen de precipitaciones tipo monzónico (Cantero et al., 1986). La precipitación media anual es de 779 mm y los mayores registros de lluvias ocurren entre octubre y marzo con picos en enero y diciembre, mientras que junio y julio son los meses más secos. Las temperaturas medias oscilan entre los 10 ºC en julio y los 24 ºC en enero (Cantero et al., 1986).
En una superficie de 15 m x 20 m se implantó Adesmia bicolor (L.) Poir. “Babo- sita” población Villa Rumipal-Córdoba (Fam. Leguminosae) en consociación con Festuca arundinacea Schreb. “Festuca alta” población 3302-LAG (Fam. Poaceae) (Grassi et al., 2023) (Fig. 1). Esta parcela se dividió en áreas de 2 m x 2 m, a las que se asignaron los distintos tratamientos, en los que se evaluaron cuatro tratamientos resultantes de la combinación de cuatro proporciones de leguminosa vs. gramínea:

Fig. 1 Parcela experimental con cultivo de Adesmia bicolor y Festuca arundinacea creciendo con- sociados. Año 2021. UNRC. Fig. 1Experimental plot with Adesmia bicolor and Festuca arundinacea cultures growing in asso- ciation. Year 2021. UNRC.
A1:F0 (Control I: A. bicolor, cultivo puro), A1:F1 (A. bicolor y F. arundinacea en proporciones similares), A3:F1 (A. bicolor y F. arundinacea en proporciones 3 a 1) y A0:F1 (Control II: F. arundinacea, cultivo puro). Estos tratamientos se dispusieron en franjas. En el arreglo se colocaron hileras paralelas de plantas a una distancia de 0,50 m. En la proporción 1:1, una hilera presentó plantas de F. arundinacea con coronas de 0,05 m de diámetro y la siguiente, champas de 0,05 m x 0,05 m de A. bicolor. En los demás casos, se ajustaron las hileras y las especies según los distintos tratamientos (Grassi et al., 2023).
Para evaluar la producción de biomasa seca total producida por A. bicolor y F. arundinacea se tomaron tres muestras al azar (0,25 m2) durante las diferentes etapas fenológicas (vegetativa, de floración y de fructificación) por tratamiento. El material vegetal fue secado en estufa a 65 °C durante 72 h. Posteriormente se determinó el contenido de N de parte aérea y subterránea (raíces y nódulos radicales) a través del método de Kjeldahl (Bremner y Mulvaney, 1982). Además, durante las mismas fases fenológicas, se contaron el número de nódulos en las raíces de A. bicolor. Los datos fueron analizados estadísticamente de acuerdo al arreglo factorial, y para las variables en las que se detectaron diferencias significativas se procedió a efectuar LSD-Fisher con Infostat (Di Rienzo et al., 2018).
RESULTADOS Y DISCUSIÓN
Biomasa seca total producida por A. bicolor consociada con F. arundinacea durante las fases de crecimiento
En la especie A. bicolor se observó que la mayor producción de biomasa seca total se produce en el cultivo puro (A1:F0). En la etapa de floración se observaron diferencias estadísticamente significativas con respecto al cultivo puro (A1:F0) y los consociados (A1:F1 y A3:F1) (p= 0, 0002) (Tabla 1).
En general la biomasa seca total de A. bicolor disminuyó lo largo de su creci- miento, con excepción del tratamiento de consociado A3:F1 en el cual se observó un incremento de biomasa nuevamente en la etapa de fructificación, con diferencias estadísticamente significativas de este tratamiento entre las fases de crecimiento (p= 0,046) (Tabla 1).
Estos resultados concuerdan con estudios realizados por Scheffer-Basso et al. (2002) donde se observó que en especies estoloníferas del género Adesmia, tales como A. latifolia y A. punctacta, el incremento de biomasa coincide con un mayor desarrollo de hojas. En A. bicolor el inicio del incremento en la producción de la biomasa total (aérea y subterránea) se registra a comienzos de la época estival (oc- tubre) coincidiendo con la ramificación de ramas cotiledonares y profilares y con aumento de la longitud de sus entrenudos (Pérez et al., 2020). Esto puede atribuirse a las mayores temperaturas de esta época del año y la distribución de lluvias, ya que las mayores precipitaciones ocurren entre octubre y marzo, con picos en diciembre y enero, siendo normalmente los meses más secos, junio y julio. La variabilidad de las lluvias en el área de estudio es muy grande, característica común de la mayoría de las regiones áridas y semiáridas de la Argentina (Veneciano et al., 2005).
En la especie F. arundinacea se observó en la etapa vegetativa una mayor produc- ción de biomasa seca total en el cultivo puro (A0:F1) con respecto a los consociados (A1:F1 y A3:F1), con diferencias estadísticamente significativas entre los tratamien- tos (p= 0, 009) (Tabla 1).
Tabla 1 Biomasa seca total de las especies en consociación a lo largo de las fases de crecimiento. Año 2021. UNRC. Letras minúsculas y mayúsculas diferentes en la misma columna, indican diferencias significativas entre los distintos tratamientos en cada fase de crecimiento y en cada tratamiento a lo largo fase de crecimiento, respectivamente (p < 0,05) según test LSD Fisher. Table 1. Total dry biomass of consociated species throughout growth stages. Year 2021. UNRC. Dif- ferent lowercase and uppercase letters in the same column indicate significant differences between the different treatments in each growth stage and in each treatment throughout the growth stages, respectively (p< 0.05) according to Fisher’s LSD test.
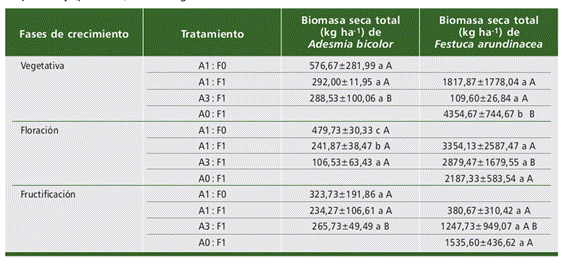
Por otro lado, se determinó que la producción de biomasa seca total en el culti- vo puro (A0:F1) disminuyó a lo largo de su ciclo de vida, siendo superior en etapa vegetativa y presentado diferencias estadísticamente significativas con respecto a las otras fases de crecimiento (p= 0,002).
En la etapa de floración la mayor producción de biomasa seca total fue en los tratamientos consociados (A1:F1 y A3:F1), si bien las diferencias no fueron estadís- ticamente significativas (p= 0,74) (Tabla 1).
Contenido de N de la biomasa aérea y subterránea de A. bicolor
y F. arundinacea consociadas durante las fases de crecimiento
El contenido de N de la parte aérea de A. bicolor presentó valores superiores en la etapa de floración, con respecto a la etapa vegetativa y de fructificación. En el trata- miento A3:F1 se registró el valor más alto de contenido de N (135,2 kg ha-1,) lo que mostró un efecto positivo de las especies consociadas (Tabla 2).
Con respecto al contenido de N en biomasa subterránea se observaron siempre valores inferiores que en la biomasa aérea. Los valores más altos de contenido de N fueron de 82,0 kg ha-1 en el cultivo puro (A1:F0) en etapa vegetativa y 78,4 kg ha-1 en el tratamiento consociado A3:F1 en floración (Tabla 2).
Tabla 2 Contenido de N de las especies en consociación a lo largo de fases de crecimiento. Año 2021. UNRC. Table 2. N content of consociating species throughout the growth stages. Year 2021. UNRC.
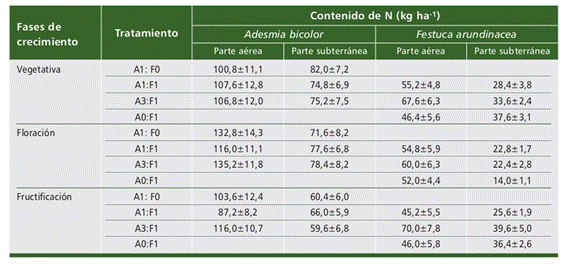
El contenido de N de la parte aérea de F. arundinacea mostró resultados muy variables. El tratamiento de consociados A3:F1 presentó el valor más alto de N en las tres etapas de crecimiento (67,6 kg ha-1 en etapa vegetativa, 60,0 kg ha-1 en floración y 70,0 kg ha-1 en fructificación) reflejando nuevamente una tendencia similar de las especies consociadas en esa proporción.
Al igual que en A. bicolor el contenido de N en parte subterránea mostró siempre valores inferiores que en la parte aérea. Todos los tratamientos presentaron valores menores de contenido de N en la etapa de floración. El valor más alto de N se ob- servó en el tratamiento de consociados A3:F1 en la etapa de fructificación (39,6 kg ha-1, Tabla 2).
En los resultados obtenidos se comprobó que en todos los tratamientos A. bico- lor presentó mayor contenido de N en la parte aérea que en la subterránea (raíces y nódulos), situación que coincide con estudios realizados en otras especies como A. latifolia (Scheffer Basso et al., 2001). Esto puede deberse a que en Fabáceas forrajeras de hábito de crecimiento estolonífero, la parte aérea presenta una mayor biomasa que la parte radical. Esta situación es diferente en especies rizomatosas donde la mayor cantidad de materia seca se encuentra en la raíz (Scheffer- Basso et al., 2002). En los tratamientos de consociaciones cabe resaltar que en general se obtuvieron valores mayores de N en relación a los cultivos puros, lo que coincide con estudios realizados en especies como Vicia villosa con Avena strigosa Sereb y Avena byzantina (Ferreira Casamayouret y Paniagua Alcaráz, 2013). Esto muestra un efecto benéfico en relación a la consociación gramínea-leguminosa. La combinación de las especies permite equilibrar la calidad de biomasa residual y mejorar la calidad del forraje. Además la gramínea aumenta su valor nutritivo y mejora su condición a través de la incorporación de N al suelo por medio de la fijación biológica que realiza la legu-
minosa asociada (Ferreira Casamayouret y Paniagua Alcaráz, 2013).
Cantidad de nódulos de A. bicolor consociada con F. arundinacea durante las fases de crecimiento
En la especie A. bicolor se observó que en todos los tratamientos la mayor produc- ción de nódulos se produce en la etapa vegetativa y que disminuyó notablemente a lo largo de las fases de crecimiento, presentando valores inferiores en la etapa de floración y fructificación (Fig. 2).
En función de los resultados se determinó que A. bicolor presenta mayor capa- cidad para fijar el N en simbiosis con los rizobios, principalmente en sus primeros estadios de vida, por la alta producción de biomasa seca total, contenido de N y número de nódulos.
A. bicolor a diferencia de otras leguminosas nativas presenta abundante nodu- lación, esto coincide con otras especies del género, tales como A. latifolia, así como otras leguminosas nativas, como Lupinus aureonitens, Rhynchosia hauthalli, Stylosanthes sp. y Trifolium leiocarpa (Scheffer-Basso et al., 2000; Westphal Muniz et al., 2012). El momento en que se produce el mayor número de nódulos coincide con estudios previos en otras especies del mismo género, como por ejemplo Adesmia araujoi, y especies de Fabáceas como Lupinus aureonitens, Rhynchosia hauthalli, Stylosanthes leio- carpa y Trifolium riograndense (Scheffer-Basso et al., 2000).

Fig. 2 Número de nódulos producidos por Adesmia bicolor consociada con Festuca arundinacea durante las diferentes fases de crecimiento (vegetativa, de floración y de fructificación). 2021. UNRC. Letras minúsculas diferentes indican diferencias estadísticamente significativas en cada tratamiento a lo largo de las fases de crecimiento (p < 0,05) según test LSD Fisher. Fig. 2Number of nodules produced by Adesmia bicolor associated with Festuca arundinacea during different growth stages (vegetative, flowering and fruiting). 2021. UNRC. Different lowercase letters indicate statistically significant differences in each treatment throughout the growth stages (p < 0.05) according to Fisher’s LSD test.
En A. bicolor, la disminución de la biomasa seca aérea y el número de nódulos después de la etapa vegetativa puede deberse a que la FBN en un primer momento promueve cambios morfo-fisiológicos en las plantas que resultan en la baja del creci- miento. Los nódulos representan un drenaje adicional de fotoasimilados y nutrientes, compitiendo con otros órganos de la planta (Cassman et al., 1980).
CONCLUSIÓN
Se concluye que A. bicolor es efectiva para fijar N tanto en el cultivo puro como consociada principalmente en sus primeros estadios de vida.
Teniendo en cuenta los valores de biomasa total de F. arundinacea y el contenido de N en ambas especies se observaron resultados prometedores en los tratamientos consociados A1:F1 y A3:F1, por lo cual se sugiere continuar con estudios que per- mitan comprender el comportamiento de esta asociación.

