











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
En Argentina, las investigaciones arqueológicas que incluyen análisis de microrrestos vegetales adquirieron mayor preponderancia y aumentaron considerablemente en los últimos años a lo largo de la Cuenca del Plata (Zucol y Loponte 2008; Pérez Meroni et al. 2010; Bonomo et al. 2011; Cornero y Rangone 2015; Colobig et al. 2016; Colobig y Ottalagano 2016; Auge et al. 2018; entre otros). El mismo proceso se ve reflejado en las producciones científicas sobre las costas uruguayas (ie.Beovide 2011; Beovide y Campos 2015). La sistematización de estos trabajos incentivó la identificación de una variedad de taxones presentes en el registro arqueológico en zonas donde la preservación de macrorrestos vegetales es escasa.
En este contexto, comenzaron a desarrollarse estudios centrados en la interrelación entre las plantas y grupos humanos de época prehispánica asentados en la porción meridional del litoral rioplatense, zona distintiva por presentar una gran riqueza biocultural, hoy declarada y conocida como Reserva Mundial de Biosfera Parque Costero del Sur (UNESCO). La trayectoria de nuestras investigaciones enfatiza y enlaza los resultados de la identificación taxonómica con la información procedente de otros registros, camino que nos permite conocer con mayor profundidad los contextos socioculturales en los que interactuaban las personas, la alfarería y las plantas en el pasado.

Figura 1: a) ubicación del sitio arqueológico Las Marías, emplazado dentro del Parque Costero del Sur. b) excavación de Las Marías donde se observa la presencia del bosque de tala (Foto: M. Auge, 2014).
El objetivo de este trabajo es presentar los resultados del análisis de microrrestos vegetales recuperados en los diferentes tipos morfo-funcionales de cerámica del sitio Las Marías, ubicado en el actual partido de Magdalena (Magdalena, provincia de Buenos Aires) (Figura 1). Estos estudios aportan información sobre las prácticas asociadas al manejo y aprovechamiento de plantas por las poblaciones humanas que incluyen los conocimientos y creencias involucrados en la relación de los seres humanos y su ambiente (Pochettino y Lema 2008). De esta forma, nos introducen en la discusión sobre el tipo de prácticas de manejo del entorno vegetal que habrían implementado estos grupos, y la importancia de estos a nivel regional.
El sitio Las Marías: estado actual de conocimiento
En el partido de Magdalena, a 1 km de la costa rioplatense se localiza el sitio Las Marías (Figura 1). Se trata de un asentamiento a cielo abierto datado en 1820 + 50 años AP (Pogonias cromis) y 1590 + 40 años AP (Lama guanicoe) (Paleo et al. 2002, Paleo y Pérez Meroni 2007). Se emplaza en zonas elevadas producidas por la acumulación de sedimentos conchiles depositados por ingresiones marinas, que se encuentran dispuestos en forma paralela a subparalela a las costas del Río de la Plata y proporcionaron el sustrato necesario para el desarrollo del “bosque de tala” (Paleo y Pérez Meroni 2004, 2007). Esta formación boscosa es una de las características florísticas más destacables del área y se compone principalmente por Celtis tala Gillies ex Planch. (tala), Scutia buxifolia Reissek (coronillo), Jodina rhombifolia (Hook. & Arn.) Reissek (sombra de toro), por mencionar las más abundantes (Delucchi y Torres Robles 2009). La ocupación humana habría encontrado en el bosque refugio y protección que, junto a ambientes circundantes asociados a pastizales, humedales y el río, ofrecen una variada gama de recursos. Se propuso que las poblaciones que ocuparon Las Marías tenían una economía basada en la caza, pesca y recolección de gran cantidad de especies locales. En el registro arqueofaunístico y asociados fundamentalmente a actividades antrópicas se destacan los peces (principalmente Pogonias cromis y Pterodoras granulosus), aves (ie. Rhea americana) y mamíferos de diversos tamaños como cérvidos (Ozotoceros bezoarticus y Blastocerus dichotomus), carnívoros terrestres (Puma concolor) y marinos (Arctocephalus australis, Otaria flavescens), y roedores caviomorfos (ie. Cavia aperea, Ctenomys talarum). Se evidencia el consumo integral de muchas especies, incluyendo el aprovechamiento tecnológico para confeccionar instrumentos óseos como alisadores, puntas y punzones (Day Pilaría 2018). El hallazgo de elementos alóctonos como material lítico y restos óseos de Lama guanicoe permitieron proponer que estos grupos podrían haber participado en redes de interacción a escala regional (Paleo y Pérez Meroni 2005-2006).
La cerámica constituye el registro más abundante y se halla con un alto grado de fragmentación. Desde hace 30 años, el equipo del Laboratorio de Análisis Cerámico (LAC), de la Facultad de Ciencias Naturales y Museo (UNLP), implementó diferentes estudios arqueométricos para el abordaje de la cerámica: análisis de la decoración, difractometrías de rayos X, cortes delgados, análisis de porosidad, estudios experimentales y análisis químicos de residuos orgánicos. La tarea de remontaje permitió reconocer al menos tres tipos básicos de formas, tamaño y funciones diferenciadas (Figura 2): 1) contenedores de procesamiento y/o servicio, 2) contenedores de almacenaje y 3) instrumentos de transferencia, conocidas como alfarerías tubulares (Paleo y Pérez Meroni 2005-2006; 2008; Ghiani Echenique y León 2014; Ghiani Echenique et al. 2017).

Figura 2: a) Contenedores de procesamiento y/o servicio, b) Contenedores de almacenaje y c) Instrumentos de transferencia.
Entre la multiplicidad de hallazgos que componen el registro arqueológico de Las Marías, los vegetales son los que presentan menos visibilidad en forma macroscópica. Los carbones no cuentan con condiciones de preservación favorables debido a la humedad y el pH del suelo sumado a la importante actividad biótica. Cabe destacar que la conservación del material vegetal se relaciona también con características intrínsecas de cada taxón y con factores antrópicos, como la selección diferencial por parte de las poblaciones humanas de determinadas especies y órganos vegetales, los tipos de procesamientos realizados, patrones culturales de descarte y depositación (Ford 1979).
Los primeros análisis arqueobotánicos de la zona se realizaron sobre fragmentos de contenedores de procesamiento con marcas de uso en forma de adherencias y/o hollín (Pérez Meroni et al. 2010; Lema et al. 2010). Estos estudios identificaron células pétreas y almidones de Celtis tala, y cristales y almidones de Jodina rhombifolia. Algunos granos de almidón presentaron distintos grados de alteración, como baja a nula birrefringencia, y fisuras en la zona del hilio. Por otro lado, se hallaron algunas estructuras ubicuas en el reino vegetal (como granos de almidón esféricos). Estas tareas aportaron evidencia directa del uso de vegetales, permitieron identificar procesos de manejo del entorno y distintas prácticas de procesamiento y consumo de plantas de la rivera rioplatense. En este sentido, en la presente contribución continuamos con esta línea de trabajo y pretendemos profundizar a futuro con su aplicación a nuevos registros arqueológicos de la costa del Río de la Plata.
Materiales y métodos
Colección de referencia
Con el propósito de generar material comparativo para el análisis de microrrestos vegetales elaboramos una colección de referencia de especies relevantes. Los taxones que conforman la colección fueron seleccionados en función de las especies propias del bosque de tala mencionadas en distintas fuentes documentales con algún uso o procesamiento que podría vincularse al registro cerámico que aquí analizamos (como alimentos, bebidas, medicinas). También se incluyeron plantas domesticadas mencionadas para regiones vecinas al área de estudio. Para tal fin, analizamos fuentes de los siglos XVI al XIX (Ramírez 2007 [1528]; Lopes de Sousa 1927 [1530-1532]; Schmidel 1903 [1534-1554]; Vargas Machuca 1892 [1599]; Paucke 2010 [1749-1767] y Hieronymus 1882), y trabajos científicos de los siglos XX y XXI (Lévi-Strauss 1952; Arenas 2003; Keller 2007, 2008; Pochettino et al. 2016; Doumecq 2019). Para su abordaje tuvimos en cuenta las estrategias metodológicas propuestas por Nacuzzi (2002), entre las que se propone el análisis textual y contextual de los documentos.
La colección de referencia comprende, además de la caracterización de las estructuras en estado fresco, la descripción de modificaciones producidas por distintos tipos de procesamiento experimental (machacado, tostado, horneado, hervido y fermentado) observadas según el taxón. Para la descripción de las estructuras micrográficas utilizamos los parámetros consensuados en el International Code for Starch Nomenclature (ICSN 2011) y el International Code for Phytolith Nomenclature 2.0 (Neumann et al. 2019). En el caso de plantas cultivadas y/o ampliamente estudiadas recurrimos a bibliografía especializada (ie. Winton y Winton 1932; Reichert 1913). En esta oportunidad tendremos en cuenta el conjunto de microrrestos hallados en las muestras arqueológicas, ya que estas forman parte del contexto relacional que estamos analizando y son imprescindibles para el alcance de una determinación taxonómica, así como para el reconocimiento de prácticas de procesamiento.
Material arqueológico
Seleccionamos para este trabajo trece fragmentos cerámicos correspondientes a las tres categorías morfo-funcionales que rotulamos como “B”. Se trata de nueve fragmentos de contenedores de procesamiento y/o servicio, dos de contenedores de almacenaje y dos de instrumentos de transferencia. La diferencia en la cantidad de elementos seleccionados para cada grupo cerámico se corresponde con la frecuencia de estos en el registro arqueológico. Los fragmentos de contenedores de procesamiento y/o servicio superan en número a la cantidad de los otros tipos morfofuncionales, de la misma manera que presenta una mayor diversidad de formas y tamaños.
Para la recuperación de microrrestos vegetales raspamos los fragmentos cerámicos en su cara interna y externa con instrumental metálico esterilizado a fuego. Con el fin de evitar la contaminación de la muestra se trabajó en ambiente cerrado y con superficies de trabajo limpias. Escogimos como medio de montaje aceite de inmersión ya que permite la observación de las estructuras vegetales en sus tres dimensiones. Realizamos una observación directa, sin tratamientos químicos previos para preservar la totalidad de las estructuras vegetales, en Microscopio Óptico Leica MDLM (MO) bajo aumentos de 20x y 50x, con luz normal y polarizada, y fotografiamos con cámara Toup CamTM modelo UCMOSO5100KPA.
Las condiciones edáficas de la región de estudio requieren que determinan que los materiales arqueológicos sean sometidos a tareas de acondicionamiento para su análisis e identificación. La reciente apertura de nuevos sectores de excavación en el sitio Las Marías nos permitió incluir tres muestras testigo previo a actividades de lavado, siglado y sistematización. De esta forma, analizamos tres fragmentos de contenedores de procesamiento y/o servicio siguiendo el protocolo de recuperación de microrrestos vegetales mencionado anteriormente. Estas muestras control, que agrupamos bajo la categoría “M”, elevan la cantidad de fragmentos analizados a un total de dieciséis.
En los casos en los que registramos costras carbonizadas adheridas a bases de contenedores de procesamiento, para su observación en MO las tratamos con ácido láctico durante doce días y disgregamos mecánicamente con un agitador automático (tres sesiones de veinte minutos cada una). En estas muestras, el ácido láctico también sirvió como medio de montaje. Este protocolo, testeado de forma favorable en residuos carbonizados recuperados en pipas (Lema et al. 2015; Auge et al. 2018), permite aclarar y mejorar la visión al MO del material, sin perder la diversidad de estructuras vegetales.
Resultados
Material de referencia
La información de las plantas relevadas en las fuentes documentales y trabajos etnobotánicos que incorporamos a la colección de referencia fue sistematizada en la Tabla 1. Se trata de vegetales que tienen elementos con parámetros comparables a los observados en las muestras arqueológicas, algunas son taxones de crecimiento espontáneo en la zona de estudio y otras son especies domesticadas de circulación macrorregional. En la misma, también se consignan los tipos de procesamiento asociados a distintos órganos vegetales, así como usos y prácticas sociales en los que intervenían estas plantas. Cabe aclarar que Lopes de Sousa [1530-1532] en su viaje hace contacto con nativos en la zona de la desembocadura del río Santa Lucía y delta del Paraná, con quienes realiza intercambios de pescado, venado y “oveja”, pero no incluye vegetales domesticados como sí lo hacen otros viajeros que transitan la región paranaense (Ramírez [1528] y de Schmidel [1534-1554]). Sin embargo menciona que en períodos de hambruna “nam comemos senam enam ervas cozidas” (Lopes de Sousa 1927 [1530-1532]: 318). De la misma forma, distintos exploradores señalan que se valieron de hierbas, posiblemente silvestres, para alimentarse cuando no había otros recursos (Ramírez [1528]; Lopes de Sousa [1530-1532]; Schmidel [1534-1554]).
En la Tabla 1 se puede observar una serie de usos asociados a órganos que tienen su correlato en el registro arqueológico aquí presentado. Sin embargo, en las fuentes analizadas también se mencionan otras formas de relacionarse con las mismas plantas en función de otras partes y propiedades de estas. Es el caso del uso de semillas y hojas de Manihot esculenta como alimento y medicina (Hieronymus 1882; Keller 2007, 2008). Árboles y arbustos de crecimiento espontáneo como Celtis tala presentan múltiples evidencias de uso como combustible (Pochettino et al. 2016; Doumecq 2019). Las hojas y corteza de Jodina rhombifolia son utilizadas como medicinales, y su tallo en la fabricación de una variedad de instrumentos (Hieronymus 1882; Pchettino et al. 2016). Según la creencia popular, este arbusto es protector, no le afectarían las inclemencias del tiempo como sequías, vientos o rayos, y el comportamiento de sus hojas anuncia la presencia de lluvias (Pochettino et al. 2016). Los órganos foliares de Manihot grahami (falso café) son tóxicos en estado fresco (Freire et al. 2005; Martínez Crovetto 2012), pero en infusión son utilizados como medicina (Keller 2007, 2008; Martínez Crovetto 2012), y su madera blanda es buena iniciadora de fuego (Doumecq 2019).
A continuación, presentamos los rasgos y elementos diagnósticos resultantes del análisis del material fresco seguido de la caracterización de rasgos resultantes de distintas actividades de procesamiento:
Celtis tala Gillies ex Planch (Celtidaceae), “tala”: posee frutos cuyo endocarpo contiene células pétreas rectangulares y circulares (Figura 3a) (Lema et al. 2010). Las hojas presentan cistolitos silicificados con valor diagnóstico que se desarrollan por debajo de pelos o células prominentes y pueden contener grandes cristales romboidales (Figura 3b). Estas estructuras foliares fueron descriptas anteriormente por Fernández Honaine et al. (2005).
Jodina rhombifolia (Hook. & Arn.) Reissek (Santalaceae), “sombra de toro”: la semilla presenta células pétreas de formas irregulares (Figura 3c), campanuliformes, ovoides, subrectangulares y rectangulares, así como cristales en forma de prisma de base cuadrada (Lema et al. 2010).
Zea mays L. (Poaceae), “maíz”: posee en sus semillas principalmente granos de almidón simples, poliédricos, con facetas de presión, tamaño variable de 2 a 35 µm; hilum céntrico en forma de V, como un punto o línea; cruz central definida. Pueden aparecer en agregados de dos a más individuos (Winton y Winton 1932; Korstanje y Babot 2005; entre otros).
Tabla 1: Información obtenida de fuentes documentales sobre especies que fueron incluidas en la colección de referencia. Se consigna su denominación según el nombre registrado en las fuentes, la identificación taxonómica asignada en las fuentes, el órgano utilizado, su procesamiento y prácticas asociadas, los autores y la región en la que hacen mención de las plantas. En los casos en que no se especifica en las fuentes analizadas y los datos brindados permiten hacerlo, señalamos una posible identificación taxonómica con un asterisco (*).

Figura 3: Resultados de la colección de referencia: A células pétreas de C. tala; B señalamos cistolito foliar de C. tala; C célula pétrea de J. rhombifolia; D-D´ almidones de Z. mays bajo luz normal (D) y polarizada (D´). E-E´ almidones de M. esculenta con bajo luz normal (E) y polarizada (E´). F-G´ almidones de M. esculenta luego de fermentación bajo luz normal (F y G) y polarizada (F´ y G´), se señalan alteraciones resultados de fermentación (Fig. A y C: colección LEBA; Fig. D: colección Laboratorio 129). Escala 20 µm.
Manihot esculenta Crantz. (Euphorbiaceae), “mandioca”: los granos de almidón de estas raíces son simples o compuestos, en forma de campana, circulares u ovales. El tamaño varía entre 6 hasta 35 µm de longitud. El hilum usualmente es central, y sobre él puede haber una fisura en forma de línea transversal, en Y o en cruz. La cruz de extinción generalmente es central con brazos rectos y, en menor frecuencia, levemente excéntrica con brazos ondulados. Son comunes las facetas de presión en almidones truncados y en circulares. Granos grandes y pequeños pueden presentarse en agregados (Winton y Winton 1932; Pagán Jiménez 2015).
Estudios experimentales sugieren la aparición de distintos rasgos de valor diagnóstico (i.e. fisuras, grietas, perdida de birrefringencia, hidrólisis, entre otros) que dan cuenta de diferentes procesos culinarios, que varían en intensidad según los tiempos y los pasos involucrados, así como dependiendo de las plantas que intervienen. Los resultados de distintas formas de procesamiento de Zea mays fueron publicados por distintos autores (Babot 2003; Pagán Jiménez 2013; Petrucci y Lema 2016; entre otros). El tostado del maíz genera una combinación de partículas de carbón, y almidones intactos junto a otros con pérdida de birrefringencia, con deformación de la cruz de extinción, así como gelatinizados. Se observa en muchos almidones un centro oscuro en la zona del hilum, observable bajo luz normal y polarizada (Babot 2003; Petrucci y Lema 2016). Asimismo, el hervido genera un incremento en el tamaño de los granos y pérdida parcial o total de la birrefringencia. De este tipo de procesamiento también resulta un orificio circular sobre el hilum, que se presenta como un centro oscuro bajo luz normal y polarizada (Petrucci y Lema 2016).
En cuanto a los resultados del proceso de fermentación sobre raíces de M. esculenta, Chandler-Ezell y equipo (2006) registraron una combinación de almidones alterados, agregados y gelatinizados. La fermentación de harina de mandioca en nuestras experimentaciones generó además alteraciones en la cruz de extinción (Figura 3e), orificios en la zona del hilum que, en algunos casos resultó en granos de almidón con forma anular; pérdida de birrefringencia; fragmentaciones y fisuras pudiendo dividir los granos en dos o más partes. Observamos también alteraciones en el contorno y superficie en grado bajo, medio y alto asociado a una baja visibilidad al MO de los granos (Figura 3f).
Material arqueológico
El análisis de las muestras recuperadas en los fragmentos cerámicos de Las Marías nos permitió observar diversas estructuras vegetales, así como fragmentos de organismos procariotas y animales. En la Tabla 2 (ver Material Anexo) sintetizamos los resultados obtenidos en los dieciséis fragmentos analizados separados en tres grandes grupos: contenedores de procesamiento y/o consumo, contenedores de almacenaje e instrumentos de transferencia.
Contenedores de procesamiento y/o servicio
Todas las muestras recuperadas de este grupo morfo-funcional contenían microrrestos, ya sean vegetales como protistas y/o animales (Figura 4a). Las estructuras vegetales registradas incluyen fibras de distintos tipos: (1) con lumen ancho en relación a las paredes y extremos aguzados; (2) planas, con torsiones sobre su eje, lumen estrecho en relación a la pared y extremos aguzados; (3) con lumen estrecho en relación a las paredes anchas de extremos redondeados, con saliencias a modo de mamelones.
Hallamos fitolitos de Celtis tala y otros afines a distintos grupos vegetales (Danthonioides, Panicoides, Arecoides y Festucoides, ver Figura 4b). Estos elementos silíceos se encuentran tanto en la superficie interna como externa de los contenedores, pero cabe resaltar que fueron mayoritarios, en cantidad y tipos morfológicos, en los fragmentos cerámicos del grupo M (muestras control). Registramos distintos morfotipos de esclereidas, principalmente células pétreas o braquiesclereidas y macroesclereidas (Crang et al. 2018), algunas de las cuales corresponden a Celtis tala (Figura 4g) y Jodina rhombifolia (Figura 4h). Observamos también tricomas estrellados (Figura 4i) y un grano de polen (Figura 4j).
En seis contenedores hallamos granos de almidón, los cuales se recuperaron principalmente de las superficies internas. Observamos almidones poliédricos que denominamos Tipo 1 y asignamos a Zea mays. Algunos presentaron centros con proyecciones oscuras en la zona del hilum asociados a microcarbones (Figura 4k y 4l´). Otro conjunto de almidones estuvo representado por granos simples, con forma de campana. Muchos de estos gránulos no polarizaban, o presentaban una cruz de extinción incompleta. Estos almidones, que resultaron indeterminados, los llamamos Tipo 2 (Figura 4l y 4l´).
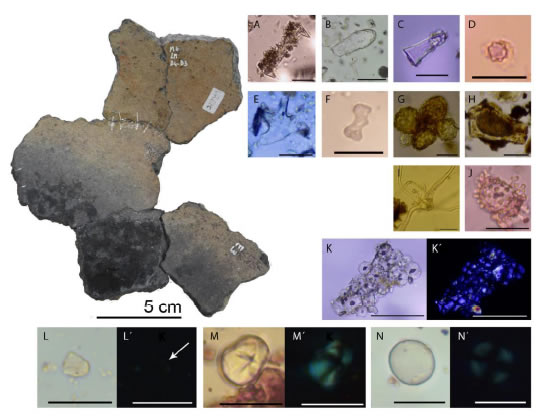
Figura 4: Izquierda-superior reconstrucción de base y cuerpo de un contenedor de procesamiento. A espícula de espongiario; B-F fitolitos: B cistolito de C. tala; C Danthonioide; D Arecoide; E Asteriforme; F halterio polilobulado; G células pétreas de C. tala; H célula pétrea de J. rhombifolia; I tricoma estrellado; J grano de polen; K almidones Tipo 1 de Z. mayz; L almidón Tipo 2 indeterminado y L´ polarización, indicamos con una flecha el almidón con pérdida de birrefringencia; M almidón Tipo 3 aff. M. esculenta; N almidón Tipo 4 indeterminable. Escala de microrrestos 20 µm.
El tercer tipo de almidones que registramos consistió en granos simples, ovoides, con una fisura horizontal o en “Y” en la zona del hilio y una cruz de extinción levemente excéntrica con signos de alteración, como brazos muy anchos y difusos (Figura 4m y 4m´). Los almidones Tipo 3 resultan afines a uno de los morfotipos característicos de Manihot esculenta. Además, el cuarto tipo de granos de almidón estuvo representado por elementos simples y esféricos a elipsoides, con cruz de extinción central (Figura 4n y 4n´). Los almidones Tipo 4 son ubicuos en el reino vegetal, por lo que son indeterminables.
Contenedores de almacenaje
En este tipo de contenedores registramos espículas de espongiarios (Figura 5a) y estomatocistes de algas Chrysophyceae en las superficies internas. Observamos también elementos indeterminables, como estructuras planas perforadas de posible origen animal (Figura 5b) y masas rojizas que en algunas ocasiones brillan bajo luz polarizada. El componente fitolítico se compone de elementos diagnósticos para Danthonioide, Panicoide y Festucoide (Figura 5c y 5e).
Observamos granos de almidón correspondientes a los Tipos 1 (Figura 5f y 5f´), 3 y 4 mencionados antes. Asimismo, registramos una nueva categoría de grano de almidón que denominamos Tipo 5. Se encuentran en forma individual o en agregados. La mayor parte presenta contornos y superficies muy degradados, asociados a fisuras que se proyectan desde el hilum. Las formas varían entre ovales, con tamaños similares al Tipo 3, y campanas, en su mayoría unidas por su base (Figura 5g y 5g´). La cruz de extinción levemente excéntrica por lo general está alterada, presentándose engrosada, difusa, y en algunos casos con múltiples brazos o brazos con torceduras (Figura 5h y 5h´). La birrefringencia bajo luz polarizada es media a baja y prima una baja visibilidad bajo luz normal, lo que dificultó su visualización. Considerados en conjunto, los Tipos 3 y 5 de almidones resultan afines a Manihot esculenta. Asimismo, los patrones de desgaste y alteración presentes en el Tipo 5 podrían corresponderse con actividades de fermentación observadas en la colección de referencia.

Figura 5: Izquierda parte superior de un contenedor de almacenaje; A espículas de espongiarios; B elemento indeterminable; C-E fitolitos: C Danthonoide, D Panicoide y E Festucoide; F-F´ almidón Tipo 1 de Z. mays; G-H almidones Tipo 5 aff. M. esculenta: G-G´ campanas, H-H´ señalamos cruz de extinción con brazos con modificaciones. Escala de microrrestos 20 µm.
Instrumentos de transferencia
Registramos espículas de espongiarios y estomatocistes de algas Chrysophyceae (Figura 6 A). En tanto el componente fitolítico, registramos morfologías no diagnósticas y otras de afinidad Danthonioide (Figura 6b y 6c). Observamos también almidones Tipo 1 (Zea mays), tanto agregados como individuales (Figura 6d y 6d´), así como otros que reúnen las características Tipo 3 y 5 (afines a M. esculenta;Figura 6e y 6f´); y Tipo 4. Por último, al igual que en las muestras cerámicas anteriores, registramos fragmentos de tejido vegetal y masas de color amarillento a rojizo, que en algunos casos son brillantes bajo luz polarizada y resultan indeterminables.
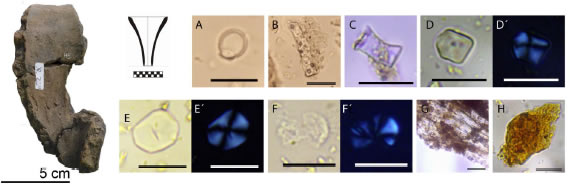
Figura 6: Izquierda fragmento de instrumento de transferencia y su reconstrucción digital; A estomatociste de alga Chrysophyceae, B fitolito elongado con superficie agujereada, C fitolito Danthonioide; D-D´almidón Tipo 1 de Z. mays, E-F´ almidones aff. M. esculenta: E-E´ Tipo 3; F-F´ Tipo 5; G fragmento de tejido; H masa rojiza indeterminable. Escala de microrrestos 20 µm.
Reflexiones y consideraciones finales
El análisis realizado nos permitió observar la presencia de microrrestos vegetales en todas las muestras correspondientes al conjunto cerámico del sitio arqueológico Las Marías. Así también, pusimos a prueba estrategias metodológicas que resultaron satisfactorias para el caso de estudio. Registramos una diversidad de estructuras vegetales, correspondientes tanto a plantas silvestres como domesticadas con distintas formas de procesamiento.
Desde el punto de vista metodológico, la selección de muestras control nos permitió apreciar que aquellos fragmentos cerámicos que no fueron expuestos al lavado y acondicionamiento registran mayor cantidad y diversidad de estructuras vegetales. Estas estrategias nos permiten ajustar los protocolos de trabajo en laboratorio. Por otro lado, el uso de ácido láctico para decolorar costras quemadas adheridas a las superficies cerámicas facilitó la observación de elementos con buena preservación y su posterior determinación. La implementación de este procedimiento nos concedió subsanar estas dificultades y por ende constituyó un aporte metodológico significativo.
Para aproximarnos a los contextos de uso y prácticas asociadas por parte de los grupos que habitaron la zona articulamos la información procedente del análisis de microrrestos vegetales, con las fuentes documentales, la colección de referencia y los estudios cerámicos realizados. Destacamos que las fuentes documentales no sólo son herramientas para la formulación de hipótesis acerca del pasado, sino para realizar interpretaciones a partir de confrontar nuestras realidades con otras que nos permitan pensar en una variedad de situaciones o mundos posibles (Haber 2011). El relevamiento de crónicas y diarios del siglo XVI al XIX, así como de publicaciones etnobotánicas recientes, en asociación con el análisis paleoetnobotánico nos permitió incorporar a la colección de referencia plantas tanto silvestres de la región rioplatense-paranaense, como otras domesticadas de circulación macrorregional (como mandioca o maíz). Observamos la mención de distintos órganos almacenadores de almidón potencialmente útiles y asociados a diversas prácticas, ya sea alimenticias, medicinales o de intercambios sociales. En este sentido, cabe aclarar la importancia que reviste el análisis contextual de los documentos dado que permite plantear múltiples preguntas e interpretaciones. Tal es el caso del diario de Lopes de Sousa [1530-1532], donde la ausencia de menciones sobre vegetales domesticados vinculados a la descripción del modo de vida de poblaciones nativas se transforma en un elemento significativo para confrontar con otros relatos de la misma época en esa región. Una instancia para preguntarnos sobre los intereses que intervinieron, la abundancia estacional de recursos, el tipo de contacto que realizaron con poblaciones nativas, entre otros. Por otro lado, en fuentes documentales tempranas registramos para situaciones críticas el uso alimenticio de plantas posiblemente silvestres (Ramirez [1528]; Lopez de Sousa [1530-1532]; Schmidel [1599]). Por lo general, dichas plantas no se asocian explícitamente al conocimiento botánico de poblaciones nativas (con excepción de algunos casos en Schmidel [1599]), aunque no debemos descartarlo, ya que todas las expediciones tenían integrantes indígenas que hacían de intérpretes y guías.
El análisis arqueobotánico evidenció microrrestos vegetales en los tres tipos morfo-funcionales, en algunos casos granos de almidón con rasgos que demuestran distintos tipos de procesamiento para su aprovechamiento. Los mismos también se recuperaron en bordes externos de las tres categorías alfareras: en cuellos curvos de contenedores de almacenaje, en extremos afinados de instrumentos de transferencia y en incisiones y superficies lisas cercanas al borde de contenedores de procesamiento y/o servicio. Consideramos que esto podría vincularse con los usos dados a los objetos y con la presencia de zonas que permitan la acumulación, entrampe y preservación de las estructuras vegetales. Asimismo, las espículas de espongiarios localizadas principalmente en superficies internas podrían relacionarse con la presencia de agua en las sustancias que se procesaron, almacenaron o manipularon.
Muchas estructuras registradas, entre ellas algunos granos de almidón con patrones diagnósticos, no resultaron compatibles con las muestras de referencia, de este modo los asignamos por el momento a la categoría “indeterminados”. En consecuencia, consideramos que es indispensable seguir enriqueciendo la colección comparativa con materiales frescos y sometidos a distintos tipos de procesamientos, tanto para ajustar las determinaciones taxonómicas, como para aproximarnos de forma integral a los saberes botánicos de las poblaciones humanas del pasado.
Investigaciones previas de abordaje morfo-funcional del registro cerámico del sitio arqueológico Las Marías (ie. Paleo y Pérez Meroni 2008) encuentran en el presente trabajo una evidencia directa sobre los contenidos vegetales y tipos de procesamientos realizados sobre ellos. Podemos observar que los contenedores de almacenaje e instrumentos de transferencia se asocian al manejo de sustancias líquidas, ya sea fermentadas o maceradas, infusiones, entre otras; así como a la manipulación, almacenamiento o trasvaso de sustancias molidas como harinas. En tanto, los contenedores de procesamiento y/o servicio presentan evidencias de exposición al calor, que podrían corresponderse con la preparación de hervidos, asados, horneados o tostados.
Los resultados de las muestras analizadas revelan las primeras evidencias directas de la presencia y procesamiento de plantas domesticadas para la zona. Es importante destacar que estas se encuentran asociadas con otras de crecimiento espontáneo y que resultan características del bosque de tala, como Celtis tala y Jodina rhombifolia.
Los almidones de Zea mays se asocian con rasgos diagnósticos vinculados con la exposición al calor pudiéndose tratar de tostado, horneado o hervido. Estudios previos sobre restos orgánicos de tiestos cerámicos procedentes de este sitio realizados mediante cromatografía gaseosa con espectrómetro de masa, dieron como resultado que podrían corresponder a la cocción de peces o a maíz dulce (Paleo y Pérez Meroni, 2007; Pérez Meroni et al. 2010). En esa oportunidad, no se contaba con evidencias que avalen la utilización del maíz por parte de estos grupos. A partir del análisis de microrrestos aquí presentados podemos realizar una nueva lectura y sustentar con otra línea de evidencias la utilización de plantas domesticadas como Zea mays y posiblemente Manihhot esculenta. Los microrrestos asignados como afines a M. esculenta presentan características congruentes con prácticas de fermentación, maceración, u otras donde se produce la hidrólisis de los granos por acciones enzimáticas. Estas prácticas son compatibles con la confección de vinos y otras bebidas a partir de Manihot sp. registradas en crónicas tempranas (Schmidel [1534-1554]). En la zona de estudio crece Manihot grahamii Hook (o falso café). Este árbol no cuenta con estructuras de almacenamiento subterráneas como M. esculenta, y no hemos registrado el uso de sus raíces en fuentes documentales. Probablemente su vínculo con las poblaciones humanas se basó en el consumo de hojas con fines medicinales, la obtención de leña y sombra (Keller 2007, 2008; Martínez Crovetto 2012; Doumecq 2019).
En concordancia con el registro arqueológico, Celtis tala y Jodina rhombifolia presentan registros de uso en fuentes documentales tanto de sus frutos como hojas. Las células pétreas y cistolitos foliares descriptos en este trabajo se hallan en su totalidad en fragmentos de contenedores de procesamiento y/o servicio. Asimismo, se encuentran mayoritariamente en superficies internas y dentro de costras carbonizadas, así como en bordes o agujeros de suspensión o reparación, localizaciones que consideramos congruentes con el uso de cocción, fermentación, contención, derrame, servido o entrampe en pequeñas cavidades. Estas evidencias se incorporan a otros hallazgos arqueológicos en contextos de procesamiento de frutos de tala, tanto en cerámica como en instrumentos de molienda (Babot et al. 2007; Gonzalez y Frere 2009; Lema et al. 2010; Pérez Meroni et al. 2010), como así también en fogones (Aldazabal y Vazquez 2015). Oportunamente, al indagar sobre prácticas alimenticias asociadas a vegetales en el sitio Las Marías se propuso el uso de C. tala y J. rhombifolia, y se consideró que su procesamiento estaría vinculado a actividades de tostado y molienda implementadas para facilitar la cocción de frutos y semillas con tejidos ricos en almidón (Lema et al. 2010; Pérez Meroni et al. 2010). Por otro lado, también existen registros antracológicos (Ghiani Echenique et al. 2020) y etnobotánicos (Doumecq 2019) del uso de estos árboles como combustible en la zona.
Los trabajos realizados hasta el momento evidencian que la diversidad de modos en que se relacionan las poblaciones locales con estas plantas de crecimiento espontáneo sugieren la presencia de distintas formas de manejo del entorno (Casas 2001). Este conjunto de actividades humanas diversas de control del ambiente tendientes a la preservación y proliferación de alguna/s población/es vegetal/es pueden considerarse como prácticas de cultivo en su sentido amplio (Casas 2001; Lema 2010). De esta forma, Jodina rhombifolia y Celtis tala podrían haber participado en actividades de protección, fomento y/o recolección que aseguren su presencia e involucramiento en distintas actividades (desde alimentos, medicinas, tinturas, hasta sombra, protección y reparo, creencias populares, entre otras). Las plantas domesticadas, aquellas que necesitan de la intervención humana para su supervivencia (sensu Ford 1979; Lema 2010), podrían haberse introducido a través de interacciones sociales con áreas vecinas (es el caso de Zea mays y posiblemente Manihot esculenta). Esta propuesta deberá ser complementada con análisis de sedimentos para esclarecer la probabilidad de actividades hortícolas en el seno de estas poblaciones ribereñas.
En este trabajo utilizamos categorías biológicas (como domesticada, cultivo, silvestre o espontánea) y económicas (como uso, recurso), y mantenemos la división entre seres humanos y plantas con fines prácticos y analíticos teniendo en cuenta sus limitaciones y connotaciones. Estas reflexiones nos permiten superar dicotomías como silvestre/domesticado y cazador-recolector/agricultor, y pensar una población con patrones de movilidad relativamente bajos que practique actividades de caza, pesca y recolección de una amplia gama de recursos, incluidas distintas técnicas de cultivo de vegetales, como parte de una diversidad de formas de manejo del entorno natural-cultural.
Agradecimientos
Esta contribución fue realizada con subsidios provenientes de UNLP y PICT, así como una beca doctoral CONICET. Agradecemos a lxs evaluadores anónimxs y a la Dra. Day Piraría por la lectura detenida y sus comentarios que enriquecieron este manuscrito, y a las Dras. Lema, Petrucci y Capparelli por facilitarnos el acceso digital a las colecciones de referencia del LEBA (FCNyM) y del Laboratorio 129 (Div. de Arqueología del Museo de Ciencias Naturales de La Plata) en tiempos de COVID-19. A dichos laboratorios pertenecen algunas imágenes correspondientes a la colección de referencia que publicamos en este trabajo.

