










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRIA. Revista de investigaciones agropecuarias
versión On-line ISSN 1669-2314
RIA. Rev. investig. agropecu. vol.44 no.2 Ciudad Autónoma de Buenos Aires ago. 2018
ARTÍCULOS
Efecto del rolado de baja intensidad (RBI) sobre la distribución radical de pastos y arbustos en un bosque del Chaco occidental
Ledesma, R.1; Elías, A.2; Kunst, C.1; Navarrete, V.1; Godoy, J.1
1 Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA) Santiago del Estero, Jujuy 850, Santiago del Estero, G4200CQR, Argentina. Correos electrónicos: ledesma.roxana@inta.gob.ar, kunst.carlos@inta.gob.ar
2 Universidad Nacional de Santiago del Estero, Facultad de Agronomía y Agroindustrias, Cátedra de Ecología, Av. Belgrano (S) 1912, Santiago del Estero, Argentina.
Recibido 12 de abril de 2017
Aceptado 27 de marzo de 2018
Publicado online 12 de septiembre de 2018
RESUMEN
El incremento de leñosas en detrimento de los pastos es un proceso muy común en la región chaqueña. El rolado de baja intensidad (RBI) es usado para incrementar la oferta forrajera de gramíneas, reducir la densidad y volumen de leñosas manteniendo la estructura arbórea. Los objetivos de este trabajo fueron: a) estudiar el efecto del RBI con siembra de Megathirsus maximus cv Gatton panic sobre la distribución de raíces de pastos y arbustos en un sitio “alto” con vegetación de bosque nativo, y b) probar la existencia del modelo de separación de nichos por exploración radicular, es decir, mayor densidad de raíces de gramíneas en el horizonte superior versus mayor densidad de raíces de arbustos en el horizonte inferior. Los tratamientos fueron: RBI y testigo (sin disturbar). El estudio se llevó a cabo en el Campo Experimental, INTA Santiago del Estero. En tres calicatas por tratamiento se contaron las raíces hasta profundidad = 1 m, diferenciándolas por tamaño y grupo funcional. Se realizó un análisis de varianza y las medias se compararon usando el test de Duncan. La mayor cantidad de raíces de gramíneas se localizó hasta los primeros 50 cm aproximadamente, mientras que las raíces de arbustos se encontraron a lo largo de todo el perfil. La evidencia indica que el mecanismo de separación de nichos por exploración radicular no ocurre. El RBI incrementó la densidad de raíces de gramíneas, principalmente de M. maximus, en los horizontes superiores, disminuyendo a medida que aumenta la profundidad. En el RBI hay una mayor densidad de plantas de M. maximus, comparada con los pastos del testigo. La densidad de raíces de arbustos fue mayor en el testigo. El RBI incrementó la densidad de raíces de menor tamaño de gramíneas. El RBI produjo una mejora de las condiciones físicas del suelo, lo que permitió el mayor desarrollo radical de M. maximus ejerciendo un efecto competitivo sobre las arbustivas.
Palabras clave: Arbustos; Chaco semiárido; Gramíneas; Rolado de baja intensidad; Raíces.
ABSTRACT
Woody plant encroachment and the reduction of grass density is a common process in Chaco region. Low intensity rolling chopping (RBI) is used to increase grass standing crop, reduce the density and volume of woody species, maintaining the tree structure. The objectives of this work were a) to evaluate the effect of RBI plus seeding of Megathirsus maximus cv Gatton panic on the root distribution of grasses and shrub species, in a “highland” site with forest and b) to examine the existence of niche separation by root exploration mechanism, it means high grass roots density in the top soil, versus high shrubs roots density in the subsoil. The study was conducted at the Experimental Station INTA Santiago del Estero, Argentina. Root density up to 1 m depth was assessed in three pits by treatment and differentiated by size and functional group, in four categories of soil depth. An analysis of variance was performed and root density means were compared using Duncan test. Most grass roots were located to 50 cm approximately, while the roots of shrubs were found along the entire soil profile. This evidence indicates that niche separation by root exploration mechanism not occurs. RBI increased grass roots density, mainly of M. maximus, at the soil upper layer. Its mean root density decreased with increasing depth. In RBI treatment, there are more M. maximus plants than in control. RBI increased the grasses fine roots density. Mean density of shrub roots was greater in the control than in the roller chopping. RBI improved the physical conditions of the soil, allowing a greater root density of M. maximus, and exerting a competitive effect over shrubs.
Keywords: Grasses; Rolling chopping; Roots; Semiarid Chaco; Shrubs.
INTRODUCCIÓN
El aumento de densidad y cobertura de leñosas, en general arbustivas, a expensas de los pastos es un problema generalizado en regiones semiáridas del mundo y representa uno de los cambios más notables en su fisonomía y composición botánica (Graz 2008, Archer et al., 2011). El incremento de áreas dominadas por leñosas en detrimento de las áreas dominadas por pastizales produce modificaciones en el paisaje, en la estructura de las comunidades vegetales, en la productividad de los pastos (Van Auken, 2009) y en la diversidad y funciones de los ecosistemas (Archer et al., 2011). El incremento de las leñosas puede ser ocasionado por diferentes causas: climáticas (Snyder y Tartowski, 2006) cambios en el régimen del fuego (Archer et al., 2011) y pastoreo (De Knegt et al., 2008). . El sobrepastoreo modificaría la coexistencia de pastos y arbustos al influir negativamente sobre el desarrollo de los primeros. Al aumentar las áreas de suelo desnudo, las raíces de leñosas penetran de manera más profunda en el suelo capturando más recursos, especialmente el agua (Walter, 1971).
En la región chaqueña argentina, la acción del sobrepastoreo, sobretala y cambios en el régimen de fuego homogeneizó la vegetación convirtiéndola en una formación vegetal lignificada denominada localmente “fachinal” (Adámoli et al., 1972). Actualmente, el tratamiento agronómico más empleado para modificar la estructura del fachinal y aumentar la aptitud ganadera es el “rolado” (Kunst et al., 2008). El rolado consiste en el pasaje de un cilindro de metal con cuchillas y lastrado con agua (rolo), traccionado por tractor o topadora, que aplasta las arbustivas e incrementa la oferta de gramíneas tanto en pastizales semiáridos como húmedos (Watts et al., 2006). En Argentina, existen antecedentes de rolados en el Caldenal-Monte Occidental (Aguilera y Steinaker, 2001; Adema et al., 2004); en Los Llanos de La Rioja (Blanco et al., 2013) y en Mendoza (Mora y Rosales 2014). En el Chaco occidental el rolado es generalmente acompañado con la siembra de pasturas subtropicales, como Megathyrsus maximus (Jacq) B. K. Simon & S. W. L. Jacobs var. Maximus. Cv Gatton (Kunst et al., 2012). En esta región existe información sobre el impacto del rolado en su enfoque general sobre la dinámica del agua (Kunst et al., 2003), la distribución de la radiación (Ledesma, 2006); la calidad biológica del suelo (Anríquez et al., 2005); y sobre la dinámica de las arbustivas y productividad de las pasturas (Kunst et al., 2012). La EEA INTA Santiago del Estero desarrolló el enfoque de rolado de baja intensidad (RBI) que emplea maquinaria de tamaño medio (rolo de ancho menor a 3 m y tractor de bajo porte vs. rolado tradicional: rolo de ancho 4 a 8 m con topadora). El RBI perturba al mínimo la vegetación presente para mantener los beneficios de las leñosas como sombra, aporte de mantillo y aporte de frutos (Kunst et al., 2008). No existe referencia sobre el efecto del RBI en la distribución de las raíces de pastos y arbustos. Las raíces actúan como reservorio, sostén y son la principal fuente de ingreso del carbono orgánico dentro del suelo actuando sobre el ciclado de carbono y nutrientes (Russell, 1977). El estudio de la dinámica radical es laborioso, intensivo y demanda mucho tiempo, por lo que son escasos los trabajos realizados, especialmente en ambientes con leñosas (Maccinis et al., 2010). Sin embargo, su estudio es uno de los caminos más importantes para entender la relación suelo-planta y planta-planta en un ecosistema (Escamilla, 1999) y su conocimiento constituye la base para las mejoras en los sistemas de manejo.
Este trabajo consistió en un estudio comparativo de la densidad de raíces de gramíneas y arbustivas en un sitio ecológico alto con vegetación de bosque bajo dos tratamientos: testigo y rolado con enfoque RBI.
Las hipótesis planteadas fueron:
1. El tratamiento RBI modifica la densidad de raíces. Se sabe que el RBI disminuye la cobertura de arbustos y permite el desarrollo de gramíneas (Kunst et al., 2008). Este efecto observado en la parte aérea, también se replicaría a nivel subterráneo en la densidad radical por grupo funcional (gramíneas y arbustos).
2. Los pastos presentan mayor densidad de raíces en el horizonte superior del suelo, mientras que las arbustivas presentan sus raíces a mayor profundidad, de acuerdo al mecanismo de separación de nichos (Walter, 1971). Una mayor densidad de raíces de pastos en el horizonte superior y de arbustivas en horizontes inferiores sería una evidencia que dichos grupos funcionales capturan recursos de nichos diferentes, implicando una partición de recursos en profundidad y un acceso casi exclusivo al agua y nutrientes de ambos nichos (Ward et al., 2013).
En forma complementaria, se estudió el efecto del RBI sobre la densidad aparente del suelo debido a que influye significativamente sobre la calidad de suelos y la distribución de raíces (De Abade y Lal, 2016).
MATERIALES Y MÉTODOS
El estudio se realizó en el Campo Experimental Francisco Cantos perteneciente a la EEA-INTA Santiago del Estero (28º 03´ LS; 64º 15´LW; 169 m s.n.m.), en la subregión Chaco occidental. El clima es semiárido, la temperatura media anual es de 21 °C y el promedio anual de lluvias es 574 mm, entre noviembre y mayo. Los suelos se clasificaron como Haplustoles torriorténticos (Anríquez et al., 2005). En la región chaqueña argentina, la vegetación nativa es un mosaico de bosques y sabanas y a una escala de 1:20.000, se distribuye a lo largo de una catena topográfica, desde el sitio ecológico alto hacia el bajo, pasando por el ecotono (Kunst et al., 2012). El sitio alto presenta generalmente una fisonomía vegetal de bosque de dos quebrachos, la media loma o ecotono, un parque, y el bajo, una sabana.
Trabajo de campo
Se trabajó en un ensayo de RBI (rolado de baja intensidad), instalado en 2006, en un sitio ecológico alto con vegetación de bosque. Los tratamientos fueron: (a) RBI, con siembra de M. maximum cv Gatton panic y (b) Testigo, sin tratamiento mecánico ni siembra. El RBI modifica al mínimo la estructura arbórea, dejando en pie la mayoría de los individuos de DAP > 5 cm, y aplasta/corta arbustos para mejorar la accesibilidad al tránsito y al forraje, generando un sistema silvopastoril (Kunst et al., 2008). En forma previa al tratamiento, la distribución espacial de los árboles fue desagregada, con una densidad promedio de 210 árboles.ha-1 y un área basal de 8.41 m2.ha-1 (Kunst et al., 2008).
Esta distribución y densidad fue respetada por el RBI. La densidad de arbustos se estimó en 3000 pl.ha-1; las especies dominantes son Celtis ehrenbergiana (Klotzsch) Liebm (tala), Acacia gilliesii Steud (garabato) y Capparis atamisquea Kuntze (atamisqui) (Kunst et al., 2012). Inmediatamente después de aplicado el RBI, la densidad de M. máximum se estimó en 14 ± 5 plantas.m-2, magnitud que había disminuido a 8 ± 4 plantas.m-2enel año del estudio (Kunst, datos sin publicar). No se observó germinación ni incremento en la densidad de pastos nativos. Para el presente estudio se seleccionaron dos parcelas del ensayo principal, una con tratamiento RBI y un testigo, ambas sin pastoreo. En cada una de ellas se ubicaron al azar tres calicatas de 3 m de largo, 1 m de ancho y 1 m de profundidad, siguiendo a Macinnis et al., (2010). Estas se excavaron alejadas 2-3 m del fuste principal para evitar encontrar una raíz importante del árbol. Sobre una de las caras de cada calicata se colocó una malla de alambre cuadriculada (5 x 5 cm) (Knoop y Walker, 1985). Cada calicata se subdividió en 3 submuestras de 1 m2 (1 x 1 m) y cada submuestra se dividió en cuatro clases de profundidad: 0-25 cm; 25,1-50 cm, 50,1-75 cm y 75,1-100 cm.
Identificación y clasificación de raíces
El trabajo de campo se realizó entre diciembre de 2014 y enero de 2015. Se contó el número de raíces expuestas en cada cuadrícula por submuestra y clase de profundidad, clasificándolas según las siguientes categorías (a) grupo funcional: arbustivo o graminoso; y (b) tamaño: raicillas (diámetro < 0,1 cm), pequeñas (diámetro = 0,11 - 0,5 cm); medianas (diámetro = 0,51 a 1,5 cm) y grandes (diámetro ˃ 1,51 cm). Los datos se expresaron en raíces.m-2. Los sistemas radicales de los árboles no fueron considerados en este estudio. Hubo ausencia de latifoliadas herbáceas.
Densidad aparente
Para estudiar el efecto del RBI sobre la Dap, la misma fue determinada a tres profundidades: 0 a 0,05 m; 0,5 m y 1 m en las mismas calicatas. Las muestras se tomaron con un cilindro de volumen conocido, y la Dap se determinó por gravimetría (Klute, 1986).
Análisis estadístico e interpretación
La densidad de raíces se empleó como un indicador de su distribución en el perfil. Se realizó un análisis de la varianza empleando raíces.m-2 como variable dependiente y como variables independientes: tratamiento, calicata, grupo funcional, profundidad, tamaño de raíces y sus respectivas interacciones. Se evaluó la interacción entre factores. Los datos fueron transformados a rangos. Se empleó el paquete estadístico InfoStat versión 2015 (Di Rienzo et al., 2008). La Dap fue sometida a un ANVA, con las variables independientes; tratamiento, profundidad y su interacción. Para interpretar el efecto del RBI sobre esta variable, se comparó la Dap media observada con un umbral crítico para penetración y acceso a nutrientes (Dap ≈1,40 g.cm-3) sugerido para suelos francos (Wilson et al., 2013). Las medias fueron separadas mediante el test de Duncan, con un α = 0,05 para establecer significancia estadística.
RESULTADOS
Densidad de raíces
El factor de clasificación tamaño explicó el 37% de la variabilidad total de la densidad de raíces y grupo funcional, el 5% (ambos con p<0,001). Entre los factores restantes se destacaron profundidad (p<0,001; 2,7%) y las interacciones tratamiento*grupo funcional (p<0,0001; 4,9%), tratamiento*tamaño de raíces (p<0,0043; 1,3%) y tamaño de raíces*grupo funcional (p<0,0005; 8,6%). Calicata no fue significativa. A continuación se hará foco en los factores de interés con efecto significativo (fig. 1). Tratamiento. La densidad de raíces fue significativamente mayor en el testigo que en el RBI (p=0,0280) (media ± DE: 310 ± 711 vs. 207 ± 477 raíces.m-2 respectivamente), englobando grupo funcional, tamaño y profundidad. Grupo funcional. En términos absolutos, la densidad de raíces de arbustivas fue significativamente mayor que la de gramíneas, (media ± DE: 401 ± 731 versus 115 ± 402,07 raíces.m-2, respectivamente: p<0,0001), englobando tratamiento, tamaño de raíces y profundidad de suelo (fig. 1). La densidad media de raíces de gramíneas fue mayor en el RBI y pertenecieron a M. máximum, mientras que en el testigo pertenecieron a los géneros Trichloris, Setaria, y Gouinia. Tamaño de raíces. La densidad de raicillas (diámetro < 0,1 cm) fue significativamente mayor que las otras 3 categorías (p<0,0001) (fig. 1).
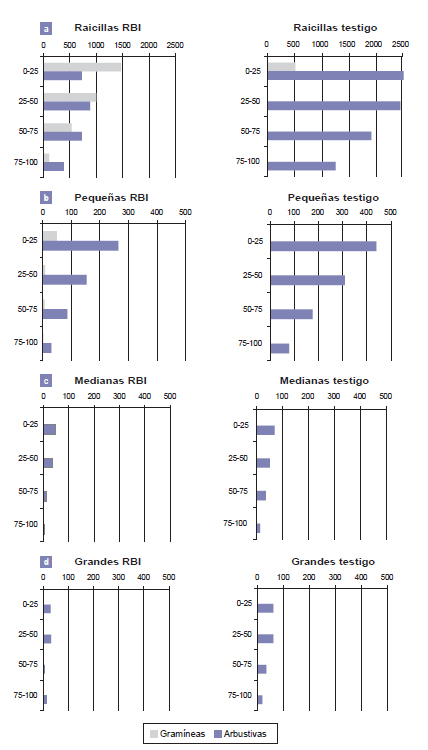
Figura 1. Densidad media de raíces de gramíneas y arbustivas (número.m-2) en dos tratamientos: RBI (rolado de baja intensidad) con siembra de Megathirsus maximus cv. Gatton panic; y Testigo (bosque sin disturbar) para cuatro clases de profundidad de suelo. Campo Experimental Francisco Cantos, INTA EEA Santiago del Estero, sitio ecológico alto con vegetación de bosque de dos quebrachos. Referencias: Tamaño de raíces: (a) raicillas: diámetro < 0,1 cm; (b) pequeñas diámetro = 0,11 - 0,5 cm; (c) medianas, diámetro = 0,51 a 1,5 cm y d) grandes diámetro ˃ 1,51 cm.
Profundidad de suelo. La mayor densidad media de raíces (arbustivas + gramíneas), independiente del tamaño, se observó en los primeros 25 cm, y disminuyó a medida que aumentó la clase de profundidad (fig. 1). La reducción de la densidad media de raíces fue más evidente en las gramíneas. La densidad de raíces de arbustivas fue elevada hasta los 50-75 cm, y luego disminuyó. Tratamiento*grupo funcional. La densidad de raíces de arbustos en el testigo fue mayor a la de gramíneas en todo el perfil del suelo: 588,2 ± 888 raíces.m-2 versus 32,47 ± 135 raíces.m-2, respectivamente. En el testigo, la mayor densidad media de raíces de gramíneas nativas se observó en la clase 0-25 cm de profundidad, mientras que las raíces de arbustos se distribuyeron en todo el perfil evaluado. En el RBI, M. máximum fue dominante y sus raíces se extendieron hasta los 100 cm de profundidad y con una densidad apreciable. Tratamiento*tamaño de raíces. La densidad de raicillas fue 775 ± 725 raíces.m-2 en el rolado y 1064 ± 1100 raíces.m-2 en el testigo, pero considerando ambos grupos funcionales en todo el perfil. En el testigo solo se cuantificaron raicillas de gramíneas nativas en la primera clase de profundidad, mientras que en el RBI se observaron además los otros tamaños hasta el metro de profundidad. Grupo funcional*tamaño de raíces. En gramíneas no se observaron raíces con diámetro > 1,51 mm. Las raicillas de arbustos alcanzaron un valor medio de 681± 610 raíces.m-2 en el RBI, versus 2016±727 raíces.m-2 en el testigo. La cantidad de raicillas disminuyó a medida que aumentó la profundidad (fig. 1).
Densidad aparente
Los valores de Dap observados oscilaron entre 1,03 y 1,47 g.cm-3. La Dap no fue influenciada por tratamiento (p= 0,75), pero sí por profundidad de suelo (p<0,0533). No se observó diferencia significativa entre RBI y testigo (1,21 g.cm-3 versus 1,23 g.cm-3, respectivamente). La mayor Dap media se observó a mayor profundidad (fig. 2). La Dap media no superó en ningún caso el umbral crítico propuesto para suelos francos.
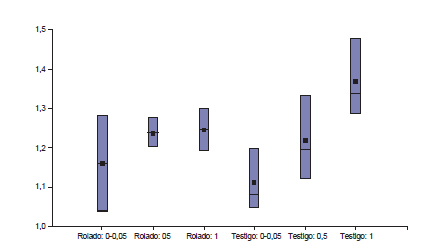
Figura 2. Densidad aparente promedio del suelo (Dap) por tratamientos RBI (rolado de baja intensidad) y Testigo a 3 clases de profundidad de suelo (0-0,05 m; 0,5 m y 1 m, respectivamente. Campo Experimental Francisco Cantos, INTA EEA Santiago del Estero, sitio ecológico alto con vegetación de bosque de dos quebrachos.
DISCUSIÓN
La primera hipótesis plantea que el tratamiento RBI con siembra de M. maximus modifica la densidad media de raíces. Esta hipótesis debe ser aceptada solo en el caso de raicillas. Las dos interacciones de interés en relación con esta hipótesis son tratamiento*grupo funcional y tratamiento*tamaño de raíces. El principal efecto del RBI fue la modificación en la proporción del tamaño y del grupo funcional al que pertenecían las raíces. Las raíces de arbustos disminuyeron en el RBI con respecto al testigo, y aumentaron las de pastos, especialmente las raicillas, presentando una mayor profundidad de exploración (fig. 1). Este incremento en la densidad media de raicillas de gramíneas observado en el RBI podría deberse a distintas razones, a saber:
• Adaptaciones. Las especies modifican sus estructuras para optimizar la captura de los recursos más limitantes, por ej. mayor cantidad de raíces finas cuando el suministro hídrico se agota (Wang et al., 2016).
• Competencia. Al contrario de lo observado en el RBI, la densidad media de raíces de arbustivas en el testigo fue superior y se presentaron en todo el perfil. Este hecho podría ser una característica intrínseca del sitio ecológico alto, o podría deberse a que las arbustivas en este sitio compiten con las herbáceas y gramíneas nativas hasta el punto de reducir su densidad. Se observó una alta variabilidad de la densidad de raíces verticalmente, lo que sugiere una distribución heterogénea de recursos en el suelo (Mordelet et al., 1997).
• Cambios en la disponibilidad de recursos. El RBI modifica localmente la dinámica del agua (Kunst et al., 2003 y 2012), y por lo tanto el crecimiento de raicillas de ambos grupos vegetales. El RBI modifica la distribución de la radiación bajo la canopia de los árboles (Ledesma, 2006). M. maximum se adapta muy bien la sombra y genera un sistema radical denso (Schroth, 1999), algunas gramíneas nativas pueden desarrollarse, pero lo hacen con un menor porte y un sistema radical menos abundante.
La segunda hipótesis planteada fue que en el RBI los pastos presentarían mayor densidad media de raíces en el horizonte superior. La evidencia recolectada en este estudio indica que esta hipótesis debe ser rechazada. La mayor densidad de raíces de gramíneas y arbustos se observó en ambos tratamientos en los primeros 25 cm, sugiriendo competencia por recursos, concordando con Mordelet et al. (1997), Kulmatiski et al. (2010) y February et al. (2013). Se debe tener en cuenta, sin embargo que la bibliografía no distingue entre árboles y arbustos. La coexistencia pastos-leñosas en ecosistemas donde la precipitación es estacional y los pastos tienen un período de dormancia, como en la región chaqueña (Walker, 1981), podría ser explicada mediante el mecanismo de separación de nichos por exploración radical (Walter, 1971). Los pastos absorberían el agua de los horizontes superiores del suelo mientras que los arbustos captarían agua de horizontes más profundos, (Belsky et al., 1997). Los pastos son competidores superiores frente a los arbustos en las capas superficiales del suelo, mientras que las leñosas lo son en las capas inferiores (Knoop y Walker, 1985). La distribución superficial de raíces observada en este estudio no presenta en primera instancia evidencia para sostener el modelo de separación de nichos. Sin embargo, Scheiter y Higgins (2007) indican que a pesar de que el mecanismo de separación de nichos no se dé en la realidad, aún es posible la coexistencia de pastos y leñosas. La habilidad competitiva de las especies está correlacionada con la densidad de raíces, especialmente las finas, debido a que estas son un indicador de la ocupación del suelo (Casper y Jackson, 1997). La alta densidad total de raíces (arbustivas y gramíneas) en el horizonte superior del suelo en este estudio, y su disminución en profundidad, independientemente del tratamiento y factores de clasificación, coinciden con la información proporcionada por la bibliografía para los dos grupos funcionales y para distintos ecosistemas (Mordelet et al., 1997, Makita et al., 2011). Este resultado sugiere que los sistemas radiculares en el sitio ecológico alto en ambos grupos funcionales son superficiales, característica que estaría asociada a la ubicación espacial de la mayor disponibilidad de agua y nutrientes en esos ambientes.
La Dap es una propiedad del suelo considerada dinámica, debido a que puede ser afectada por el manejo. Los resultados obtenidos sugieren que el aplastamiento de las arbustivas mediante el RBI no modificó de manera significativa este atributo, aun después de un plazo considerable luego de la perturbación mecánica. La alta varianza en la densidad de raíces observada en el presente estudio se atribuye a la presencia de sectores en los perfiles de suelo que no presentaban ninguna raíz, una característica intrínseca de este atributo vegetal. Ello sugiere una distribución heterogénea de nutrientes en el suelo (Wang et al., 2016). Las limitaciones de este experimento consisten en ser una “foto instántanea” de un proceso dinámico (February e Higgins, 2010) y también el escaso número de repeticiones por tratamiento, debido a la laboriosidad del muestreo. Su mérito es ser el primer estudio que se realiza sobre la distribución de raíces en un sitio alto con vegetación de bosque en la región chaqueña, tratado con RBI (perturbación) y sin tratar.
CONCLUSIONES E IMPLICANCIAS PARA EL MANEJO
En este estudio el RBI permitió conservar la estructura arbórea y reducir la cobertura de arbustivas sin eliminar este último estrato. Se modificaron las proporciones de raíces de gramíneas y arbustos y el tamaño de estas, incorporándose al perfil del suelo raíces de la pastura M. máximum cv Gatton panic. El rolado sin la siembra de pasturas puede incrementar la oferta de gramíneas nativas, sin embargo esta respuesta generalmente no es importante en sitios ecológicos altos con vegetación de bosque. En este sentido, la implantación de M. maximum genera una cobertura del suelo e incrementa el espacio radical explorado.
El RBI modificó las proporciones de raíces de gramíneas y arbustos y el tamaño de estas en un sitio ecológico alto con bosque. En este estudio, la coexistencia pastos y arbustos no pudo explicarse mediante el mecanismo de separación de nichos por exploración radical. Las raíces de ambos grupos funcionales compartan el mismo espacio por lo tanto las prácticas culturales para aplicar deben procurar evitar las pérdidas del agua del sistema, conservando la cobertura y las demás funciones ecosistémicas. Además, en un sistema silvopastoril creado con un RBI, el manejo debe evitar el sobrepastoreo y favorecer la implantación correcta de las pasturas implantadas, de manera de poder desarrollar un denso sistema radical que le permita la absorción de agua y nutrientes, la competencia con los arbustos y el mayor aporte al stock de carbono al suelo.
AGRADECIMIENTOS
Se agradece la contribución de los Proyectos INTA: PNFOR: Interacciones ecológicas en sistemas silvopastoriles; PNPA-PE 1126074 “Desarrollo, Integración y Transferencia de Tecnologías para Manejo Sustentable de Servicios de la Vegetación Natural para Fines Ganaderos” y los proyectos regionales “Contribuciones al desarrollo integral del oeste de Santiago del Estero” y “Contribuciones al desarrollo integral del centro de Santiago del Estero”.
BIBLIOGRAFÍA
1. ADÁMOLI, J.; NEUMANN, R.; RATIER DE COLINA, A.; MORELLO, J.1972. Chaco aluvional salteño. Revista de Investigaciones agropecuarias. INTA, Buenos Aires, Rep. Argentina Serie 3, Clima y Suelo, Vol. ix, N.o 5. [ Links ]
2. ADEMA, E.; BUSCHIAZZO, D.; BABINEC, F.; RUCCI, T.; GOMEZ HERMIDA, V. 2004. Mechanical control of shrubs in a semiarid region of Argentina and its effect on soil water content and grassland productivity. Agricultural Water Management 68 (2004) 185-194. [ Links ]
3. AGUILERA, M.; STEINAKER, D. 2001. Rolado y siembra en arbustales semiáridos de San Luis. En: PANIGATTI, J.; BUSCHIAZZO, D.; MARELLI, H. (Eds.). Siembra Directa II. INTA. pp. 289-302. [ Links ]
4. ANRIQUEZ, A.; ALBANESI, A.; KUNST, C.; LEDESMA, R.; LOPEZ, C.; RODRIGUEZ, A.; GODOY, J. 2005. Rolado de fachinales y calidad de suelos en el chaco occidental, Argentina. Ciencia del suelo (Argentina) 23 (2): 145-157. [ Links ]
5. ARCHER, S.; DAVIES, K.; FULBRIGHT, T.E.; MCDANIEL, K.; WILCOX, B.; PREDICK, K. 2011. Brush Management as a Rangeland Conservation Strategy: A Critical Evaluation. En: BRISKE, D.D. (Ed.). Conservation Benefits of Rangeland Practices: assessment, recommendations, and knowledge gaps. Unites States Department of Agriculture, Natural Resources Conservation Service, p. 429. [ Links ]
6. BLANCO, L.; NAMUR, P.; FERRANDO, C.; RETTORE, A.; NAMUR, P.; AVILA, R.; MOLINA, J.; ORIONTE, E. 2013. Evolución de la vegetación después del rolado y siembra de pastos nativos en La Rioja. Revista de la Facultad de Agronomía UNLPam Vol. 22. [ Links ]
7. BELSKY, A. 1997. Tree grass ratios in East African savannas: a comparison of existing models. Journal of biogeography. 1990. 17,483-489. [ Links ]
8. CASPER, B.; JACKSON, R. 1997. Plant competition underground. Annual Review of Ecology and Systematics 28: 545-570. [ Links ]
9. DE ABADE, V.; LAL, R. 2016. Towards a standard technique for soil quality assessment. Geoderma 265: 96-102. [ Links ]
10. DE KNEGT, H.; GROEN, T.A.; VAN DE VIJVER, C., PRINS, H.H.; LANGEVELDE, F. 2008. Herbivores as architects of savannas: inducing and modifying spatial vegetation patterning. Oikos 117: 543554. doi: 10.1111/j.2008.0030-1299.16403.x [ Links ]
11. DI RIENZO, J.A.; CASANOVES, F.; BALZARINI, M.G.; GONZALEZ, L.; TABLADA, M.; ROBLEDO, C.W. 2008. InfoStat, versión 2008, Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. [ Links ]
12. ESCAMILLA, B.J.A. 1999. Estudios de suelos y de ecosistemas vegetales. En: ORELLANA, R.; ESCAMILLA, B.J.A.; LARQUE SAAVEDRA, A. (Eds.). Ecofisiología Vegetal y Conservación de los Recursos Genéticos. Centro de Investigación Científica Yucatán. Mérida. pp.121-123. [ Links ]
13. FEBRUARY, E.; HIGGINS, S. 2010. The distribution of tree and grass roots in savannas in relation to soil nitrogen and water. South African Journal of Botany 76: 517-523. [ Links ]
14. FEBRUARY, E.; COOK, G.; RICHARDS, A. 2013. Root dynamics influence tree–grass coexistence in an Australian savanna. Austral Ecology 38: 66-75.
15. GRAZ, F.P. 2008. The woody weed encroachment puzzle: gathering pieces- Ecohydrology 1: 340-348. [ Links ]
16. KLUTE, A. 1986. Methods of soil analysis. Part 1: Physical and mineralogical methods. Agronomy 9, ASA, ASSA, inc, Publisher Madison, Wilconsin. EUA. p.1188. [ Links ]
17. KNOOP, W.T.; WALKER, B.H. 1985. Interactions of woody and herbaceous vegetation in a southern African savanna. Journal of Ecology, Vol. 73, N.° 1. pp. 235-253. [ Links ]
18. KULMATISKI, A.; BEARD, K.; VERWEIJ, R.; FEBRUARY, E. 2010. A depth-controlled tracer technique measures vertical, horizontal and temporal patterns of water use by trees and grasses in a subtropical savanna. New Phytologist 188: 199-209. [ Links ]
19. KUNST, C.; LEDESMA, R.; BASAN, M.; ANGELLA, G.; PRIETO, D.; GODOY, J. 2003. Rolado de fachinales e infiltración de agua en el suelo en el Chaco occidental argentino. RIA. Revista de Investigaciones Agropecuarias 32: 105-122. [ Links ]
20. KUNST, C.; LEDESMA, R.; NAVALL, M. 2008. RBI: Rolado Selectivo de Baja Intensidad. Ediciones INTA, pp. 139. [ Links ]
21. KUNST, C.; LEDESMA, R.; ALBANESI, A.; ANRIQUEZ, A.; BRAVO, S.; GODOY, J. 2012. Disrupting woody steady states in the Chaco region (Argentina): Responses to combined disturbance treatments. Ecological Engineering. 42: 42-53. [ Links ]
22. LEDESMA, R. 2006. Desarrollo de sistemas ganaderos: una alternativa de manejo en ecosistemas degradados del Chaco semiárido. Universidad Nacional de Santiago del Estero, Santiago del Estero, Argentina. Tesis de maestría. p.134. [ Links ]
23. MACINNIS, C.M.O.; FUENTES, S.; O’GRADY, A.P.; PALMER, A.R.; TAYLOR, D.; WHITLEY, R.J.; EAMUS, D. 2010. Root biomass distribution and soil properties of an open woodland on a duplex soil. Plant and Soil, 327(1-2), 377-388.
24. MAKITA, N.; HIRANO, Y.; MIZOGUCHI, T.; KOMINAMI, Y.; DANNOURA, M.; ISHII, H.; FINÉR, L.; KANAZAWA, Y. 2011. Very fine roots respond to soil depth: biomass allocation, morphology, and physiology in a broad-leaved temperate forest. Ecological Research 26: 95-104. [ Links ]
25. MORA, S.; ROSALES MERCADO, I. 2014. El rolado en Mendoza. Ediciones INTA. p. 42. [ Links ]
26. MORDELET, P.; MENAUT, J.C.; MARIOTTI, A. 1997. Tree and grass rooting patterns in an African humid savanna. Journal of Vegetation Science 8: 65 Very fine roots respond to soil depth: biomass allocation, morphology, and physiology in a broad-leaved temperate forest 70. [ Links ]
27. RUSSELL, R. 1977. Plant roots systems; their function and interaction with the soil. McGraw – Hill Book Company, Londres, p. 298.
28. SCHEITER, S.; HIGGINS, S. 2007. Partitioning of root and shoot competition and the stability of savannas. The American Naturalist 170: 587-601. [ Links ]
29. SCHROTH, G. 1999. A review of belowground interactions in agroforestry, focussing on mechanisms and management options. Agroforestry Systems 43: 5-34. [ Links ]
30. SNYDER, K.; TARTOWSKY, S.L. 2006. Multi-scale temporal variation in water availability: Implications for vegetation dynamics in arid and semi-arid ecosystems. Journal of Arid Environments 65 (2006) 219-234. [ Links ]
31. VAN AUKEN, O.W. 2009. Causes and consequences of woody plant encroachment into western North American grasslands. Journal of Environmental Management 90, 2931-2942. [ Links ]
32. WALKER, B.H.; LUDWIG, D.; HOLLING, C.S.; PETERMAN, R.M. 1981. Stability of semiarid savanna grazing systems. Journal of Ecology N.° 2, pp. 473-498. [ Links ]
33. WALTER, H. 1971. Ecology of tropical and subtropical vegetation. En: WALKER, B.H.; LUDWIG, D.; HOLLING, C.S.; PETERMAN, R.M. (Ed.). Stability of semiarid savanna grazing systems. Journal of Ecology, Vol. 69. N.° 2, pp. 473-498. [ Links ]
34. WANG, W.; WU, X.; HU, K.; LIU, J.; TAO, J. 2016. Undestorey fine root mass and morphology in the litter and upper soil layers of three Chinese subtropical forests. Plant and Soil 406: 219-230. [ Links ]
35. WARD, D.; WIEGAND, K.; GETZIN, S. 2013. Walter’s two layer hypothesis revisited: back to the roots! Oecologia 172: 617-630.
36. WILSON, M.; SASAL, M.; CAVIGLIA, O. 2013. Critical bulk density for a Mollisol and a Vertisol using least limiting water range: Effect on early wheat growth. Geoderma 192: 354-361. [ Links ]

