











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkMás de 220 especies de aves completan su migración enteramente dentro de Sudamérica como parte del sistema austral del Neotrópico (Cueto y Jahn 2008), siendo este uno de los sistemas migratorios más diversos del mundo (Chesser 1994, Jahn et al. 2020). Sin embargo, la información disponible sobre la migración de varias de estas especies continúa siendo inexistente o, en el mejor de los casos, los ma-peos son de grano grueso (Faaborg et al. 2010).
La ciencia ciudadana -en este caso, participación de voluntarios en la recolección de registros de aves-proporciona una nueva oportunidad para estudiar patrones migratorios en aves sudamericanas al brindar una amplia base de datos de acceso libre (Schu-bert et al. 2019). Su uso para estudios de migración en aves se ha incrementado en los últimos años. Por ejemplo, investigaciones recientes utilizaron datos de ciencia ciudadana para describir: migraciones en especies poco conocidas (Lees y Martin 2014, Gorleri et al. 2021), patrones novedosos de migración (Lees 2016, DeGroote et al. 2020), migración parcial (Hayes et al. 2018), patrones fenológicos a escalas regionales (Gorleri y Areta 2022) y continentales (Schubert et al. 2019); o incluso para evaluar potenciales efectos del cambio climático sobre patrones migratorios futuros (Da Silveira et al. 2021). La gran ventaja que otorgan estas plataformas es la oportunidad de contar con gran cantidad de registros colectados a escalas geográficas amplias, lo cual permite inferir movimientos migratorios cuando se observan cambios en la distribución de los registros de una temporada a otra. No obstante, dado que los datos son colectados de manera no sistemática por voluntarios, es necesario realizar un apropiado manejo de los mismos para que las inferencias resultantes sobre la migración, o cualquier otro aspecto ecológico analizado, sean productos fiables y no sesgados a los sitios más muestreados.
La plataforma de ciencia ciudadana eBird (Su-llivan et al. 2014) colecta datos mediante protocolos semi-estructurados, es decir, protocolos que incorporan metadatos del muestreo (La Sorte et al. 2018). Esta información resulta útil para mitigar sesgos asociados a la recolección de datos de manera oportunista (Kelling et al. 2019). Por ejemplo, eBird incentiva la creación de listas completas de especies, donde el observador no solo reporta todas las especies que detecta durante el muestreo, sino también incluye información auxiliar del esfuerzo de muestreo realizado en cada lista (distancia recorrida, duración, número de participantes, etc.). Como resultado, es posible extraer información de presencia-ausencia de especies al verificar si una especie es reportada o no en una lista, además de obtener información del esfuerzo invertido en la obtención de cada registro (La Sorte et al. 2018). Esta información de alta calidad (datos de presencia-ausencia + metadatos del mues-treo) es necesaria para controlar los sesgos de muestreo, típicos de datos que son colectados sin seguir un protocolo sistemático (e.g. sesgo espacial, Johnston et al. 2021). En particular para el estudio de migración en aves, estos datos resultan útiles al ser sometidos a modelos probabilísticos que logran predecir cómo las especies cambian sus distribuciones a lo largo del año (Coxen et al. 2017, Gorleri et al. 2021).
Los tiránidos (Familia Tyrannidae) representan el 33% de las aves migratorias del sistema migratorio austral del Neotrópico (Chesser 1994, 1998), sin embargo, es uno de los grupos más complejos de estudiar. El Piojito Trinador (Serpophaga griseicapilla) es uno de estos tiránidos de ecología migratoria aún poco conocida, pero que podría beneficiarse de los datos actualmente disponibles en eBird. La especie cría desde Chubut por el oeste siguiendo la diagonal árida del desierto del monte hasta La Rioja (Bodrati y Ferrari 2014) y el oeste de Córdoba (Salvador et al. 2017), sin embargo, la extensión hacia el norte de su área de reproducción es controversial. Mientras algunos autores sostienen que cría exclusivamente en Argentina (Pearman y Areta 2020), otros mapean su rango reproductivo hasta el chaco de Bolivia (Herzog et al. 2016) y Paraguay (Lesterhuis et al. 2018). La situación durante el invierno parece ser más clara. Es señalado como visitante invernal regular en diversas localidades del chaco (Zyskowski et al. 2003, Di Giacomo 2005, Gorleri 2017, Coria et al. 2021) y litoral argentino (Fandiño y Giraudo 2010), y visitante ocasional en Uruguay (Fjeldsá 2020) y el sur de Brasil (Bencke et al. 2002, 2010, Bittencourt de Farias 2016). Aunque el Piojito Trinador es una especie frecuente, existe escasa información bibliográfica sobre su migración, lo cual puede deberse a ser una especie relativamente nueva para la ciencia (Straneck 1993, 2007), por ende, carente de registros históricos. No obstante, la ciencia ciudadana ha logrado compilar miles de registros del piojito en pocos años, muchos de ellos potencialmente informativos cómo para lograr iluminar su ecología migratoria.
En este trabajo, nuestro objetivo fue analizar la ecología migratoria del Piojito Trinador a fin de esclarecer su estatus regional mediante la utilización de los nuevos datos de eBird aplicados a modelos que predicen su migración y fenología. Específicamente, buscamos: (1) describir su área de cría y de invernada a resoluciones espaciales relativamente finas y con control del sesgo espacial en los datos, (2) conocer sus preferencias climáticas y de hábitat, y (3) examinar la sincronización o timing de su migración a escala regional.
Métodos
Compilación de datos
Creamos un set de datos de presencia-ausencia del Piojito Trinador para ser utilizado en los modelos de distribución y de fenología. Los datos fueron obtenidos exclusivamente de la plataforma eBird (eBird Basic Dataset 2021), dado que, a diferencia de otras plataformas diseñadas para la recolección de registros ornitológicos en Sudamérica, eBird permite crear un set de datos de presencia-ausencia a partir de las listas de especies (ver Bird et al. 2014, Guille-ra-Arroita et al. 2015). Para crear el set de datos de presencia-ausencia primero filtramos los registros de detección del Piojito Trinador subidos en listados completos desde el año 2010 a junio 2021. Luego inferimos como registros de ausencias a todos los listados completos donde no se reportó al piojito dentro del rango de fechas especificado. Este proceso llamado “zero-filling’ -o rellenado con ceros- lo llevamos a cabo en R utilizando el paquete auk (Strimas-Mackey et al. 2018).
Los datos de ciencia ciudadana exhiben amplia variabilidad en los modos de recolección de registros, de modo tal que es necesario estandarizar los datos para remover valores atípicos, por ejemplo, observaciones que resultan de listados de duración excesiva o con distancias recorridas muy largas. Filtramos al set de datos de presencia-ausencia para retener solo aquellos registros que cumplían con los siguientes requisitos: registros de listas completas que utilizaron el protocolo “estacionario” o “con desplazamiento”, con una duración no mayor a 5 horas, con una distancia recorrida no mayor a 5 km, y con 10 personas o menos participando del muestreo. Se demostró que estos criterios post hoc crean un set de datos más estandarizados a partir del set de datos original (Johns-ton et al. 2021).
Modelos de distribución
Estimamos la migración del Piojito Trinador evaluando el cambio en sus distribuciones de cría y de invernada mediante el uso de modelos de distribución de especies que utilizan los registros georreferencia-dos, asociados a capas climáticas y ambientales para predecir distribuciones (Guisan y Zimmerman 2000). Para ello, creamos un modelo de distribución del área de cría y otro del área de invernada y evaluamos el cambio observado en las predicciones entre ambas temporadas.
Para crear el modelo del área de cría utilizamos solo aquellos registros de presencia y ausencia del Piojito Trinador que datan de noviembre a febrero, meses en los cuales se ha corroborado la reproducción de la especie (Bodrati y Ferrari 2014, Salvador et al. 2017). En contraste, para el modelo de invernada utilizamos solo aquellos registros que datan de mayo a agosto, meses opuestos a los meses de reproducción. A modo de evitar utilizar registros de ausencias en regiones donde el piojito no se distribuye, limitamos los registros de ausencia al área potencial de distribución de la especie tanto para su área de cría como de invernada. Delimitamos el área potencial de distribución de cría y de invernada mediante la generación de dos polígonos convexos mínimos: uno englobando a todos los registros de cría y el otro a los registros de invernada. Utilizamos estos polígonos para la extracción de los registros de ausencia para cada temporada, por lo tanto, no utilizamos ausencias que caigan fuera de los límites de cada polígono. El set de datos resultante para el modelo de cría contó con 619 registros de presencias y 41 505 ausencias, el de invernada contó con 561 registros de presencia y 15 096 ausencias (ver Material Suplementario S1).
Realizamos un submuestreo de los registros para corregir el sesgo espacial en los datos y el desbalance de clases (ver Robinson et al. 2018). El sesgo espacial ocurre cuando las observaciones se distribuyen de manera no aleatoria en el espacio, mientras que el desbalance de clases se refiere al hecho de que existen muchos más registros de ausencia que registros de presencia. Ambos sesgos son comunes en datos colectados de manera no sistemática y pueden afectar nuestra capacidad para obtener inferencias confiables a partir de estos. Afortunadamente, estos sesgos pueden abordarse en gran medida mediante el submuestreo espacial de los datos antes del modelado (Robinson et al. 2018, Johnston et al. 2021). Para realizar un submuestreo adecuado, creamos una grilla de hexágonos con un espacio de 5 km entre los centros de hexágonos a lo largo del polígono establecido para el área de cría y de invernada del Piojito Trinador (ver métodos: set de datos de cría y de invernada). Luego en cada hexágono seleccionamos al azar un registro de ausencia y un registro de presencia para cada semana del año y para cada polígono. Luego del submuestreo espacial contamos con 502 presencias y 19 988 ausencias para el polígono de cría, y con 460 presencias y 10 014 ausencias para el polígono de invernada. Este proceso lo llevamos a cabo con el paquete de R dggridR (Barnes y Sahr 2017) y lo repetimos para cada corrida de los modelos a modo de asegurar que todas las observaciones tengan iguales chances de ser seleccionadas. Con el submuestreo espacial nos aseguramos la independencia espacial de registros de presencia y ausencia, evitando utilizar registros de cada clase que solapen espacialmente durante la creación de modelos. A su vez, obtuvimos un balance más equilibrado entre el número de registros de presencias y ausencias.
Los modelos de distribución de especies asocian los registros georreferenciados a predictores de hábitat y climáticos y así logran predecir las condiciones ambientales óptimas para encontrar la especie de interés (Guisan y Zimmerman 2000). Utilizamos una serie de variables que describen tipos de hábitat, topografía, y clima y las asociamos a los registros del Piojito Trina-dor para modelar su distribución estacional. Las variables de hábitat las obtuvimos de los productos de datos Copernicus (Buchhorn et al. 2020). Dado que los organismos interactúan con su entorno y no con un punto específico, caracterizamos el paisaje que rodea a cada registro en vez de simplemente extraer la clase de hábitat indicada para el sitio de cada registro. Para esto, calculamos el porcentaje de cada tipo de hábitat presente dentro de un buffer circular de 2,5 km de radio centrado en cada sitio con registros de presencia o ausencia del piojito. Debido a que las capas de hábitat de este producto de cobertura terrestre datan del año 2015 en adelante, asociamos los registros del 2010 al 2014 a las capas del año 2015. Las variables climáticas utilizadas fueron precipitación media mensual y temperatura media mensual, obtenidas de WorldClim v2.1 (Fick y Hijmans 2017). Asociamos a cada registro de presencia y ausencia a los valores de precipitación y temperatura correspondientes al mes del registro. Adicionalmente, utilizamos dos variables que no cambian en el tiempo: elevación (promedio y desvío estándar) obtenidas de Amatulli et al. (2018) y distancia euclidiana a ríos de la capa vectorial de ríos y lagos de los productos de Natural Earth versión 4.1.0 (Natural Earth 2020). El promedio y desvío estándar de la elevación fue computado dentro de una grilla de 2,5 x 2,5 km centrada en cada localidad. Estandarizamos la resolución de las variables a 2,5 minutos para que las capas coincidan utilizando el paquete de R ráster (Hijmans y van Etten 2016). La lista completa de variables utilizadas es proporcionada en los Materiales Suplementarios S2.
Utilizamos como covariables en los modelos la información de esfuerzo de muestreo disponible en cada lista de eBird. Estas covariables describen la heterogeneidad del muestreo, lo cual tiene efectos directos en la detectabilidad de la especie de estudio (Gorleri et al. 2021, Johnston et al. 2021). Incluimos las siguientes covariables: tipo de muestreo (estacionario o con desplazamiento), distancia recorrida, duración del muestreo, y número de participantes.
Seleccionamos y submuestreamos 20% de los registros de presencia y 20% de los registros de ausencia como set de testeo para la evaluación de cada modelo utilizando la validación cruzada. Para el entrenamiento de cada modelo eliminamos las observaciones que fueron al set de testeo. Repetimos el proceso de separar el set de datos en subconjuntos de entrenamiento y testeo 100 veces, creando 200 sets de datos únicos contra los cuales fueron evaluados nuestros modelos (100 para cada temporada). Utilizamos el paquete de R ranger (Wright y Ziegler 2017) para correr un modelo de bosque aleatorio balanceado (Balanced Random Forest) para la temporada de cría y de invernada. El bosque aleatorio balanceado es una modificación del algoritmo de bosque aleatorio diseñado para su uso con datos con exceso de ceros o ausencias (Chen et al. 2004). Creamos 1000 árboles de clasificación para cada tipo de modelo y fijamos en 4 el número de variables que el modelo podía seleccionar en cada división para cada árbol (James et al. 2013). Finalmente, calibramos los modelos siguiendo los métodos delineados en Johnston et al. (2021), a fin de evitar inflar los valores de prevalencia en las probabilidades de detección observadas luego de haber ejecutado el submuestreo espacial.
Evaluamos el rendimiento predictivo de los modelos de cría y de invernada mediante el área bajo la curva (AUC; Fielding y Bell 1997). Esta métrica evalúa la capacidad de las predicciones de probabilidades de cada modelo para discriminar entre sitios de ocurrencia y no ocurrencia, asumidos a partir de los registros de presencia y ausencia de la especie. Los valores de AUC varían de 0.5 (no hay capacidad de discriminación) a 1 (capacidad de discriminación perfecta), siendo aceptables valores mayores a 0.7 (Hosmer y Lemeshow 2000). Posteriormente realizamos una prueba t de Student de dos colas no pareado para determinar si existieron diferencias significativas en el rendimiento de los modelos de cría y de invernada.
Extrapolamos las predicciones resultantes de los modelos de cría y de invernada con mejor rendimiento predictivo al espacio geográfico para visualizar la distribución estimada del Piojito Trinador en cada temporada. Las probabilidades de detección extrapoladas al mapa fueron calculadas asumiendo un muestreo de un observador estándar con una duración de 1 hora, y recorriendo una distancia de 1 km. Realizamos la predicción al 1 de enero 2021 para el modelo de cría, y al 1 de julio 2021 para el modelo de invernada. Adicionalmente calculamos el ranking de importancia de predictores y valores de dependencia parcial que describen la relación de cada una de las variables explicativas con la variable respuesta (respuestas especie-hábitat). Para la creación de los gráficos de dependencia parcial utilizamos las 100 réplicas de los modelos para cada temporada, calculando la media e intervalos de confianza del 95% sobre los valores de dependencia parcial obtenidos para cada uno de los predictores. Los valores de dependencia parcial fueron calculados con las funciones por defecto del paquete R ranger.
Modelos de fenología
Examinamos la fenología o timing migratorio del Piojito Trinador estimando su abundancia relativa a lo largo de las semanas del año en 4 cuadrantes que delimitan las principales regiones donde puede registrarse a la especie: el sur del monte (esquina inferior: 44°28’S, 61°25’O; esquina superior: 35°54’S, 69°18’O), la región de sierras centrales de Argentina (34°12’S, 62°45’O; 29°22’S, 66°08’O), el litoral (35°28’S, 57°23’O; 28°33’S, 61°45’O) y la región chaqueña (28°12’S, 57°25’O; 21°21’S, 65°55’O). Con las predicciones resultantes calculamos la fecha estimada de arribo y partida, y la fecha de máxima abundancia (pico) para cada uno de los cuadrantes especificados. Para lograr las predicciones primero extrajimos de eBird (eBird Basic Dataset 2021, Material Suplementario S3) los registros de presencia (con información de cantidad de individuos observados) y ausencia del Piojito Trinador para cada uno de los cuadrantes, y asignamos a cada registro la semana del año correspondiente con el paquete de R lubridate (Grolemund y Wickham 2011). Luego replicamos el proceso de submuestreo espacial (ver métodos arriba) a modo de emparejar las ausencias con los registros de presencia.
Estimamos la abundancia relativa para cada cuadrante a lo largo de las semanas de año mediante la construcción de Modelos Aditivos Generalizados (binomial negativo), utilizando la cantidad de individuos reportados por semana del año como variable respuesta y las semanas del año como variable explicativa. Corrimos los modelos en el paquete de R mgcv (Wood 2017). A modo de contemplar diferencias en la detectabilidad debido a la variación en el esfuerzo de muestreo incluimos covariables de esfuerzo como tipo de muestreo (estacionario o con desplazamiento), distancia recorrida, duración del muestreo, y número de participantes. Para cada modelo, estimamos el parámetro de suavización utilizando 5 grados de libertad y ajustamos cada modelo con un spline de regresión cíclica (CCRS). En el CCRS, el inicio y el final del suavizador están restringidos para coincidir en valor; por lo tanto, estos son útiles para ajustar modelos con componentes cíclicos como los efectos estacionales (Pedersen et al. 2019). Determinamos las semanas estimadas de arribo y partida para cada cuadrante mediante el cálculo de los percentiles 5 y 95 de la suma acumulada de las predicciones de abundancia relativa del modelo de cada cuadrante, y también determinamos los picos de abundancia visualizando las semanas del año con mayores valores estimados.
Resultados
Los modelos de distribución mostraron un notable cambio en la distribución geográfica estimada para los períodos de cría y de invernada del Piojito Trinador, evidenciando el comportamiento migratorio de la especie (Figura 1). El modelo de cría mostró que la especie presenta altas probabilidades de ser detectada a lo largo del monte de llanuras y mesetas, el chaco árido y en el sur del espinal (Figura 1A). En contraste, el modelo de invernada señala una amplia distribución de la especie en las llanuras del chaco hasta el norte de Paraguay y sur de Bolivia, y hacia el litoral argentino, principalmente en el espinal del Santa Fe y suroeste de las provincias de Corrientes y Entre Ríos (Figura 1B). Se observa muy poco solapa-miento entre ambos modelos, el cual se ve limitado principalmente a la región peri-serrana de San Luís y Córdoba. El rendimiento predictivo de los modelos de distribución fue aceptable en términos de AUC (modelo de cría: AUC = 0.89 ± 0.01; modelo de invernada: AUC = 0.87 ± 0.01), siendo el modelo de cría superior (t-test = 7.42, P-valor < 0.001).
El arbustal resultó ser el hábitat con mayores chances para detectar al Piojito Trinador tanto en su período reproductivo como de invernada (Figuras 2 y 3). Esto se evidenció en las curvas de respuesta de especies-hábitats, donde se observó un incremento en la probabilidad de detección del piojito a mayor proporción de arbustal en el paisaje. A su vez, el arbustal resultó el predictor de hábitat que mayor peso otorgó a las predicciones de los modelos, es decir, clasificó por encima del resto de los hábitats en el ranking de importancia de predictores (Figura 2 y 3). Los modelos también predijeron que el piojito prefiere tierras bajas y relativamente secas en ambas temporadas, lo cual se reflejó en una respuesta negativa de las probabilidades de detección de la especie a los predicto-res de precipitación y de elevación, siendo la especie hallada en regiones con precipitaciones que varían entre 0 - 150 mm mensuales durante la temporada de cría (noviembre a febrero) o de invernada (mayo a agosto), y en áreas con elevaciones que varían de 0 -1 500 msnm (Figura 2 y 3). El modelo de cría también predijo una asociación positiva de la especie a áreas de cultivos (Figura 2), aunque dicho hábitat ranqueó noveno, por debajo del resto de los predictores más relevantes. A su vez, el modelo de invernada predijo que la especie está asociada a ríos, a humedales herbáceos y a sitios urbanizados durante la temporada no reproductiva (Figura 3).
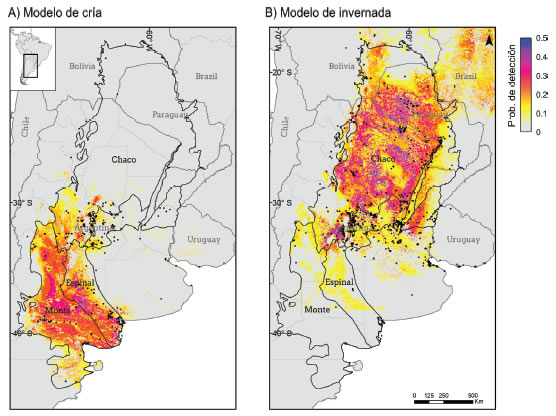
Figura 1: Modelos de bosque aleatorio balanceado que reflejan la probabilidad de detección del Piojito Tinador (Serpophagagriseicapilla) en el espacio geográfico durante la temporada de (A) cría y (B) invernada. Las probabilidades de detección fueron calculadas asumiendo un muestreo de un observador estándar con una duración de 1 hora y recorriendo una distancia de 1 km. Realizamos la predicción al 1 de enero 2021 para el modelo de cría, y al 1 de julio 2021 para el modelo de invernada. La resolución de los mapas es de 2.5-min. Los puntos negros indican los registros utilizados para entrenar cada modelo.
Los modelos fenológicos hechos con datos de abundancia describieron el timing migratorio del Piojito Trinador a lo largo de su distribución. En el cuadrante del monte, el Piojito Trinador exhibió su máxima abundancia en la semana 47 (mediados de noviembre), y sus fechas de arribo y partida de la región reproductiva resultaron la semana 34 (mediados a finales de agosto) y la semana 14 (inicios de abril), respectivamente (Figura 4). En el cuadrante de la región central (que incluye mayormente a Córdoba y San Luís) la curva fenológica presentó una forma sinusoidal con dos picos de abundancia: el primero en la semana 16 (mediados de abril) y luego en la semana 37 (mediados de septiembre), coincidente con fechas esperables para los pasajes migratorios de otoño y de primavera, respectivamente (Figura 4). En la región litoral y el chaco la especie no exhibió picos de abundancia, más bien, el modelo adoptó una forma parabólica negativa con el máximo estimado en la semana 30 (fines de julio) para el chaco, y en la semana 27 (inicios de julio) para el litoral. En la región del chaco las fechas estimadas de arribo y partida fueron las semanas 19 (inicios de mayo) y 39 (inicios de octubre). Para la región litoral las fechas estimadas de arribo y partida fueron las semanas 13 (fines de marzo) y 40 (inicios de octubre) (Figura 4).
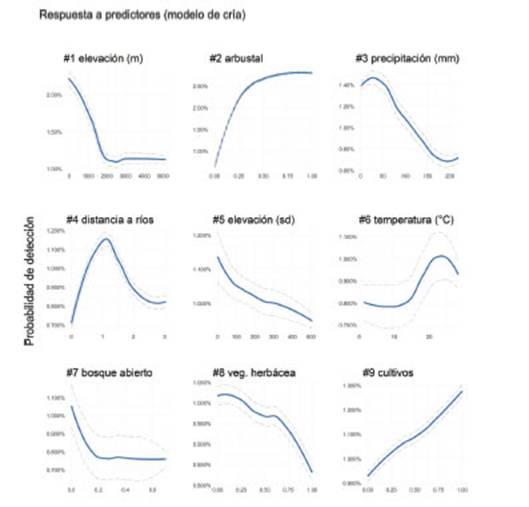
Figura 2: La dependencia parcial de la probabilidad de detección del Piojito Trinador (Serpophagagriseicapilla) para los predictores climáticos y ambientales más relevantes (#1 al #9) identificados por el modelo para la temporada de cría (ver Figura 1). La línea azul indica el promedio y la línea con guiones los intervalos de confianza del 95% de los valores de dependencia parcial obtenidos luego de 100 réplicas de los modelos. Suavizamos las curvas con una función de loess.

Figura 3: La dependencia parcial de la probabilidad de detección del Piojito Trinador (Serpophaga griseicapilla) para los predictores climáticos y ambientales más relevantes (#1 al #9) identificados por el modelo para la temporada de invernada (ver Figura 1). La línea azul indica el promedio y la línea con guiones los intervalos de confianza del 95% de los valores de dependencia parcial obtenidos luego de 100 réplicas de los modelos. Suavizamos las curvas con una función de loess.
Figura 4: Modelos aditivos generalizados que reflejan la abundancia relativa del Piojito Trinador (Serpophaga griseicapiüa) a lo largo de las semanas del año en los cuadrantes (A) Chaco, (B) Litoral, (C) Sierras Centrales, y (D) Monte. Las barras verticales indican la semana estimada de arribo y partida de la especie para cada modelo y las estrellas indican picos de abundancia para cada modelo. Rojo: registros en época reproductiva (noviembre a febrero), amarillo: registros en época de migración (marzo a abril y septiembre a octubre), azul: registros en época de invernada (mayo a agosto).
Discusión
Describimos con precisión la migración del Piojito Trinador mediante la utilización de un set de datos estandarizado y de alta calidad de eBird. Señalamos al monte austral como la principal región reproductiva de la especie, siendo el litoral argentino y el Gran Chaco sus áreas de invernada habituales. Encontramos que la especie tiende a frecuentar arbustales en regiones con climas áridos o semi áridos, aunque también se asocia a habitats arbustivos cercanos a cursos de agua durante el invierno. Además, evidenciamos una potencial ruta migratoria de la especie que involucra al chaco árido y sierras de Córdoba y San Luis, con picos de migración que ocurren a mediados de abril (pasaje migratorio de otoño) y mediados de septiembre (pasaje migratorio de primavera).
Distribución estacional
Nuestros modelos sugieren que el Piojito Trina-dor se reproduce en el monte, chaco árido y sur del espinal y migra hacia el Gran Chaco y norte del espinal para pasar el invierno austral. Este patrón general de migración coincide en gran medida con lo reproducido en Pearman & Areta (2020). También se refleja en varias menciones sobre la presencia estival del Piojito Trinador en el monte y chaco árido durante la primavera-verano (Cueto et al. 2008, Bodrati y Ferrari 2014) con evidencia de su reproducción (Salvador et al. 2017), y en la región chaqueña y del espinal norte durante el otoño-invierno (Zyskowski et al. 2003, Di Giacomo 2005, Gorleri et al. 2011, Gorleri 2017, Fandiño y Giraudo 2010). En contraste, no hallamos evidencia de que la especie realice movimientos migratorios entre el chaco-espinal-cerrado, como lo sugieren Capllonch (2018). Aunque el modelo de invernada predijo la presencia de la especie para el cerrado en Brasil, consideramos que esto se debe más bien a una sobre-predicción del modelo como resultado de los pocos registros de ausencia disponibles en dicha región para su entrenamiento (ver Material Suplementario S1).
La ecorregión del monte de llanuras y mesetas delimitó el área núcleo de reproducción del Piojito Tri-nador, reforzando la hipótesis de que esta especie es endémica de cría de Argentina (ver Pearman & Areta 2020). Aunque se ha mencionado al Piojito Trinador como posible nidificante en Bolivia (Herzog et al. 2016) y Paraguay (Lesterhuis et al. 2018), la evidencia de que esto suceda sólo es respaldada por una serie de registros estivales de la especie para tales países. Estos registros podrían tratarse de migrantes tardíos o individuos que por múltiples motivos hayan permanecido durante el verano en su área de invernada, no necesariamente reflejando su potencial reproducción. A pesar de que tomamos un enfoque poco conservador para la construcción del modelo de cría (es decir, no excluimos a registros extralimitales), igualmente el modelo indicó nulas probabilidades de detección de la especie en el norte de Argentina, Bolivia y Paraguay, apoyando la idea de que los registros actuales resultan de observaciones aisladas. Cabe mencionar también que existe la posibilidad de confusión del Piojito Trinador con otras especies de piojitos similares que abundan en verano en la región chaqueña (por ejemplo, con Inezia inornata). Por lo tanto, vemos necesario documentar la presencia del Piojito Trinador mediante registros auditivos durante la temporada estival. Esto permitirá lograr un mejor entendimiento de su distribución estival fuera de las áreas esperadas de reproducción teniendo evidencia concreta.
Por otro lado, aunque existen registros que documentan al Piojito Trinador en Uruguay y el sur de Brasil (estados de Santa Catarina y Rio Grande do Sul) durante otoño-invierno (Bencke et al. 2002, Bencke 2010, Bittencourt de Farias 2016, Fjeldsá 2020), los modelos de invernada no encontraron áreas potencialmente aptas para encontrar a la especie en dichas regiones. No obstante, esto es esperable dado que los registros resultan de observaciones aisladas y el Piojito Trinador es considerado un visitante irregular en tales países (Fjeldsá 2020). El modelo de invernada, por lo tanto, refleja las bajas (o nulas) chances de localizar un Piojito Trinador en Uruguay y el sur de Brasil asumiendo un muestreo de un observador estándar con una duración de 1 hora y recorriendo una distancia de 1 km.
Hábitats y condiciones climáticas de preferencia
Encontramos que el Piojito Trinador se asoció a hábitats arbustivos en regiones climáticas relativamente áridas o semi-áridas en ambas temporadas. Reportes locales también mencionan que la especie prefiere arbustales o bordes de bosque durante la temporada reproductiva (ver Cueto et al. 2008, Salvador 2017) y no reproductiva (Di Giacomo 2005, Gorle-ri et al. 2011, Gorleri 2017). No obstante, cabe aclarar que las respuestas a hábitats señaladas en este estudio deben ser interpretadas a escalas regionales y no a escalas locales dada la resolución de 5 km de radio de las capas de cobertura terrestre que utilizamos (ver también Bauder et al. 2021).
Aunque la especie no parece cambiar abruptamente de preferencias climáticas y ecológicas de una temporada a otra, observamos que existe una mayor afinidad a sitios cercanos a ríos y humedales durante la temporada no reproductiva. En línea con este hallazgo, Gorleri (2017) menciona que el Piojito Trinador es una de las especies más abundantes en arbustales y bosques asociados a humedales durante el invierno en el centro-oeste de Formosa, Argentina (ver también López-Lanús y Mencia 2017). Dicha tendencia también es observada en otras especies de tiránidos migratorios que crían en bosques xerófilos de Sudamérica y que invernan en ambientes más húmedos o asociados al agua, por ejemplo, en el Piojito Picudo Inezia inornata (Gorleri et al. 2021) o Suirirí Pico Corto Sublegatus modestus (Lees et al. 2013).
Por otro lado, nuestros modelos encontraron como hábitats favorables para el Piojito Trinador a las áreas de cultivo (temporada de cría) y urbanizadas (temporada de invernada). No obstante, ambos resultaron ser los predictores con menor importancia relativa entre los predictores más relevantes identificados por los modelos. Si bien existen varios registros del Piojito Trinador en ambientes antropizados, no consideramos a estos ambientes como potencialmente favorables para la especie en base a nuestra experiencia de campo. Es posible que la asociación positiva hallada por los modelos a las áreas urbanas y de cultivos sea producto del sesgo espacial de los datos. Los datos de ciencia ciudadana suelen estar mayormente distribuidos en cercanías a ambientes urbanos o en áreas rurales (Tiago et al. 2017) lo cual dificulta la capacidad de los modelos de distribución de realizar una correcta lectura de la injerencia de estos ambientes en la distribución de especies (Boakes et al., 2010, Johnston et al. 2020). A pesar de haber mitigado el sesgo espacial en nuestros modelos mediante el uso de ausencias y a través del submuestreo espacial (ver métodos), creemos que los efectos del sesgo se vieron igualmente reflejados en los modelos al predecir como favorables a las áreas urbanas y de cultivos.
Fenología migratoria
Las curvas fenológicas sugieren que el Piojito Tri-nador parte desde el monte a inicios de abril a través de la región de sierras centrales de Argentina (Córdoba y San Luis), y que permanece durante gran parte del otoño-invierno en el litoral argentino y el chaco, para luego retornar posiblemente mediante la misma ruta. Esta potencial ruta de migración se evidenció por los dos picos de abundancia observados en el cuadrante de la región de sierras centrales, ambos coincidentes con las fechas esperables de pasajes migratorios de otoño y primavera de la especie (mediados de abril y mediados de septiembre). Otras especies de tiránidos también utilizan la región central de Argentina como ruta migratoria, como el Fiofío Silbón (Elae-nia albiceps chilensis, Bravo et al. 2017). A pesar de que no podemos descartar que el Piojito Trinador utilice otros corredores migratorios, argumentamos que el uso del corredor central cobra sentido para aquellas especies arborícolas, ya que pueden favorecerse de la continuidad de bosque que comunica el monte con la planicie chaqueña.
En el cuadrante del chaco y de la región litoral no encontramos evidencia de pasaje migratorio. Coincidente con nuestros modelos de distribución, las formas parabólicas de las curvas fenológicas señalan a estas regiones como los sitios establecidos de invernada para la especie. La fenología descripta en nuestro estudio para el chaco (permanencia de la especie desde inicios de mayo a inicios de octubre) coincide en términos generales con lo expuesto en Di Giacomo (2005) para el este de Formosa, quien señala al 30 de abril y 1 de octubre como fechas extremas de arribo y partida del Piojito Trinador. Por otro lado, las diferencias que observamos en las fechas de arribo para la región del litoral (finales de marzo) y del chaco (inicios de mayo) podrían explicarse debido a una mayor proximidad geográfica de la primera al área núcleo de reproducción del piojito, con lo cual se esperaría un arribo más temprano de la especie. También es plausible que ocurran movimientos poblacionales entre sitios de invernada, con un primer arribo de la especie a la región litoral y posterior movimiento al chaco. Este fenómeno, que involucra cambios entre ecorregiones durante el período de invernada, está documentado mediante el uso de geolocalizadores en otras especies de tiránidos como Tyrannus savana y Elaenia albiceps (Jahn et al. 2016, Bravo et al. 2017). Por otro lado, que las fechas de partida de la migración primaveral hayan coincidido entre el chaco y litoral (inicios de octubre) es congruente con la hipótesis de una migración primaveral rápido y mejor sincronizado (en relación a la migración en otoño) por parte de las especies migratorias (Nilsson et al. 2013).
El Piojito Trinador logra su pico máximo de abundancia en la región sur del monte a mediados de noviembre (plena época reproductiva), reiniciando su ciclo anual. No obstante, tuvimos un muy bajo número de muestras disponibles para el entrenamiento del modelo creado para el cuadrante del monte (n=282), con lo cual los resultados podrían no ser del todo fiables. En particular, creemos que la fecha de arribo estimada para el sur del monte (mediados de agosto) resulta muy temprana para la especie. Indudablemente, un incremento en el número de muestras proporcionará una mejor calibración del modelo fenológico. Dicho esto, y debido al importante el rol ecológico que juega el monte para muchas especies migratorias (Cueto et al. 2008), resaltamos la necesidad de dirigir mayores esfuerzos de muestreo hacia esta ecorregión durante la temporada estival en plataformas de ciencia ciudadana. Esto permitirá incrementar la cantidad y calidad existente de datos disponibles, y mejorará el poder predictivo de los análisis ecológicos que puedan desprenderse de los mismos.
Conclusión
Brindamos nueva evidencia sobre la enorme utilidad de los datos de ciencia ciudadana para la reconstrucción de patrones migratorios y fenológicos en aves en resoluciones más detalladas que previamente descriptas. A su vez, nuestros resultados reflejan la importancia de los datos que proporcionan información del esfuerzo de muestreo para controlar sesgos inherentes a los procesos no sistemáticos de recolección de datos, hoy disponibles en datos colectados a través de eBird en nuestro continente. El incremento en la calidad y cantidad de registros observacionales en bases de datos de ciencia ciudadana será de gran ayuda para descubrir y profundizar nuestro conocimiento de los patrones migratorios de las aves del
Neotrópico, un campo de estudio que aún requiere de estudios más detallados.
Recibido: 16 de Enero de 2022
Aceptado: 3 de Mayo de 2022

