











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Rhea americana (ñandú) es un ave exclusiva de Sudamérica,se encuentran dentro del orden de los Rheiformes y la familia Rheidae, que incluye al suri (Rhea tarapacensis), al choique (Rhea pennata), y al ñandú común (Rhea americana). A esta última corresponden cinco subespecies: R. a. americana, que se distribuye en el norte y este de Brasil; R. a. intermedia, que habita el sur de Brasil y Uruguay; R. a. nobilis, en el este de Paraguay; R. a. araneipes en el este de Bolivia y sudeste de Brasil y Paraguay; y R. a. albescens, que cuenta con una amplia distribución fundamentalmente en el norte y centro-este de Argentina (Martella et al. 2018).
En la actualidad, en nuestro país,R. a. albescens 1 se encuentra presente en las ecorregiones del Chaco seco y Chaco húmedo, Monte, Espinal, Pampa, Esteros del Iberá y Campos y Malezales (Gasparri 2016). Es un ave no voladora y constituye la de mayor tamaño del continente (López Lanús et al. 2008). De acuerdo con información etnográfica y etnológica, el ñandú fue aprovechado por distintos grupos humanos que habitaron la región de estudio en el pasado (Miraglia 1975¸Paucke 1942 [1749-1767]), mientras que actualmente constituye una “especie etnobiológica clave” (sensu Medrano y Rosso 2016) para algunos grupos del Gran Chaco. Es decir, que, en virtud de su utilidad y valor simbólico, se torna ecológicamente crucial para el mantenimiento del ambiente antropogénico, su sistema de subsistencia y forma de vida (Medrano y Rosso 2016).
Los primeros trabajos arqueológicos que se centran en el registro de reídos en Argentina comenzaron a desarrollarse para sitios de la Patagonia, hacia la década de 1980 (Belardi 1999; Caviglia y Figuerero Torres 1976; Fernández 2000;Miotti y Salemme 1999; Tambussi y Tonni 1985; Salemme y Miotti 1998). Estos estudios señalan una discrepancia entre la información que mencionan las crónicas consultadas por los autores, -en las cuales se hace referencia al amplio uso que le daban los grupos prehispánicos a estas aves-, y lo observado en el registro arqueológico (Giardina 2010, 2018). Posteriormente, comenzó a profundizarse el rol de los reídos dentro de las prácticas de subsistencia también en otras regiones (Acosta 2005; Álvarez 2015; Bonomo et al. 2008; Cornaglia Fernández 2014; Del Papa y Moro 2017; Fernández 2010; Frontini y Picasso 2010; Giardina 2018; Medina et al. 2011; Pérez Jimeno y Del Papa 2016; Pérez Jimeno y Servin 2017; Quintana y Mazzanti 2001; Salemme y Frontini 2011; Santini 2009, entre otros). La variedad de tópicos abordados en los últimos años ha sido amplia, abarcando temas diversos, como el uso de la información etnohistórica, el rendimiento económico de la especie, la posible determinación específica a partir de cáscaras de huevos y estudios de movilidad a partir de valores de isotopos estables. Además, se han realizado trabajos, que buscan explicar la representación anatómica de los conjuntos, relacionados con los agentes-procesos que intervienen en su preservación (e.g. Belardi 1999; Cruz 2015; Cruz y Elkin 2003; Cruz y Muñoz 2020; Fernández 2000; Frontini y Picasso 2010).
Diversos investigadores señalan que en los ambientes ribereños del sistema Paraná-Paraguay la frecuencia de Rhea americana en los conjuntos arqueofaunísticos, suele ser escasa -o estar ausente en ciertos casos, observando la misma discrepancia con las fuentes documentales antes referida (Barboza y Martín 2014; Bonomo et al. 2010, 2016; Del Papa et al. 2019; Feuillet Terzhaghi y Escudero 2011; Ottalagano 2016, 2019; Ottalagano et al. 2015; Pérez Jimeno 2007; Pérez Jimeno y Servin 2017; Píccoli et al. 2017; Rodríguez y Bonomo 2021; Santini 2009, 2013; Santini y De Santis 2011; Sartori 2013; Sartori y Colasurdo 2011; Sartori y Pérez Jimeno 2012; Sartori et al. 2017). Si bien, los análisis zooarqueológicos en esta zona se han incrementado notablemente en los últimos años, generando nueva información de grano fino provechosa para realizar estudios comparativos, la representación del ñandú en dichos conjuntos hasta ahora no había sido estudiada con una perspectiva macroregional. Por eso resultó de interés evaluar, a partir de la información bibliográfica disponible, la presencia de R. americana para establecer la variabilidad que existe en los sitios del Holoceno tardío emplazados en sectores vinculados a los ambientes ribereños del sistema Paraná-Paraguay. Para ello, se analizó comparativamente la abundancia relativa de R. americana en los registros zooarqueológicos de 25 sitios (Figura 1), su representación esqueletal y las evidencias de procesamiento antrópico. El objetivo es evaluar su representación en los conjuntos y realizar inferencias regionales en relación a su explotación, lo que permite comprender mejor el rol de esta especie dentro de las estrategias de subsistencia de las poblaciones humanas (ver Tabla en Anexo). Se considera que su disponibilidad en el Chaco Húmedo y en las proximidades de los ambientes propios de la llanura aluvial del río Paraná, así como su gran tamaño además de la posibilidad de explotar productos secundarios (e.g., plumas, huesos, uñas, tendones, etc.), la habrían tornado un recurso atractivo para los diferentes grupos cazadores-recolectores-pescadores del pasado.
Marco ambiental
Los sitios arqueológicos considerados en este trabajo se localizan, zoogegráficamente, dentro de la sub-región Guayano-brasileña del Neotrópico (Ringuelet 2004). Con fines operativos se subdividió el área estudiada, agrupando los sitios según el lugar de emplazamiento, en tres sectores: el primero en el Chaco Húmedo (CHH), el segundo en una zona aledaña a ésta, en un sector de transición entre el CHH y la ecorregión Delta e Islas del Paraná (LLA-a) (Burkartet al.1999; Parera 2002), mientras que el tercero, se localiza entre esta última ecorregión y la zona de transición con el Espinal y la Pampa (LLA-b). La mayoría de los sitios que se localizan en los dos últimos sectores se emplazan en el Paraná medio y unos pocos, donde comienza el Paraná inferior (Figura 1). A continuación, se describen las características ambientales de esas ecorregio- nes siguiendo el orden de norte a sur.
Los sitios del sector más septentrional se encuentran emplazados en el Chaco Húmedo (CHH) (Burkart et al. 1999; Parera 2002), en un sector denominado como Chaqueño Ribereño del sistema Paraná-Paraguay, en donde se desarrollan grandes humedales (Del Papa y Lamenza 2019; Neiff et al. 1994). La zona se caracteriza por el desarrollo de bosques sobre albardones y mosaicos de vegetación en los interfluvios: bosques bajos, pastizales, pajonales y palmares. Es una de las zonas con más biodiversidad de la Argentina y alberga una amplia cantidad de mamíferos, reptiles y peces. Entre las aves los ejemplares más característicos son el ñandú (Rhea americana), la pava de monte (Penelope obscura) y el tataupa (Crypterellus tataupa), entre otras.
La mayor parte de los sitios se emplazan en la ecorregión Delta e Islas del Paraná, que incluye el corredor fluvial y las planicies aluviales del tramo inferior del río Paraguay, de los tramos medio e inferior del río Paraná y el cauce del Río de la Plata (Bó 2005; Burkart et al. 1999). El sector considerado abarca sólo el tramo medio del río Paraná, que se caracteriza por un conjunto de macro sistemas de humedales de origen fluvial que se desarrollan encajonados en una gran falla geológica, a lo largo de la llanura chaco-pampeana. Si bien conforma una unidad ecoregional -en la medida en que comparte la fauna íctica y la mayoría de los mamíferos-, existen variaciones en la distribución-abundancia faunística de norte a sur, por eso se distinguen aquí dos sectores (LLA-a y LLA-b). La alta productividad ambiental se encuentra regulada por los pulsos de inundación del río Paraná que ejerce un efecto climático que se traduce en la presencia de una cuña del Dominio Amazónico, el cual permite la penetración de poblaciones animales típicamente amazónicas hacia el sur (Bó y Malvárez 1999; Cabrera 1971; Neiff 1999; Ringuelet y Aramburu 1957). De esta forma, la fauna característica de toda la zona incluye especies subtropicales adaptadas al ámbito fluvial (Gobierno de la Provincia de Santa Fe -MAGIC- y Administración de Parques Nacionales 1997). Otro aspecto característico de todos los sectores es la alta concentración de peces, fundamentalmente de los órdenes Siluriformes y Characiformes (Ringuelet 2004). La variedad de aves excede las 330 especies (De la Peña 1976; Gobierno de la Provincia de Santa Fe -MAGIC- y Administración de Parques Nacionales 1997) y está representada tanto por aves rapaces, como especies de hábitos acuáticos, garzas -Mycticorax obscurus, Syrigma sibilatrix-, biguá -Phalacrocorax brasilianus- y terrícolas como pavas de monte Penelope sp.- y ñandú -Rhea americana- (Gasparri 2016).
Por último, algunos de los sitios se encuentran en las ecorregiones del Espinal y Pampa (Burkart et al. 1999), donde se encuentran las especies animales pampeanas típicas, pero también mamíferos de hábitos acuáticos. Las aves que prevalecen pertenecen a familias que poseen hábitos acuáticos, tales como Ciconiidae (cigüeñas), Ardeidae (garzas) yAnatidae (patos) (Manzi y Gallardo 1970). Otras especies características de ambientes no anegados son Rhea americana, el leñatero -Anumbius annumbi-, los espineros -Phacellodomus sp.-, los pijuíes -Synallaxis sp.- y lechuzones del campo -Asio flammeus-. A su vez, en este mismo sector, pero del lado entrerriano, la fauna se caracteriza por las especies pampeanas mencionadas que poseen cierta plasticidad adaptativa (Gollán 1958).
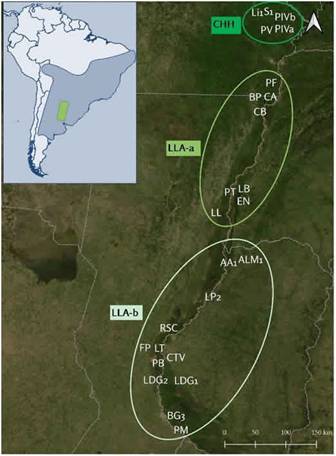
Figura 1: itios arqueológicos considerados en este trabajo y los agrupamientos realizados CHH; LL-a; LL-b. En celeste, mapa de distribución de Rhea americana (Gasparri 2016: 27). Referencias: Sotelo 1-S1-,La Ilusión 1 -Li1- Potrero IV a y b -PIVa, PIVb-, Potrero V -PV-, Puesto Fantin -PF-, Cerro Aguará -CA-, Barrancas del Paranacito -BP-, Campo Binaghi -CB-, Los bananos -LB-, Paso del Tala -PT-, El nene -EN-, La lechuza -Ll-, Arroyo Arenal 1-AA1-, Las Palmeras -LP2-, Arroyo Las Mulas1 -ALM1-, Río Salado Coronda -RSC-, Las Tejas -LT-, Pajas Blancas -PB-, Familia Primón -FP-, Co. Tapera Vázquez -CTV-, Laguna de los gansos 1 y 2 -LDG1 y LDG2-, Bajada Guereño -BG3-, Playa Mansa -PM-.
Etología de Rhea americana
El ñandú (Rhea americana) constituye una de las tres especies del orden Rheiformes (Reboreda y Fernández 2005). Es un ave no voladora de gran tamaño, endémica de la Región Neotropical, que en Argentina se distribuye desde el norte, hacia el centro-este y llega en el sur hasta Rio Negro. Habita ambientes abiertos, provistos de pastos o matorrales, donde puede ocultarse de los depredadores. Se caracteriza por tener alas poco desarrolladas y por ser muy rápida en la carrera. Alcanza el tamaño adulto recién al año de vida, y llega a medir 1,50 m de altura y un peso de 40 kg (Fernández 1998).
Es una especie gregaria que forma bandadas de entre 3 y hasta 50 individuos. En la época reproductiva, las hembras depositan los huevos en un único nido, que es una depresión en el suelo oculto entre la vegetación (De la Peña 1976). El macho incuba los huevos y cuida los pichones luego de la eclosión. Si bien habita ambientes abiertos, cuando se halla con crías prefiere ubicarse en cercanías de cuerpos de agua. Posee hábitos diurnos, dedicando la mayor parte del tiempo a su alimentación, constituida primordialmente por vegetales (Gasparri 2016).
Constituye uno de los recursos animales autóctonos de mayor tamaño en la región pampeana Argentina. Un individuo de 1 año de edad y 25 kg de peso vivo, rinde alrededor del 65 % de dicho peso en carne, el 7 % en grasa y un 13% en hueso. Permite la obtención de unos nueve o 10 Kg de carne, vísceras, entre uno y dos kg de grasa, 0,5 m2 de cuero, 0,4 kg de plumas y huesos (Giardina 2006; Navarro y Martella 2016).
Si bien es posible que Rhea americana haya estado disponible en toda el área que abarca el presente trabajo, de acuerdo a las características ambientales descriptas en el apartado anterior y su etología, las condiciones más propicias para el desarrollo de la misma, se darían en el Chaco húmedo, el Espinal y la Pampa. No obstante, en la época de nidación y crías podrían encontrarse también en los
humedales y delta del Paraná, ya que como se refiere en el párrafo precedente, ocultan sus nidos entre la vegetación y prefieren la proximidad a fuentes de agua cuando se halla con crías (Gasparri 2016).
Información etnohistórica y etnográfica sobre la relación de los grupos originarios con R. americana
Los datos etnohistóricos considerados refieren a documentos escritos por cronistas que estuvieron en el nuevo continente durante los S XVI y XVIII en el área de estudio y en otras aledañas a ésta (Paucke 1942 [1749-1767]; Schmidel 2003 [1564]). Estas crónicas otorgan información acerca de la subsistencia de grupos del Gran Chaco y la Planicie de inundación del río Paraná. Por otra parte, también se hace referencia a investigaciones etnográficas sobre grupos de momentos históricos y actuales que provienen de trabajos de campo realizados en diferentes comunidades (Arenas y Porini 2009; Medrano y Rosso 2016; Metraux 1937; Miraglia 1975; Palavecino 1933). Se hizo hincapié en aquellos grupos humanos cuyo tipo de subsistencia es comparable con la de quienes habitaron el área en el pasado pre-europeo; y, si bien los objetivos de cada uno de los autores de dichos documentos son diferentes y el grado de detalle que brindan es muy disímil, el análisis de dicha información proporciona marcos de referencia más amplios para las interpretaciones arqueológicas que aquí se realizan.
Entre los primeros documentos que se cuentan para la zona figuran los escritos de Schmidel (2003 [1564]). En su viaje observó que algunos grupos originarios asentados en la actual Provincia de Santa Fe, como los Chaná, Timbú y Mepenes, con los cuales tuvo contacto en su viaje por el río Paraná en el Siglo XVI, consumían carne de ñandú. “De allí navegamos el Paraná arriba y después de 4 días de viaje llegamos a una nación que se llama Zeckennaus Saluaischco (Chaná-Salvajes), [son] gente petiza y gruesa, no tienen más de comer que pescado y miel. […] están de guerra con los Machueradeiss; y su carne es la de ciervos, chanchos del monte, avestruces y conejos” (Schmidel 2003 [1564]: 167).
Por su parte, Paucke (1942 [1749-1767]), misionero jesuita que convivió con los Mocovíes en las reducciones San Javier y San Pedro en la provincia de Santa Fe, entre 1752 y 1767, detalla cómo ellos cazaban el ñandú a caballo y con boleadora, e igualmente describe la dificultad que la misma acarreaba: “Los indios los cazan de a caballo y cuatro o cinco indios tienen siempre que trabajar para matar un avestruz; no les sería tampoco posible de conseguirlo si a los veinte o treinta pasos no recurrieran a las boleadoras o le rompieran mediante un tiro de la macana ya el pescuezo, ya las patas...” (Paucke 1942 [1749-1767]: 237-238).
Además, este autor menciona el bajo rendimiento de carne que proporciona esta especie. Cuenta, asimismo, que hierven la carne de los muslos y derriten la grasa en un recipiente, además de asar las alas “debajo de ceniza caliente” (Paucke 1942 [1749-1767]: 328-329). También refiere que utilizaban sus huevos, así como los huesos de las patas y “los más gruesos” de las alas para hacer pífanos; las plumas blancas teñidas como adorno y la piel del cuello para hacer sus tabaqueras (Paucke 1942 [1749-1767]). En un estudio realizado sobre los escritos de Paucke, los investigadores Rosso y Pautaso analizan las relaciones de las aves con los grupos Mocovies de las misiones, concluyendo que Rhea americana sería una de las especies más explotadas al registrar en esta especie nueve “categorías de uso”: como alimento, como anunciadores, como parte de la cultura material, como bien de intercambio, para mascotismo, como medicina y para las prácticas chamánicas, astronómicas y rituales (Rosso y Pautasso 2017:151-156). Destacan el papel del ñandú que se encuentra relacionada a los diferentes ámbitos culturales de estos grupos, cosa que no ocurre con las demás aves relevadas por los autores.
Por otro lado, ya en el siglo XX, los trabajos de los investigadores Metraux (1937) y Palavecino (1933) sugieren el uso de plumas para la confección de adornos; asimismo señalan, al igual que Miraglia (1975) entre los Maká del Chaco, el uso de camuflajes o refugios para su captura, o de fuego en zona de pastizales. Según este último autor, la caza de ñandú en momentos históricos se realizaba con arco y flechas, mientras que actualmente es con el fusil. Además, menciona que, en algunos casos, inmediatamente posterior a la captura de estas aves se las trozaba -en cuartos, alas y cuello- para transportarlas en redes (Miraglia 1975).
Más recientemente, entre 1983 y 2004, los biólogos Arenas y Porini (2009) realizaron observaciones respecto de la relación de las comunidades Wichí y Qom con las diferentes especies faunísticas, entre ellas el ñandú, en el oeste de la provincia de Formosa. Al igual que Medrano (2014) quien realizó un estudio etnobiológico en la misma provincia entre 2008 y 2012, señalan que los Qom consumen y almacenan la grasa de ñandú, tanto como alimento como para usos medicinales:
“… Estas posibilidades de acceder a sustancias grasas son bien aprovechadas, pero se manifiesta claramente que la única especie que la tiene en suficiente cantidad como para guardar y almacenar es el “suri” (Rhea americana). En efecto, es tal la cantidad que posee en tiempos de gordura que se la puede guardar en botijos o recipientes adecuados para meses venideros.” (Arenas y Porini 2009: 73).
Así también, las patas del animal constituyen una reserva de carne y grasa para su consumo diferido “se separan de las piernas, se las lleva al fuego, quemándole la piel, y luego las desecan al sol o al calor del fogón. Su interés se circunscribe a momentos de gran carencia de alimentos (…) Esas patas se guardan por unos días y cuando hay necesidad las hierven tomándose un caldo preparado con ellas” (Arenas y Porini 2009:74-75).
También mencionan la utilización de su cuero para confeccionar el faldellín de las mujeres y el taparrabos de los varones, bolsas para guardar diversos objetos, así como recipiente para guardar miel y tabaqueras. Hay referencias, asimismo, de su empleo como parche resonador en el tambor o timbal; además, con sus uñas confeccionaban sonajas (Arenas y Porini 2009). En cuanto al aprovechamiento de los huesos, dan cuenta de que estos grupos suelen utilizar la quilla como plato o bol y a la fíbula como escarificador. Quién se lo clavaba adoptaba la cualidad del animal.
En otro trabajo, en el que también se realiza un estudio etnozoológico entre los Qom de Formosa, las antropólogas Medrano y Rosso (2016) concluyen que ésta no solo es el ave más utilizada y de la cual realizan un aprovechamiento integral, sino que presenta diversas connotaciones en los diferentes ámbitos sociales. Por esto, sostienen que es una "especie etnobiológica clave" (Medrano y Rosso 2016: 61), ya que superan las relaciones meramente utilitarias y la comunidad se relaciona con el ave de diversas maneras como parte de su cosmología.
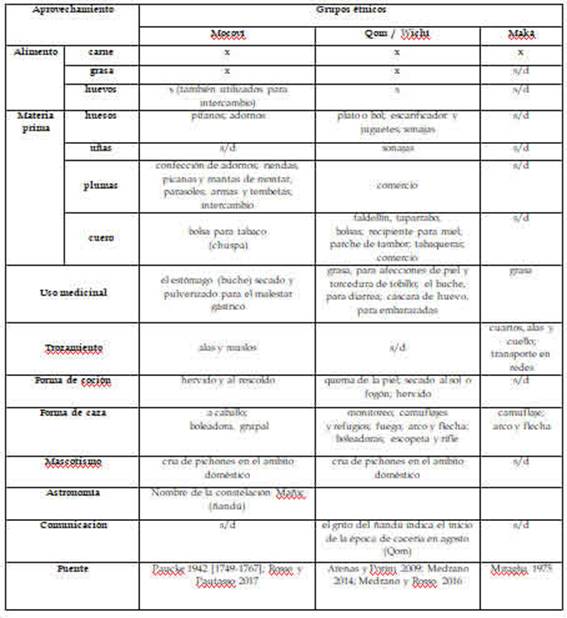
La información proporcionada por cada una de las fuentes consultadas refleja la explotación de este recurso en diferentes comunidades, desde los primeros momentos del contacto hispano-indígena hasta la actualidad. Una síntesis de los diferentes usos mencionados aquí se presenta en la Tabla 1
Metodología
El análisis de los registros de Rhea americana se realiza desde una escala regional. Los datos provienen de 25 conjuntos de diferentes sitios arqueológicos, los cuales fueron agrupados en tres sectores, como se mencionó más arriba (Figura 1). La definición de cada conjunto fue establecida en función de la información publicada por quienes realizaron los estudios zooarqueológicos de cada muestra. Se estableció como requisito que el registro arqueofaunístico total de cada sitio se componga al menos de cien especímenes (Acosta et al. 2010; Barboza y Martín 2014; Bonomo et al. 2010; Del Papa y Lamenza 2019; Del Papa et al. 2019; Feuillet Terzhaghi y Escudero 2011; Ottalagano 2016, 2019; Ottalagano et al. 2015; Pérez Jimeno 2007; Pérez Jimeno y Servin 2017; Píccoli et al. 2017; Rodríguez y Bonomo 2021; Santini 2009; Santini y De Santis 2011; Sartori 2013; Sartori y Colasurdo 2011; Sartori y Pérez Jimeno 2012; Zarza et al. 2019). Se consideró la presencia del ñandú en los registros tanto por evidencias osteológicas, como por cáscara de huevos.
Acorde a los objetivos propuestos, se decidió realizar el análisis sobre la base del NISP y el NISP%, ya que son las medidas que se encuentran disponibles para todos los conjuntos y cuyos cálculos son unívocos. Si bien, se es consciente de las dificultades de derivar directamente de estos valores patrones de aprovechamiento y/o subsistencia, se considera que el NISP permite abarcar una mayor cantidad de datos, lo cual resulta fructífero para los fines comparativos de este trabajo (Grayson 1984; Lyman 1994, 2003, 2008). Es por ello que se consideró a esta medida como óptima para poder realizar comparaciones intersitio a nivel regional, ya que otras medidas zooarqueológicas tradicionales (Número mínimo de elementos -MNE- y Número mínimo de individuos -MNI) conllevan mayores diferencias a la hora de calcularlas (Grayson 1984; Grayson y Frey 2004; Lyman 1994). Debe tenerse en cuenta que, si bien la información faunística es amplia, el nivel de análisis con que se han tratado los diferentes conjuntos es heterogéneo, con lo cual resulta un desafío poder realizar comparaciones de grano fino o análisis integradores.
Los aspectos tafonómicos y metodológicos (e.g. uso de zaranda) deben haber incidido en la representación de los diferentes taxa que componen cada conjunto arqueofaunístico, dada la diversidad en la procedencia de los mismos. No obstante, tales variaciones suelen afectar en mayor medida, ya sea a especies/elementos muy pequeños o aquellas unidades anatómicas que poseen baja densidad mineral ósea. Los análisis tafonómicos que realizaron los diferentes autores en los conjuntos incluidos en este estudio, de acuerdo a la bibliografía disponible, difieren por su menor o mayor profundidad. En estos últimos casos, los resultados reflejan que, si bien hay agentes que afectan el registro arqueofaunístico, en general existe una buena preservación y una alta recuperación de especies medianas-pequeñas (e.g. roedores y peces) (Pérez Jimeno 2007; Píccoli et al. 2017; Santiago 2012; Sartori 2013; Sartori y Colasurdo 2011; Sartori y Pérez Jimeno 2012). Por esto, teniendo en cuenta que los conjuntos arqueológicos estudiados presentarían -a grandes rasgos- condiciones similares de acumulación y/o preservación,y si bien puede haber sesgos debido a la DMO, se presume aquí que no fueron de significancia, por lo menos para implicar su ausencia en el registro.
Además de tener en cuenta la representación anatómica, se relevaron las evidencias de modificaciones antrópicas en los diferentes sitios, en aquellos casos que se dispone de información. Respecto de estas últimas se consideraron su morfología y localización en la superficie del hueso, de acuerdo a los criterios utilizados por diferentes autores para la zona de estudio (Del Papa et al. 2019; Pérez Jimeno 2007; Pérez Jimeno y Servin 2017).
Finalmente se analizó la distribución geográfica de este registro y se observó si éste se podría relacionar, ya sea, con características ecológicas o con un momento particular del Holoceno tardío, teniendo en cuenta los fechados radiocarbónicos disponibles para cada uno de los sectores consignados. Por último, para la elaboración del mapa se procedió a georreferenciar cada sitio según la información publicada, utilizándose el programa QGIS 3.8 Zanzibar.
Subsistencia y explotación faunística
En la zona seleccionada los análisis arqueofaunísticos fueron realizados por diversos equipos de investigación, notándose un notable incremento en este tipo de estudios en los últimos 15 años. Considerando que el objeto de este apartado es sintetizar la información generada para los tres sectores diferenciados en la llanura aluvial del Paraná, aquí se exponen de manera resumida los trabajos que realizaron análisis específicos de los conjuntos y que a partir de ellos generan expectativas en torno a las estrategias de subsistencia de los grupos cazadores-recolectores-pescadores de la región (ver Anexo).
Para el sector Chaqueño Ribereño del sistema Paraná-Paraguay (CHH), los registros arqueológicos más antiguos corresponden a los sitios Sotelo 1 yPotrero IVb, con fechados radiocarbónicos de 1690±90 y 1680±100 años AP, respectivamente (Santini y De Santis 2011); mientras que el segundo sitio también posee fechados con una antigüedad estimada de ca. 900 años AP (Zarza et al. 2019). Los demás sitios de esta región quedan comprendidos dentro de este rango cronológico, excepto por el más tardío, La ilusión 1 (630 ± 40 años AP) (Del Papa y Lamenza 2019; Santini y De Santis 2011). A partir de los análisis zooarqueológicos de los sitios de este sector, se planteó que la subsistencia se habría basado en la explotación intensiva de recursos tales como peces, coipo y moluscos, todos provenientes de ambientes lénticos cercanos a los campamentos. En ellos se distinguieron dos modalidades de explotación, una basada en el aprovechamiento predominante de Myocastor coypus, complementado con Blastocerus dichotomus, Mazama sp., Cavia aperea, peces y moluscos, y otra donde predomina la explotación de cérvidos (si bien los valores de abundancia son similares, varía la importancia relativa en el conjunto) además de una mayor proporción de peces. La estacionalidad, de pulsos de agua-seca, fue propuesta como una posible causa de estas diferencias (Santini 2009; Santini y De Santis 2011; Santini et al. 2003).
En el sector LLA-a, el depósito más antiguo corresponde al sitio La Lechuza, que posee un fechado radiocarbónico de 1760±60 años AP (Cornero et al. 2007), siendo más recientes las ocupaciones de los sitios Puesto Fantin (930±70 años AP) y Cerro Aguará (530±70; 895±60 años AP) (Pérez Jimeno 2007; Santini 2009). El modelo de ocupación humana para esa zona (Pérez Jimeno 2007), sostiene que la misma habría estado ocupada por grupos semisendentarios, ligados estrechamente a los ambientes lénticos generados por la dinámica del río Paraná. Esos grupos, a su vez, habrían contado con una tecnología especializada (cerámica, ósea y/o lítica), que les permitía basar su subsistencia en la pesca (ver Pérez Jimeno 2005, 2007; Musali y Pérez Jimeno 2016) y la caza de mamíferos, fundamentalmente cérvidos -Blastocerus dichotomus y Ozotoceros bezoarticus-, pero también roedores -Myocastor coypus, Cavia aperea e Hydrochoerus hydrochaeris-) (Mucciolo y Pérez Jimeno 2015; Pérez Jimeno 2007; Santiago 2012), predominando unos u otros de acuerdo a la alternancia de períodos de bajantes o crecidas. Además, complementarían la dieta con otros recursos como aves, reptiles y la recolección de moluscos. De esta manera, los ambientes lénticos serían explotados intensivamente durante los períodos de bajante del río, ya que a comienzos de los períodos de inundación esos grupos se fraccionaban y desplazaban hacia otros sectores topográficamente más elevados (ver Pérez Jimeno 2007). En coincidencia con esto hay algunas diferencias respecto a la composición de los conjuntos, que ha sido explicada en términos espaciales, como las del sitio Campo Binaghi, en el que se registró la explotación de recursos propios de áreas topográficamente más altas y abiertas (ver Sartori y Pérez Jimeno 2012).
En este mismo sector, LLA-a, entre los sitios localizados en Corrientes, sobre la margen izquierda del río Paraná, el de mayor antigüedad hasta el momento es El Nene de 1753±40 AP años AP, mientras que Paso del Tala y Los Bananos cuentan con dataciones más tardías, 356±41 años AP y 355±43, respectivamente (Píccoli et al. 2017). Dichos sitios "habrían sido ocupados de manera redundante y con permanencias prolongadas,
tomando en cuenta la falta de incongruencia espacial y/o temporal de las especies explotadas y la disponibilidad de los recursos necesarios para la manufactura cerámica" (Píccoli 2014: 301). Los estudios zooarqueológicos realizados, señalaron una subsistencia basada en peces y mamíferos propios del ambiente donde están emplazados los sitios, destacándose roedores como Myocastor coypus e Hydrochoerus hydrochaeris (Barboza 2014, 2016; Barboza y Martín 2014; Martín 2015). Para esta zona Píccoli (2014) y Píccoli et al. (2017) sostuvieron la existencia de un bajo riesgo en la obtención de recursos, la cual no estaría marcada por una especificidad estacional.
En el sector LLA-b, en el segmento de la llanura aluvial del Paraná que atraviesa el espinal y pampa, también se localizaron sitios sobre ambas márgenes. En la provincia de Santa Fe el fechado más antiguo se registró en el sitio Río Salado Coronda II (3077 ± 67 años AP, Feuillet Terzaghi y Loponte 2021) -el cual constituye el más antiguo de toda la región-; mientras que en Entre Ríos, en el sitio Laguna de Los Gansos 1 con 1775±51 años AP (Bonomo et al. 2016). En este sector los fechados más tardíos corresponden a Familia Primón (370±30 años AP, Sartori 2013), en la provincia de Santa Fe, y Co. Tapera Vázquez (520±60 años AP, Rodríguez y Bonomo 2021), en la provincia de Entre Ríos. Uno de los modelos propuestos para la zona considera que, ca. 1000 años AP, se produce una recurrencia en la ocupación de espacios insulares y sectores de borde de la planicie de inundación del Paraná (Sartori 2013, Sartori et al. 2017). Estos últimos se ubican en terrenos altos, con una buena visibilidad, situados próximos a cursos de agua, pero no inundables. Esto permitiría el abastecimiento de recursos críticos como agua, leña y gran cantidad de vegetales y animales consumibles. Por su parte, los espacios insulares son intensamente ocupados, debido al acceso a los recursos faunísticos que propician, fundamentalmente la obtención de peces en ambientes de bajo riesgo y mayor concentración. En cuanto a los recursos faunísticos para la zona, éstos reflejan la diversidad de taxa aprovechados, entre los cuales se encuentran aquellos que son de alta jerarquía (i.e. B. dichotomus, O. bezoarticus y H. hydrochaeris) y de baja jerarquía que pueden ser obtenidos en masa (i.e. M. coypus y peces). La variabilidad de especies presentes indica un predominio de peces en los sitios insulares, mientras que los sitios del borde occidental de la llanura aluvial reflejan un mayor aprovechamiento de mamíferos. De todos modos, en ambos casos se opta por estrategias que reducen o minimizan el riesgo al incluir presas de baja jerarquía (coipo, peces) en la dieta (Sartori 2008, 2010, 2013; Sartori et al. 2020).
Ya atravesando la región pampeana en el Paraná medio e inferior, del lado entrerriano Bonomo y colaboradores (2010, 2011, 2016) plantean que se habrían ocupado principalmente los ambientes fluviales a los cuales se accedería mediante el uso de canoas y donde los cursos de agua integrarían sistemas de circulación entre sitios. Estos autores proponen que varios sitios estarían localizados en montículos de tierra antrópicos (“cerritos”), los cuales habrían sido intencionalmente elevados. Respecto al registro faunístico, ha sido planteada una dieta basada en recursos acuáticos (Bonomo et al. 2011, 2016), reconociéndose varios taxa, entre los cuales las especies más abundantes habrían sido Myocastor coypus, Hydrochoerus hydrochaeris, Blastocerus dichotomus, Cavia aperea y carnívoros (Rodríguez y Bonomo 2021).
Resultados
De los 25 registros zooarqueológicos considerados, cinco corresponden a sitios que se localizan en el sector CHH, con dataciones entre los ca. 600 y 1700 años AP; ocho en el sector LLA-a con fechados de entre ca. 350 y 1750 años AP; y 12se sitúan en el sector LLA-b, comprendidos cronológicamente entre los ca. 500 y 3000 años AP.
De los sitios analizados, solo se verificó la presencia de Rhea americana en nueve de ellos -36% de los conjuntos-. El NISP total del área estudiada, es de 69 especímenes (Tabla 2), de los cuales 55 corresponden al total registrado en el sector CHH; 12 al sector LLA-a y dos al sector LLA-b. En los cuatro conjuntos con presencia de Rhea americana en el primer sector (CHH), los valores de NISP varían entre 2 y 26, mientras que en el segundo (LLA-a) entre tres y seis especímenes, y en el tercero (LLA-b), son sólo de un espécimen en cada sitio (Figura 2). En cuanto a los NISP% para esta especie en todos los conjuntos no supera el 1,32%, valor que solo alcanza en un sitio, mientras que en los demás no supera el 0,5% (tabla 2); y, de acuerdo a la información bibliográfica consultada, el NMI es solo de un individuo en cada uno de los sitios.

Tabla 2: Representación de R. americana -NISP y NISP%. * En este caso se identificó a nivel Familia (Rheidae).
Atendiendo a la distribución espacial, se observa un predominio de registros de ñandú en el sector norte de la zona analizada, y éstos decrecen hacia el sur (Figura 2). Cabe señalar que hay una concordancia, entre el sector donde se dan las mejores representaciones en términos de NISP y NISP% -que es el sector del CHH y donde hasta el momento se han registrado más conjuntos arqueológicoscon presencia de R. americana. Esto además coincide con una mayor frecuencia de la clase Aves en los sitios de este sector(Figura 3). En cambio, en los sitios de la LLA-a, donde la representación de este macrotaxón es similar, la presencia del ñandú es más limitada. Finalmente, en los sitios de LLA-b, ambas categorías taxonómicas son menos

Figura 2: Distribución de R. americana en el área de estudio: señala los sitios donde se registra sin modificaciones antrópicas; donde se consignaron especímenes de esta especie con modificaciones antrópicas.

Ahora bien, considerando el marco temporal, los registros analizados abarcan desde ca. 3000 años AP hasta los ca.350 años AP. No obstante, no se observa una relación entre la presencia/ausencia del ñandú vinculada a la temporalidad, ya que como puede verse en la Tabla 2 y Figura 4, en el sector CHH, está
presente en similares proporciones en conjuntos correspondientes a momentos más tempranos (i.e PIVa 1180±70 años AP; 1260±80 años AP) como en otros más tardíos (i.e Li1 630±40 años AP). En la LLA-a sucede algo parecido, en un sitio más temprano (i.e LL 1760±60 años AP) la representación es similar a la de otros más tardíos (i.e. PF 930±70 años AP y CA 530±70 años AP; 895±60 años AP). Así también, en la LLA-b, hay muy baja representación y/o ausencia tanto en sitios más tempranos como en otros más recientes (ver Tabla 2 y Figura 4).

Figura 4: Fechados arqueológicos de los sitios: a) en azul son sitios en los cuales no se registran restos de R. americana, b) en verde indican conjuntos con presencia de R. americana.
En general, la presencia de R. americana fue señalada a partir de especímenes óseos, aunque en dos sitios del sector CHH se registraron además cáscaras de huevo (Li1 N=5 y PIVb N=1). En cuanto a la representación anatómica (Tabla 3), en el conjunto de los sitios se observó un predominio de las unidades anatómicas de los miembros posteriores, principalmente del segmento distal (tibiatarso, tarso-
metatarso y falanges). Solo se registraron elementos del esqueleto axial en un sitiodel sector CHH (en PIVa), y del cráneo en uno de la LLA-a (en CA).
En el sector CHH los elementos mejor representados corresponden a: vértebras (N=10), tibiotarso (N=7), húmero (N=5) y falanges (N=5). En el sector LLA-a, el cráneo se registró escasamente -solo está presente en un sitio-, mientras que el esqueleto axial está ausente y el esqueleto apendicular se encuentra representado en menor proporción que en el sector antes referido, aunque por los mismos elementos (tibiotarso, tarso-metatarso y falanges). En el sector LLA-b, de acuerdo a la información que se cuenta hasta el momento, solo está representada en un solo sitio por un elemento del miembro posterior (Tabla 3).
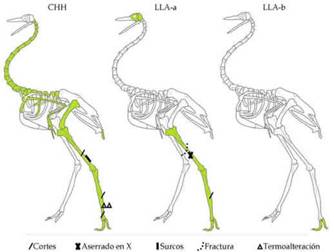
Figura 5: Representación anatómica de R. americana en los tres sectores considerados (CHH, LLA-a y LLA-b). Se indican en cada caso la presencia de alteraciones antrópicas relevadas.

En cuanto a las evidencias de procesamiento antrópico se observaron en cuatro sitios: dos de ellos se emplazan en el sector CHH (sitios Li1 y PV), dos en la LLA-a (sitios PF y CA), mientras que en el sector más al sur de la zona analizada -y que es la que presenta los NISP
más bajos-, no se registra ninguno. En Li1 se registraron un tarso-metatarso y un tibiotarso con marcas de corte, y dos tarso-metatarsos termoalterados (Del Papa y Lamenza 2019). En PV, se identificaron un tarso-metatarso con marcas de procesamiento y una diáfisis de tibiatarso con dos surcos longitudinales profundos y paralelos, los cuales se observan en toda la extensión de su cara posterior presentando incisiones superficiales subparalelas a éstos, las que fueron interpretadas como "tentativas de formatización" (Del Papa et al. 2019:4). Además, en este elemento los surcos habrían sido producidos para obtener láminas óseas como formas base, mediante la técnica de ranurado (sensu Averbouh y Provenzano 1999). Los autores consideran que esas láminas óseas podrían haber sido utilizadas para la formatización de puntas planas, acanaladas y bipuntas, siendo los grupos morfológicos que presentan mayor modificación de la superficie (Del Papa et al. 2019).
Por último, dentro de la LLA-a, en el sitio PF, se registró una diáfisis con huellas de corte (Santini 2009) y en el sitio CA un tibiatarso con aserrado en X y fractura transversal curvada (Pérez Jimeno y Servin 2017). Este último patrón, que también se registró en huesos de mamíferos del mismo sitio y en otros del Paraná medio, se ha asociado a la producción de tecnofacturas. Como en el caso anterior, este marcado en X seguido de fractura por impacto permite obtener dos extremidades en punta con bajo costo, al implicar menos tiempo de trabajo y materiales (Buc et al. 2013; Pérez Jimeno 2007; Pérez Jimeno y Servin 2017). También en CA se registraron diversos grupos de puntas, que como señalaron Del Papa et al. (2019), el grado de modificación de los soportes para su formatización podría
dificultar la identificación de la especie utilizada como soporte.
Discusión
A partir de la información relevada pudo observarse cómo, en general, el registro de Rhea americana es escasoen toda el área considerada. No obstante, una mayor presencia se observa en los sectores CHH y LLA-a, donde el macrotaxón Aves, si bien constituye porcentajes bajos del NISP total de los sitios, está mejor representado, incluyendo varias especies medianas/grandes de hábitos acuáticos (Anatidae, Anhinghidae, Phalacrocoridae, Podicipedidae, entre otros). Si bien se trata de aves de ámbitos acuáticos, diferentes al propio de R. americana, lo que se quiere señalar es que esta Clase fue incluida en la subsistencia.
Si se atiende a la distribución espacial, puede advertirse que -en términos generales- existe una concordancia entre su mayor representación con las características ambientales en donde se emplazan los sitios. Esta misma situación ha sido observada en otros estudios regionales en la zona en donde se concluye que la variabilidad y abundancia arqueofaunística van de la mano de la oferta de los recursos disponibles en sectores próximos (Sartori et al. 2014, 2017).
De los tres sectores, el del CHH es donde el ñandú se registró con una mayor representación, aunque en un solo sitio, con cierta completitud anatómica. Algunos de los especímenes presentaban marcas antrópicas, y, además, en uno de los sitios, se hallaron cáscaras de huevo. En LLA-a solo está presente en sitios de la margen derecha del Paraná. Es posible que su ausencia en la margen izquierda se vincule a diferencias en la integridad de los conjuntos -respecto de los observados en sitios arqueológicos de la otra margen-, ya que los autores han señalado que el tamaño y la frag-
mentación de los restos óseos habría incidido en la determinación anatómica y taxonómica (Píccoli 2014; Píccoli et al. 2017). No obstante, hasta que no se cuente con mayor evidencia no puede descartarse que la ausencia de este taxón no se deba a elecciones culturales (ver infra). En LLA-b la representación de Rhea americana es menor aún que los conjuntos de la LLA-a, registrándose escasamente y solo en un sitio de cada una de las márgenes. En estos casos no se considera, al menos de manera exclusiva, que las condiciones de preservación diferencial puedan estar afectando significativamente a esta especie, ya que por ejemplo en el sitio FP se han registrado elementos que son de baja preservación, tales como, escamas de peces (Sartori et al. 2020).
Estudios arqueofaunísticos regionales reflejan una mayor cantidad de taxa (NTAXA) hacia el sector norte, la cual decrece en sentido sur, donde se da una marcada disminución en la representación de Aves y Reptiles en los conjuntos (ver Acosta et al. 2010; Sartori et al. 2014, 2017). Por otro lado, solo cinco sitios distribuidos en los tres sectores considerados (Li1, LL, ALM1, CTV y PM) presentan evidencias de otros taxa propios de ambientes más secos y abiertos, como camélidos y/o venado. Cabe señalar que la mayoría de estos sitios no se encuentran en la zona insular, sino en el sector continental que presenta ambientes más aptos para tales especies. No obstante, es notoria la casi nula presencia de R. americana en los sitios de la LLA-b siendo que se encuentran más próximos al Espinal y la Pampa, los cuales permiten el acceso a zonas abiertas más aptas para el hábitat de esta especie corredora. Una consideración al respecto es que la mayor parte de los sitios del interior de la llanura aluvial del río Paraná no presentan recursos propios de la zona continental (ver Sartori et al. 2014,2017), y reflejan exclusivamente la explotación de los ambientes lóticos-lénticos.
Entonces, por lo expuesto aquí y a partir de los datos bibliográficos considerados, R. americana no parece haber constituido un recurso altamente explotado en la región. Esto llama la atención si se tiene en cuenta que los recursos que pueden obtenerse de este taxón son variados e incluye tanto la carne, la grasa, los tendones, la médula, como a los huesos, uñas y plumas. Si bien los valores más altos de contenido de carne se hallan en las unidades anatómicas del miembro posterior (fémur y tibiatarso), la cintura pélvica y el esternón presentan la mayor proporción de grasa (Giardina 2016). Estas últimas secciones del esqueleto se encuentran ausentes en los registros arqueológicos analizados. Por su parte, los valores más altos de médula se encuentran en el fémur, tibiatarso y tarsometatarso, de los cuales además es factible aprovechar los tendones y los huesos como materia prima. Es decir que, hay diversas secciones de esta especie que serían de alto valor por su rendimiento, no sólo en carne sino también en grasa (Santini 2009). Aunque en la región de estudio, la grasa puede ser considerada un recurso de alto valor, y algunas fuentes documentales consultadas (Arenas y Porini 2009; Paucke 1942 [1749-1767]) refieren su utilización entre algunos grupos originarios, la elección de los taxa para su aprovechamiento puede vincularse a múltiples factores diferentes al económico (i.e. palatabilidad, preferencias, creencias). No obstante, es posible que los grupos de la zona contaran con otras presas ricas en grasas como los peces siluriformes y el carpincho - que son mencionados en las fuentes etnohistóricas-etnográficas-, los cuales se encuentran altamente disponibles en las proximidades de los sitios y pueden obtenerse con un bajo costo de búsqueda-captura (Pérez Jimeno 2007; Sartori 2013; Servin 2021). Entonces, la grasa
proporcionada por el ñandú, podría haber sido no imprescindible para los grupos que habitaron los diferentes sectores de la LLAP, más, considerando las dificultades que acarreaba su captura.
En cuanto a la representación de partes esqueletarias en toda la LLAP predominan los elementos del miembro posterior, tarso-metatarso y tibia-tarso; patrón que es coincidente para otras regiones de Argentina (Acosta 2005; Cruz y Elkin 2003; Del Papa 2012; Fernández 2010; Fernández et al.2001; Salemme y Frontini 2011; entre otros). No obstante, se pueden señalar algunas excepciones: en el sector CHH, se registró el esqueleto casi completo en un sitio (PIVa) y parte del esqueleto axial en otro (LI1); mientras que en LLA-a, el cráneo está representado solo en el conjunto de CA. El predominio de los miembros posteriores podría corresponder al trozamiento y traslado selectivo de las diferentes secciones del esqueleto, como menciona una de las fuentes documentales (Paucke 1942 [1749-1767]). Esto podría estar vinculado a que el miembro posterior y el esqueleto axial, presentan en términos generales un mayor rendimiento económico (Giardina 2006, 2010), mientras que la disponibilidad de carne y grasa del miembro anterior es muy baja. Por su parte Giardina et al. (2021) observaron que la fractura del traso-metatarso y el consumo de la médula ósea se producen en el lugar de captura; mientras que el resto del miembro posterior es trasladado para su posterior procesamiento y consumo. Por otro lado, las diferencias en la representación en los conjuntos de los sitios también podrían deberse a las diferencias estructurales y anatómicas entre el miembro anterior y el posterior, reflejadas en su correspondiente densidad mineral ósea (Cruz y Elkin 2003; Giardina 2010). Hasta el momento no se cuenta con estudios tafonómicos específicos en la región considerada que permitan evaluar la incidencia de estos factores en la preservación diferencial de las distintas unidades anatómicas de esta ave. No obstante, y asumiendo que las condiciones de preservación diferencial no habrían incidido significativamente en los sitios, se considera más factible que la baja presencia-ausencia de R. americana esté condicionada por decisiones culturales -i.e. palatabilidad, restricciones alimenticias-, su mayor o menor disponibilidad y las dificultades asociadas a su caza; o una combinación de tales aspectos.
En muchos trabajos se le ha dado una gran importancia al ñandú respecto a otros recursos alimenticios que se pueden obtener en la región pampeana; este ranking siempre tuvo como referencia el tamaño del animal (Álvarez 2015; Frontini y Picasso 2010; Martínez y Gutiérrez 2004; Medina et al. 2011). No obstante, si se tiene en cuenta el gasto energético que costaría a las poblaciones pedestres de cazadores-recolectores poder capturar un espécimen de Rhea americana, considerando sus comportamientos etológicos, decaería en importancia y pasaría a ser un recurso complementario (Giardina et al. 2021). En estos casos, sería el producto de una cacería no programada y esporádica, en relación a otros recursos alimenticios disponibles, y como fuera planteado por Salemme y Frontini (2011), la cacería se realizaría en lugares puntuales. Por el contrario, los huevos son más fáciles de recolectar y muchas veces es predecible el lugar donde se encontrarían las nidadas (Giardina et al. 2014), sin embargo, por el momento son escasas las evidencias de su consumo.
La dificultad en la captura podría ser una de las razones por la cual, pese a tener un gran contenido de carne y grasa, y ofrecer un aprovechamiento íntegro, nunca fueron realmente explotados hasta la aparición del
caballo2, que mejoró las posibilidades de caza, y es lo que registrarían las crónicas de naturalistas y viajeros, más arriba referidas. Para los primeros momentos de contacto la información de los escritos de Schmidel (2003 [1564]), sólo refiere la presencia del ñandú, entre otros tantos recursos que conformaban la dieta de las comunidades encontradas en su recorrido (ie. Chanás, Wichís y Mepenes), destacando que se daría un consumo mayoritario de peces. Los estudios etnográficos de Miraglia (1975), Paucke (1942 [1749-1767]), Arenas y Porini (2009), Medrano (2014), dan cuenta que Mocovíes, Wichís y Qom, hacían un uso integral de esta ave, tanto como alimento, como para uso medicinal y como materia prima, utilizando algunos de sus huesos, y también su cuero. Además, es factible pensar, a partir de lo señalado por estos últimos, que: por un lado, la disponibilidad del caballo para movilizarse de algunos grupos de cazadores-recolectores, puede haberles proporcionado mayores facilidades para su captura; y por otro, si una vez capturadas, se trozaban en los lugares de caza, y se trasladaban a los asentamientos solo los miembros o parte de ellos, es probable que en los sitios arqueológicos no se hallen representadas todas las secciones del esqueleto, y se observen en los registros zooarqueológicos porcentajes bajos de esta especie respecto de otros recursos, como los mamíferos. Además, las formas de cocción señaladas también podrían estar generando dificultades para su posterior identificación en dichos registros (Arenas y Porini 2009).
En consecuencia, se presenta una disonancia entre la información arqueológica y la relevada en las fuentes documentales consultadas, donde Rhea americana es retratada como un ave de relevancia, en los diferentes ámbitos de las comunidades como la Mocoví, Wichi y Qom; es decir, no solo apreciable por el valor alimenticio de su carne, grasa y huevos, sino por las plumas utilizadas, tanto para fabricar utensilios cotidianos como elementos de connotación cinegética, bélica, simbólica y chamánica; así como el uso medicinal también de su grasa, huesos y buche (Arenas y Porini 2009; Medrano y Rosso 2016). Además, señalan su valor simbólico, ya que sus cualidades podían ser transmitidas tanto a partir de su consumo como alimento o medicina, o como por la utilización de objetos fabricados con sus huesos o plumas. A su vez, probablemente por ser un ave “poderosa”, presenta ciertas restricciones en su consumo como alimento. Los aspectos antes mencionados la posicionan como una especie "etnobiológica clave" (Medrano y Rosso 2016:61), ya que posee un rol fundamental en las tramas ecológicas y culturales de la región durante el siglo XVIII (Rosso y Pautaso 2017).
Por otra parte, cronológicamente no se observa un patrón claro que refleje mayor-menor intensidad en el uso de este recurso en los momentos previos a la llegada de los europeos. Esto podría tener dos explicaciones, por un lado, que no se cuenta aún con datos que sean lo suficientemente robustos como para poder evaluar tal hipótesis; o bien, como se esboza en este trabajo su presencia-ausencia se relaciona más con el lugar de emplazamiento de los sitios y su proximidad a zonas de captura. Además, si se tiene en cuenta que la pesca ha sido señalada como la base de la subsistencia de los diferentes grupos que habitaron la región, es posible que la caza de otros recursos actúe de manera complementaria.
Por lo expuesto, puede concluirse que, con la evidencia arqueológica disponible hasta el momento para la región, R. americana habría sido un recurso poco explotado, fundamentalmente para el sector meridional de la llanura aluvial del río Paraná, al menos previa a la introducción del caballo. En los sectores de más al norte, en cambio, su representación refleja cierta relevancia como recurso complementario de la dieta, ya que las huellas antrópicas registradas en uno de los sitios del sector CHH (en un tarso-metatarso) se asoció con las propias del aprovechamiento de médula y tendones, las cuales son coincidentes con las observadas en sitios de Pampa (e.g. Acosta 2005; Frontini y Picasso 2010; Quintana y Mazzanti 1996, 2001), Patagonia (e.g. Salemme y Frontini 2011), y en sitios de la región Chaco-Santiagueña (Del Papa 2012). Mientras que otro de este mismo sector (sitio PV) y las registradas en el sitio CA, del sector LLA-a, se relacionarían tanto con el aprovechamiento de la médula como con la producción de artefactos, -si bien por el momento no se identificaron artefactos formatizados en huesos de esta especie.
Agradecimientos
Queremos agradecer a la Dra. Mónica Salemme por la lectura crítica del manuscrito y sus valiosos comentarios realizados. Así también a los evaluadores por sus observaciones y sugerencias que contribuyeron a mejorar las ideas presentadas en este trabajo. No obstante, lo expresado aquí es responsabilidad exclusiva de las autoras.
Notas
1Si bien a lo largo del texto se hará referencia a la misma como R. americana, se trata de la subespecie R. a. albescens, que es la que corresponde al área de estudio de este trabajo.
2No obstante, las observaciones realizadas por Giardina et al. (2021) también señalan que la captura del ñandú, incluso a caballo, es una tarea difícil y peligrosa para los cazadores, ya que a menudo éstos tropiezan con cuevas de roedores y se caen durante la persecución

