











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
There are few complete sponges from the upper - Ordovician to early Silurian rocks all over the world and no records of sponge bodies have been found yet in the Precordillera, western Gondwana. However, sponge spicules have sparse records in the Central and Eastern Precordillera of the San Juan Province (Figure 1a). Two hexactinellid spicule assemblages were previously recorded from the Precordilleran siliciclastic platforms during the post-glacial Hirnantian event. The first spicules discovery (Beresi, 2013) is from the Early Silurian (Rhuddanian) of the La Chilca Formation (Cuerda, 1966) exposed at Cerro El Fuerte sec-tion (Figure 1b), about 20 km east of Jáchal city Cendal Precordillera, San Juan Province. A few silicificated hexactinellid spicules including four pentactins and two hexactins were recuperated from a fine sandstone with calcareous cement by acid treatment (Beresi, 2013; Figure 2a - d and 2e - f). The second record is a post-glacial Hirnantian hexactinellid spicules from the Don Braulio Formation (Baldis et al., 1982), at the homonymous section, on the eastern flank of the Sierra de Villi-cum (Figura 1b), Eastern Precordillera of the San Juan Province (Jiménez - Sánchez et al., 2014). This association consists of few spicules together with fragments of ramose cylindrical cryptostomatid bryozoans and one crinoid plate obtain by acid treatment.
The new record of spicules described here, was also collected from the Don Braulio Formation, at the homonymous stratigraphic section, in the Sierra de Villicum. The fauna recuperated from ochre fine - grained sandstone lenses consists of a few hexactines or derivatives (triaxon with five or six rays) and one stauractin (four rays). Although very limited material, the spicules are of interest because they represent a new finding of hexactine-llid and possibly reticulosan sponge communities components of the benthic fauna of cool-water in shallow siliciclastic environments.

Figure 1 A) Map of the Province of San Juan with the location of the geologic provinces. The Argentine Precordillera is in the eastern foothills of the Andean mountain chain. Boxed area enlarged in B) showing the positions of the Cerro El Fuerte, Central Precordillera, and the Sierra de Villicum, Eastern Precordillera of the San Juan Province. Fossil localities indicated by star symbol. / Figura 1. A) Mapa de la Provincia de San Juan con la ubicación de las provincias geológicas. La Precordillera Argentina se encuentra en las estribaciones orientales de la Cordillera de los Andes. El recuadro ampliado en B) muestra las posiciones del Cerro El Fuerte, Precordillera Central, y la Sierra de Vilicum, Precordillera Oriental de la Provincia de San Juan y las localidades fosiliferas indicadas por las estrellas.
This paper describes these sponge spicules, their paleoenvironments and their signifi-cance after the end Ordovician post - extinc-tion crisis (LOME) in the Precordillera basin, localized at the high latitudes in the western margin of the Gondwana paleocontinent.
Materials and methods
The available material occurs on the surface of five lenticular fine-grained sands-tones with calcareous cement, concentrated in the upper mudstone interval of Don Braulio Formation. They consist of six isolated hexactins or derivatives with broken tips of rays and one stauractin. They are recrystalli-zed; thus, they cannot be isolated from the rock by acid treatment. Intensive sampling was conducted at layers and several lenses of the mudstone interval, without producti-ve sponges or spicules. Macroscopic samples were examined with a binocular microscope to identify the spicules. No other fossils were observed. Spicules were photographed dry, using a digital camera Olympus attached to an Olympus binocular stereo microscope mo-del SZ61; observations were carried out using the Olympus microscope. The contrast, color and brightness of images were adjusted using Core Photo - Paint x7.
All material is stored in the Paleo - Inver-tebrates Repository of the Argentine Institute of Nivology, Glaciology and Environmental Scien-ces (IANIGLA), CONICET at Mendoza Pro-vince, under the collection numbers and the code IANIGLA - PI: 3383, 3384, 3385, 3386, 3387.
Geological settingThe Geological Province of Argentine Precordillera is a fold - and thrust belt involving Palaeozoic and Tertiary sediments. It has been considered mainly two models of the origin of Precordillera: a) as a rifted drifted microcontinent, originally located in Laurentia and accreted to the Gondwana margin (Benedetto, 2004; Ramos, 2004), and b) as a terrain located on the southern margin of Western Gondwana that would have migrated to its current position by transcurrence (Baldis et al, 1989; Aceñolaza et al., 1999, 2002; Finney et al, 2003). The Precordillera has classica-lly been divided into three morpho - structural do-mains: Western (Baldis et al, 1984), Central (Baldis and Chebli, 1964; Baldis et al, 1984) and Eastern (Ortiz and Zambrano, 1981).
In the Eastern Precordillera of San Juan, the Don Braulio Formation (Baldis et al, 1982) represents the top of the quasi almost - complete Ordovician sequence at the Villicum Range. The formation consists of a mixed clastic - carbonate succession which crops out on the eastern flank of the Villicum Range. Its typical section, the Don Braulio Creek, is located on the geogra-phic coordinates of 31°12’50 53” S latitude and 68°29’8.73” W longitude. In this section the formation reaches 40 m thick (Figure 2).
The Hirnantian - Llandovery Don Braulio Formation (DBF) overlain by erosive contact the La Cantera Formation (Sandbian) in the Don Braulio Creek, as well as the La Pola Formation (Katian-Hirnantian) in the La Pola Creek, and the top is capped by olistostrome deposits of the Late Silurian La Rinconada Formation. The formation was divided in four members by Peralta (1993): the lower Diamictite Member, 15 to 20 m thick; the Mudstone and Sandstone Member,10 to 12 m thick, composed of greeni-sh grey mudstones and fine-medium grained tabular sandstones including ochre calcareous and marly fossiliferous lenses bearing trilobites, bra-chiopods, bryozoans, crinoids, bivalves and spicules, among others; the Ocher Member, 7-12 m thick, is composed of monotonous and massive yellowish red bioturbated mudstone layers and fine-grained siltstone containing Skolitus; and at the top, the Ferriferous Upper Member, 10 m thick, mainly composed of oolitic ironsto-nes, fine-grained sandstones with interbedded shales bearing graptolites from the Atavograptus atavus Biozone (Peralta, 1985) which caps the sequence (Figure 2).
The Hirnantian age of the Don Braulio Formation at the type Don Braulio Creek section, was formerly provided by the Dalma-nitina-Eohomalonothus fauna (Baldis and Blasco, 1975) and the Hirnantia fauna (Benedetto, 1986). The Hirnantia fauna contains typical brachiopods as Hirnantia sagittifera (M’Coy 1851), Dalmanella aff. D. testudinaria (Dalman 1828), Paromalomenapolonica (Temple 1965) and Eostro-pheodonta hirnantiensis (M’Coy 1851) (Benedetto 1986, 1990) together to bivalves (Modiolospis), bryozoans and trilobites (Mucronaspis). This fauna was collected from ochre calcareous lenses of the base of the Fossiliferous Mudstones and Sandstones Member immediately above the glacial diamictites (Figure 2). Thus, this fauna is the oldest preserved in the Don Braulio Formation and corresponds to the cool-water shelly fauna linked to the post-Hirnantian glaciation in Precordillera localized on the western margin of Gondwana at middle and high and latitudes (30°- 45° L.S, Torsvik and Cocks, 2009).
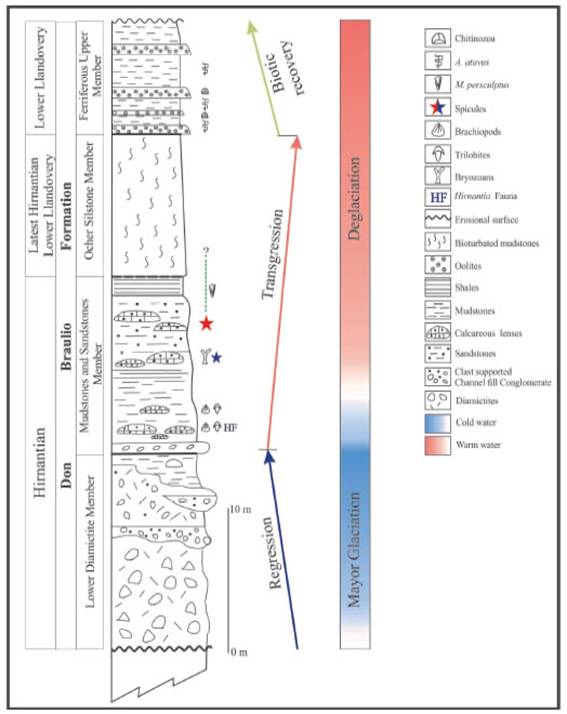
Figure 2: Stratigraphic section of the Don Braulio Formation with lithostratigraphic members exposed at the homonymous section, in the Villicúm Range, Eastern Precordillera. The stars indicate the collected Hirnantian spicules location. Blue star location by Jiménez-Sánchez et al, 2014; red star, this paper. / Figura 2. Sección estratigráfica de la Formación Don Braulio y sus miembros, expuesta en la sección homónima de la Sierra de ViMcum. Las estrellas indican la ubicación de recolección de las espículas del Hirnantiano.
In the same member, above the Hirnantian fauna, a second fossiliferous assemblage consists of dispersed valves of Hindella crassa (J. de C. Sowerby), Eostropheodonta aff. E. hirnantensis and Plectothyrella sp. (Benedetto, 1986) among others, together with Eohomalonotus villicunensis (Baldis and Blasco, 1975) and conulariids without bra-chiopods of the typical Hirnantia fauna. At the upper part, in the mudstone interval, Peralta and Baldis (1990) recorded the presence of a mono-typic graptolite assemblage Metabolograptus pers-culptus (Hirnantian, Late Ordovician, ~444Ma). There, the macrofauna decrease but, isolated sponge spicules occur on the surface of scarce, carbonate-sandstone lenses, probably, during the occurrence of the M. persculptus. This member is succeeded by the bioturbated ochre Siltstones Member with Skolitus and without shelly fauna.
The Llandovery age of the upper part of the Don Braulio Formation was provided by Volkhei-mer et al (1980) on the basis of the palynomorphs, mainly chitinozoan records and graptolites indica-ting the Atavograptus atavus Zone (Peralta, 1985).
Paleoclimatic condition: Gondwana Hirnantian glaciation eventThe extensive Gondwanan glaciation near the Katian/Hirnantian boundary (latest Ordo-vician) was the first major Phanerozoic glacia-tion, centered in northwestern Africa (Ghienne, 2003). This Ice Age (445 - 443 Ma; Sheehan, 2001) was the most spectacular, short-term, and cooling event of the Phanerozoic, followed by equally rapid warming, with changes in the ocean's chemistry and circulation (Botting et al, 2018, Scotesse et al, 2021). The end - Ordovician mass extinction (LOME) was the first of the “Big Five” extinctions of the Phanerozoic (Raup and Sepkoski, 1982). This event was among the largest known and had detrimental effects on the marine community, with 85% species loss on all platforms of Gondwana (e.g. Sheehan, 2001; Harper et al., 2014). During the Latest Ordovician ice sheets covered ~50% of Gondwana and extended to latitudes of ~35 S. The Gondwana landmass was located south of the equator at 35° to 45° LS (Torsvik and Cocks 2009, 2013). The Eastern-derived glacial sedi-ments were deposited along the eastern margin of Precordillera affecting the Don Braulio and other formations of the basin (Peralta and Car-ter, 1990).
The Hirnantian ice house and mass extinc-tion could have been triggered by an unknown bolide impact named the “Khione” Impact event (Brenchley and Marshall, 1999; Brenchley and Newall, 1984; Sheehan, 2001) which occurred during a relatively high global temperatures main 20° - 22° C (Scotesse et al, 2021).
The late-Ordovician mass extinction event (between 445.6 Ma and 443.7 Ma) has been interpreted as consisting of two pulses based on the fossil records, corresponding to the start and end of the Hirnantian glaciation (Harper et al, 2014). The first is associated with the onset of glaciation and the second was prompted by the melting of the Gondwanan ice sheet, which led to glacio - eustatic rise (Shee-han, 2001) and extensive black shale deposition (Delabroye and Vecoli, 2010). However, Wang et al. (2019) have suggested that the LOME comprises a rapid single - pulse of mass extinction, followed by a prolonged and intermittently initial recovery due to climatic changes through the Himantian, before the start of a progressive re-establishment of marine ecosystems during the early - Silurian due to better weather condi-tions. The post-extinction interval is reflected by a mudstone fossiliferous interval marked by the globally recognized Hirnantia fauna ecosys-tem, sea - level changes, and climate together with an oceanic turnover, occurred across the Ordovician - Silurian interval at a global level (Chen et al, 2005).
The Hirnantian ice house was followed by the introduction of a distinctive and cosmopoli-te Hirnantian brachiopod fauna sensu Rong and Harper (1988), which expanded globally due to a rise in global sea level following deglaciation (Harper etal, 2014). This biota is a typical cool/ cold - water, related to onshore, shallow water. It has the widest geographically distribution in the Phanerozoic and has been documented from many plates or terranes throughout the world (Rong and Harper, 1999; Rong etal, 2018, 2020) including the Precordillera.
Sponge spiculesSponges (Phylum Porifera) are a group of sessile suspension - feeding metazoans, cha-racterized by a body built around a system of water canals and chambers. Most sponges pos-sess a skeleton of mineral spicules (calcareous and siliceous), spongin fibres and spiculoids or a combination of both (Bergquist, 1978). In general terms, due to the fragility and the che-mical composition of their skeletons, sponges are preserved partly or fully articulated, but of-ten only dissociated spicules with poor preser-vation remain in the sediments released by the decay of the soft tissue. Nevertheless, sponge spicules are not uncommon in many deposits. Spicules produced by sponges are highly diverse in shape and size and assemblages of isolated spicules can indicate morphological diversity of sponges. However, spicules have often been neglected because of their minute size and the difficulty of assigning them to specific sponge taxa.
Spicules are separated conventionally into two categories, mega - and microscleres, accor-ding to their size and their role in the skeleton (e.g. Uriz etal, 2003). Megascleres are usually of greater size than microscleres, and form the main skeletal framework.
The observed spicule assemblage of the Don Braulio Formation comprises megascleres, dominantly classical hexactins (six rays) including possibly the presence of pentactines (five rays) and one stauractin with four rays. They are poorly preserved with some end rays broken so, it is impossible to determine their total length. Hexactins are different in size and superficially, some sharing the same overall form. The ex-ternal information is lost as the traces of axial canals. Spicules are recrystallized to clear calcite after presumed originally silica composition.
Class HEXACTINELLIDA Schmidt, 1870 Order and Family UNKNOWNFigure 3a, b, d, e.
Material: IANIGLA-PI 3383, 3384, 3385, 3386
Hexactin - based spicules: five spicules with six-pointed rays. Spicules are commonly incompletes with five distal rays and the proximal ray reduced to a knob (indicated by arrows) and of-ten slightly inflated.
Ray diameters of 0.11 - 0.28 mm and ray preserved lengths range from 0.57 - 1.40 mm; the tips of some rays are generally broken. No exter-nal features have been preserved.
“Reticulosa”
Order and Family uncertain Figure 3c
Material: IANIGLA-PI 3386
One stauractin spicule with four rays in orthogonal arrangement (forming an angle of 90°). Ray diameter 0.12-0.14 mm and preserved ray lengths of 1.14-1.17 mm.

Figure 3: Himantian hexactinellid sponge spicules from the Mudstones and Sandstones Member of the Don Braulio For-mation, at the Villicum Range, Eastern Precordillera. (a, b, d, e) Hexactine-based spicules with broken tips of rays and proxi-mal rays aborted (indicated by arrows) in the central discs. c) Stauractin with four rays in orthogonal arrangement. / Figura 3. Espíenlas de esponjas hexactinélidas y probable reticulosa hirnantianas del Miembro Mudstones y Sandstones de la Formación Don Braulio, en la Sierra de Villicum, Precordillera Oriental; (a, b, d, e) Espíenlas hexactinas con los extremos de los radios rotos y los rayos proximales abortados indicados por flechas, en los discos centrales; c) Estauractina con cuatro rayos en disposición ortogonal.
Discussion
Significance of Hirnantian sponges in global terms
The late-Ordovician mass extinction (LOME), 445 million years ago, resulted in 85% of species dying out. However, hexactinellid deep - water siliceous sponges may have recove-red after some mass extinction due to that they have flourished and their diversity has remained high after severe glacial events (Muir et al, 2013, 2017).
In global terms, sponge communities oc-cur from the immediate aftermath of the Ordo-vician mass extinction. So far, sponges have been reported from the late Ordovician in China, par-ticularly in South China.
Hirnantian sponge faunas from China, ex-perienced a strong increase in diversity with more than 75 species that occupied empty eco - spaces in the deeper - water, somewhat anoxic conditions. These faunas show variations of sponges of mul-tiple lineages that survived the Late Ordovician mass extinction (Botting et al, 2017, 2018; Li et al, 2019; Wang et al, 2018, 2019). For example, well - preserved siliceous sponge assemblage was dis-covered from the Kaochiapien Formation (Upper Ordovician - Lower Silurian) in southern Anhui Province of South China, after the end-Ordovician mass extinction (Li et al, 2015). Also, in the same black siliceous mudstone formation an articulated rossellid sponge was recovered from the lower part of the M. perscuptus Biozone, latest Hirnantian and the lower part of the Akidograptus ascensus Biozone (early Rhuddanian), extending across the Ordovician-Silurian boundary (Li et al, 2019). Diverse and abundant sponges with preservation of soft tissues occur in the narrow mudstone interval, from the
Hirnantian Wenchang Formation in Anhui Coun-ty, South China , including reticulosans, protomo-naxonids, hexactinellids, and demosponges which lived during the post-extinction crisis interval (Bo-tting etal, 2017). Also, a wide range of major taxo-nomic groups are represented in the Anji sponge faunas as in other sections with Upper Ordovician - Lower Silurian deposits of South China. These assemblages developed in deep water, low energy ecosystem during the post-extinction Hirnantian event and before significant benthic recovery of these faunas in the Rhuddanian (earliest Silurian) when community types were re - established.
Sponges also flourished after other mass extinctions and may have facilitated ecosystem recovery by stabilizing sediment of at least deeper - shelf ecosystems (Botting et al, 2017;Wang et al., 2019).
The probable affinity of the spicules to sponges groups
The dominant spicule-type (hexactin) of the collection, is very common within most groups of the hexactinellid sponges and, consequently, they cannot be attributed to any specific hexactinellid taxon. Disarticulated hexactin based - spicules are typical of many groups of Ordovician sponges. Hexactinellid sponges are an ultra - conservative group, with the body plans of the main lineages possibly present from the early Palaeozoic onwards (Mehl, 1992).
In the collection, five spicules have six or five straight rays showing some different mor-phology and size and only one has four rays. Thus, probably they belong to different taxo-nomic hexactin - bearing groups representing more than one hexactinellid lineage.
The stauractin spicule (four rays) possibly belongs to the reticulosans, a complex group of hexactinellid - like sponges with a thin wall of hexactins and derivatives. Reticulosan sponges are probable the stem- group of siliceans and hexactinellid sponges (Botting and Muir, 2013). Thus, the Hirnantian spicule assemblage from the Precordillera belongs to the hexactinellids and probably to reticulosan sponges.
Taleoenvironment and age of sponge spicules
The spicules occur on the surface of fine sandstone lenses included in the upper part of the transgressive muddy platform of the Don Braulio Formation at the type section of the Vi-llicum Range. Therefore, they flourished in sha-llow siliciclastic environments. Due to the scar-city of fragmented, disarticulated and dispersed occurrence of these spicules can be considered a reworked assemblage. This leads to the inter-pretation that these sponges lived or survived during and after the post - glacial sea - level rise, the transgression due to the deglaciation, after the Hirnantian mass extinction (LOME). It is probable that during the transgression sponges evolved and flourished in shallow-water condi-tions.
According to the stratigraphic position, the spicules are younger than the cool Hirnantian fauna. Up to now, the record of the latest Hirnantian graptolite M. persculptus (Peralta and Baldis, 1990) is documented in the upper muds-tone above the spicule - bearing lenses. Spicules probably occurred in the middle to the late interval of M. persculptus as in the late Hirnantian of China. The proliferation of Chinese sponges from the post - glacial Hirnantian event possibly was likely to represent a global signal according to the interpretation by Botting et al. (2018). Also, this proliferation of sponge could be due to the global warming, and sea - level rise with global anoxia (Rong et al, 2020). On the other hand, Li et al (2015), have suggested that the emergence of sponges in the marginal platform, shallower habitats, may be related to the expansion of the euxinic and anoxic condition during the transgression post - extinction event.
Despite the differences in depositional settings of the Chinese sponges (deep water) and the DBF spicule assemblage (shallow water), their occurrence indicates that hexactine-llids and probably reticulosan communities can represent a global flourishing event (according to Botting et al, 2018) after the Ordovician mass extinction event.
Conclusión
Himantian spicule assemblage of the Precordillera is a rare finding. They represent one of the few examples of sponges inhabiting cold water in shallow siliciclastic environments at high paleolatitudes of peri - Gondwana. The sponges probably flourished during the middle to the upper interval of the Metabolograptus persculptus Biozone.
Spicules with different morphologies indi-cate that these elements could belong to diverse taxa of hexactinellids and probably of reticulo-san sponges component of the shallow sea floor community of the Eastern Precordillera during the latest Ordovician.
The spicule assemblage allows us a better insight into the impacts of the mass extinction event and the post-transgression on hexactinellid and reticulosan sponges of the Precordillera and, adds valuable information about their diversity and paleogeographic reconstructions during a time of critical climatic and biotic disruptions in the paleocontinent of Gondwana.
Acknowledgments
We dedicate this paper to the memory Dr. Bruno A. Baldis with whom M. Beresi made many field trips to the studied area and collected numerous fossils. This work has been supported by found provided by Consejo Nacional de Investigaciones Científicas y Tecnológicas (CO-NICET) from Argentina. We are grateful to J.P Botting for constructive comments on this paper’s previous version. We also thank I.J.C. Gavriloff and an anonymous reviewer. This paper is a contribution to the IGCP Project 735 “Rocks and the Rise of Ordovician Life-Filling knowledge gaps in the Early Palaeozoic Biodiversification”.
Recibido : 29 de setiembre de 2022
Aceptado : 17 de abril de 2023

