











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
En este trabajo se presenta nueva evidencia arqueofaunística de la capa F4 del sitio Alero Cuevas, Puna de Salta, Argentina (Figura 1), la cual cuenta con fechados radiocarbónicos correspondientes al Holoceno temprano, ca. 9880-8500 años AP (López 2013; López y Restifo 2017).El objetivo es analizar el uso alimenticio humano de recursos faunísticos en el sitio, con el foco en la explotación de camélidos, principaltaxón recuperado y de gran importancia económica en la región. Al mismo tiempo, se busca comparar y discutir este registro en relación con la evidencia de otros sitios delHoloceno temprano en los Andes Centro Sur, especialmente del Noroeste argentino y el norte de Chile. Más precisamente, se aporta nueva información para entender los procesos tempranos de explotación alimenticia de camélidos y lastendencias en suprocesamiento y consumo a escala macrorregional.
En términos generales, la Puna argentina se extiende geográficamente desde el altiplano andino (sur de Perú, oeste de Bolivia, norte de Chile) yes, orográficamente, una meseta de altura que conforma un ambiente ecológico particular (Muscio 1999; Turner 1964;Vilela 1969). Este ambiente presenta oscilaciones altitudinales entre ca.3200-4600 ms.n.m. (con picos de más de 5000 ms.n.m.), gran cantidad de cuencas endorreicas, lagunas y salares que constituyen los espacios de circulación y concentración de agua, que permiten el crecimiento permanente de recursos vegetales y agrupamiento de animales (Muscio y López 2011; Turner 1964;Vilela 1969;Yacobaccioet al. 1994). Se caracteriza por ser un desierto de altura con la presencia de condicionamientos para la adaptación humana como la extrema aridez, la hipoxia y la alta radiación solar (Muscio y López 2011). Asimismo, en estos ambientes de altura, la diversidad de recursos animales es limitada siendo los camélidos losrecursos de mayor rendimiento económico en contraste con otros de menor ranking como los chinchíllidos (Muscio y López 2011).
La Puna de Salta, comparte características ambientales regionales del altiplano andino pero se destaca por ser un sector intermedio entre la Puna de Jujuy y la Puna de Catamarca (López 2008), a una altura promedio de 3500 ms.n.m., con baja productividad primaria, baja diversidad de biomasa, distribución segmentada de agua, caída impredecible de precipitaciones (entre 30 mm y 100 mm anuales) y una gran amplitud térmica (Morales 2011; Yacobaccio 1994). En este sector, se ubica la cuenca de Pastos Grandes con sus particularidades geoambientales definidas y divididas, de acuerdo a sus aspectos geomorfológicos, ecológicosy distribución de recursos, en: vega principal, borde de salar y las quebradas de altura (López 2008; Muscio 2004).Estas quebradas, cuentan con cursos de agua permanente que permiten una fertilidad constante para el crecimiento de pastizales. Este es el caso de la quebrada denominada localmente como Las Cuevas, donde se ubica el sitio Alero Cuevas. Estos sectores constituyen, además, el principal espacio atractivo para la concentración, permanencia y disponibilidad de animalessilvestres entre los que se destacan los camélidos.
El sitio Alero Cuevas fue detectado en el transcurso de una campaña arqueológica durante el año 2004 (López 2007). Este sitio conservó una secuencia estratigráfica con material arqueológico que pudo ser fechada a lo largo del Holoceno temprano, medio y tardío (López 2008, 2009, 2013; López y Restifo 2017). Debido a la conservación del material y su extensa cronología, hasta el presente existen diferentes investigaciones que continúan dando información novedosa para la discusión de las estrategias de consumo de camélidos y las tendencias de continuidad y cambio en la explotación de estos recursosen la Puna de Salta (López y Orsi 2017, 2019;López y Restifo 2014; Orsi 2018;Orsi y López 2019).
El área de estudio constituye un lugar propicio para la concentración de fauna silvestre como camélidos, cérvidos, aves y roedores. Por un lado, entre los camélidos silvestres se encuentra la vicuña (Vicugnavicugna), que tiene una distribución acotada a ambientes andinos, y presenta un peso medio entre 35 y 50 kg y una morfología grácil (Vilá 2012). Por otro lado, se destaca el guanaco (Lama guanicoe) que presenta una amplia distribución geográfica latitudinal desde el norte de Perú hasta Tierra del Fuego y la Isla Navarinoen el sur de Tierra del Fuego (entre ca. 8°30´S y 50°S). Esta amplia distribución implica una gran variabilidad de ambientes continentales e insulares en los cuales el guanaco habita y se encuentra adaptado. Asimismo,entre los guanacos existendiferencias sustanciales de acuerdo al tamaño corporal y los pesos que oscilan entre los 90 kg, en sectores septentrionales, y 150 kg en sectores australes (MengoniGoñalons y Yacobaccio 2006). Se ha planteado que esta diversidad adaptativa responde a los contrastes ambientales característicos del Pleistoceno y el Holoceno; por esta razón los guanacos que han existido durante la transición Pleistoceno-Holoceno temprano tendrían un mayor porte corporal que los de períodos posteriores (MengoniGoñalonsy Yacobaccio 2006).
También se registraotra fauna, por ejemplo, cérvidos como el Hipocamelusantisiensis, comúnmente llamado taruca.Esta especie posee una distribución restringida y una baja densidad (Olivera 1997). Entre las aves, existe un grupo variado en el que se distinguen los flamencos (Phoenicopterusandinus), las guayatas (Chloephagamelanoptera) y en menor medida patos (Anasspp.) y cóndores (Vulturgriphus). Asimismo, debe tenerse en cuenta al suri (Pterocnemiapennata) por su aporte proteicoen carne, médula, grasa y huevos como alimento. En este sentido, las aves constituyen un recurso del que pueden aprovecharse sus huesos y plumas para elaboración de herramientas, ornamentos y ajuares funerarios (ver Urquiza y Echeverría 2018). En la fauna pequeña haypresencia de roedores (Chinchilla brevicaudata, Ctomyopinus, Microcaviashiptoni y Lagidiumviscacia). Este último taxón se destaca por su mayor rendimiento cárnico en contraste con los anteriores, por esto es esperable encontrarlo másrepresentado en el registro arqueológico.
Se ha planteado que los camélidos fueron los recursos cruciales para la instalación humana recurrente en la Puna a partir del Holoceno temprano, debido al aporte sustancial de proteína animal de alto rendimiento para amortiguar eficazmente las presiones ambientales (Muscio y López 2011). De esta manera, se interpreta que la selección de camélidos sobre otros taxones fue una decisión optimizante para los cazadores recolectores de contextos tempranos de tierras altas en general y de la Puna de Salta en particular (López 2008; Martínez et al. 2010;Mondini y Elkin 2014; Muscio y López 2011; Olivera 1997; Urquiza y Aschero 2014). Sin embargo, a nivel macrorregional, también se reconoce una tendencia al consumo de una mayor diversidadde fauna silvestre durante el Holoceno temprano (Yacobaccio 2001, 2017;Yacobaccioet al. 2016). Específicamente, esta tendenciase identifica,en diferentes proporciones, en componentes zooarqueológicos de la Puna de Jujuy, Puna de Catamarca, y norte de Chile (Martínez et al. 2010; Núñez et al. 2010;Yacobaccio 2001; entre otros). Asimismo, los estudios muestran que la caza de animales grandes (guanacos, vicuñas, taruca) y pequeños (aves y roedores) responde principalmente a una explotación centrada en el aprovechamiento alimenticio. Más allá de esta variabilidad, los camélidos constituyen los recursos de mayor representación en el registro arqueofaunístico de la Puna.
Figura 1: Ubicación geográfica del sitio Alero Cuevas en la quebrada de Las Cuevas ca. 4400 ms.n.m., Pastos Grandes (Puna de Salta)
Antecedentes
Paleoambiente y contexto cronológico
El Holoceno temprano se ha caracterizado por condiciones de estabilidad climática, que se inician cronológicamente con el fin de los ambientes glaciares pleistocénicosa partir de ca.12600-10000 años AP, y se prolongan hasta ca. 8000-7800 años AP (Aldenderfer 2011; López y Restifo 2014; Morales 2011;Yacobaccio 2017). Los estudios paleoambientales muestran que estas condiciones habrían sido más húmedas, con una estacionalidad más débil de las precipitaciones, moderada variabilidad climática y mayor productividad primaria, componiendo un ambiente fértil menos fragmentado o heterogéneo que en la actualidad (Morales 2011; Thompson et al. 1995;Yacobaccio y Morales 2005). Esto se debió, principalmente, a que las condiciones de mayor humedad permitieron la permanencia de la estepa herbácea en altitudes más bajas que las actuales durante al menos 2000 años AP (Morales 2011; Oxman 2015; Tchilinguirianet al. 2014).
Asimismo, esta estabilidad ambiental habría favorecido la permanencia y disponibilidad de vegetales forrajeros y, por lo tanto, de una mayor abundancia de biomasa animal, principalmente representada por la alta presencia y distribución de camélidos, recursos fundamentales para la ocupación humana de la Puna (Muscio y López 2011). En este sentido, este escenario ambiental sumado a la disponibilidad de recursos en parches menos segmentados, habrían posibilitado una rápida dispersión humana y, por lo tanto, ocupaciones relativamente estables durante el Holoceno temprano (Muscio y López 2011;Yacobaccio 2017;Yacobaccio y Morales 2013). La finalización de este período está caracterizada por fluctuaciones ambientales globales que afectaron el desarrollo de la abundancia y distribución de recursos críticos de la Puna a largo plazo (Tchilinguirianet al. 2014;Yacobaccio 2013). Estas fluctuaciones se acentúan a partir de ca. 7000 años AP, y caracterizan la transición hacia el período ambiental denominado Holoceno medio en el cuál las condiciones climáticas son de mayor temperatura y aridez (Morales 2011;Tchilinguirianet al. 2007).
De acuerdo a la cronología, los fechados indican que las tierras altas de los Andes se ocuparon alrededor de losca. 11000 años AP (Muscio y López 2011, 2016;Yacobaccio 2017). En términos generales, existe un consenso de que las primeras poblaciones que ocuparon y se dispersaron en nuevos ambientes de la Puna se caracterizaban por baja demografía, alta movilidad y la caza-recolección como la principal estrategia económica (Aschero 1994;Aschero y Martínez 2001; Muscio 1999; Núñez 1992; Núñez y Grosjean 1994;Yacobaccio y Vilá 2002). Esta situación, sumada a las condiciones ambientales que proporcionaban una amplia disponibilidad de recursos, permitieron desarrollar ocupaciones más intensas y redundantes hacia los ca.9500 años AP, especialmente en cuevas y aleros (Muscio y López 2016;Yacobaccio y Morales 2013;Yacobaccioet al. 2016). Si bien los grupos humanos aprovecharon diversas especies de animales grandes y pequeños, las tendencias indican que a nivel general el consumo de camélidos fue la opción principal(Muscio y López 2011;Olivera 1997;Yacobaccio y Vilá 2002).
Evidencia de explotación alimenticia en Puna de Argentina y norte de Chile
En el Holoceno temprano, la evidencia zooarqueológica macrorregional revela un patrón variable en la selección de recursos animales disponibles en el entorno ambiental para la explotación alimenticia. En este sentido, de acuerdo al número máximo de taxa registrado en diferentes sitios arqueológicos, se ha sugerido un comportamiento regional fragmentado en el consumo y explotación alimenticia de las especies silvestres (Yacobaccio 2017). Específicamente, se identifican sitios con predominio de fauna pequeña (e.g. roedores), otros con equilibrio entre camélidos y roedores o predominio de camélidos sobre otros taxones.
El primer caso, el predominio de fauna pequeña, fue identificado en el sitio Hornillos 2, con varios fechados entre 9710 ± 270 años AP y 9150 ± 50 años AP (Yacobaccio et al. 2014). De igual manera, en el sitio Inca Cueva 4, con distintos fechados entre 10620 ± 140 años AP y 9230 ± 70 años AP (Aschero 2014;Yacobaccio 2013) los resultados indican el predominio de roedores (chinchillas y/o vizcachas) por encima de camélidos y cérvidos (Yacobaccio 2004,2017). Para ejemplificar el segundo patrón de aprovechamiento, el sitio Pintoscayoc 1, fechado entre 10720 ± 150 años AP y 9080 ± 50 años AP, evidencia una representación equilibrada entre camélidos y fauna pequeña (Elkin yRosenfeld 2001; Hernández Llosas 2000; Rosenfeld 2002).
Por su parte, la tercera tendencia, el predominio de camélidos sobre otros taxones, se ha registrado en el sitio Huachichocana III, con fechados en la capa E3 de 10280 ± 420 años AP (Fernández Distel 1986;Yacobaccio 2004, 2013, 2017). De la misma manera, en la Puna de Catamarca, en este período de ocupación temprana también se registra un predominio de explotación de camélidos, principalmente de vicuñas y en menor medida guanacos, en el sitio Quebrada Seca 3 (Elkin 1996; Reigadas 2006, 2008). Este sitio presenta una secuencia estratigráfica con contextos arqueológicos fechados entre 9790 ± 50 años AP y 9050 ± 100 años AP (Aschero 2014;Aschero y Martínez 2001;Ascheroet al. 1991; Elkin 1996; Martínez 2014;Mondini y Elkin 2006;Pintar 2008). Asimismo, en consonancia con esta tendencia, se registró el sitio Punta de la Peña 4 (Urquiza y Aschero 2014). Sus resultados indican un alto aprovechamiento de camélidos en su capa más temprana fechada entre 8970 ± 60 años AP y 8320 ± 120 años AP (Urquiza y Aschero 2014). Además, es importante remarcar que en otras áreas cercanas a Quebrada Seca 3, también se registraron otros sitios con contextos arqueológicos tempranos con distintas líneas de evidencia que sugieren la caza a distancia y posible aprovechamiento de camélidos silvestres (Martínez et al. 2010). Este es el caso de Peña de las Trampas (ca. 10200 años AP) entre otros (Martínez 2007; Martínez et al. 2010).
En el sector occidental de tierras altas, específicamente en el norte de Chile se registraron ocupaciones tempranas en diferentes sitios del Salar de Atacama, entre ellos San Lorenzo 1 con fechas de ca. 10400 años AP, Tambillo (ca. 8800 años AP), Tulán 67, Tulán 68 y Tulán 109 (ca. 10590-8200 años AP). Estos sitios presentan evidencia que fue asociada al consumo de camélidos y roedores (Núñez et al. 1999, 2002, 2005, 2010). Especialmente, se destaca el sitio Tuina 5 (ca. 10100-9800 años AP), ubicado a ca. 2800 ms.n.m. Los análisis de los conjuntos arqueofaunísticos de este sitio indicaron el aprovechamiento de diversa fauna. Entre estos huesos recuperados la evidencia muestra el predominio de camélidos por sobre otros taxones, e.g.Cervidae, Chinchillidae (Núñez et al. 2002).
Materiales y métodos
Investigaciones en el sitio
Con respecto a las investigaciones en la Puna de Salta, desde el año 2004 se vienen realizando trabajos sistemáticos en el área de Pastos Grandes que incluyeron la excavación arqueológica del sitio Alero Cuevas (López 2008). Por medio de estas investigaciones, se pudo caracterizar la variabilidad estratigráfica y cronológica de este sitio (López 2013; López y Restifo 2017). Se abrieron un total de nueve cuadrículas y cuatro sondeos en distintos sectores del alero, y en total se excavaron 10,5 m2. La estratigrafía presentó capas fechadas en el Holoceno temprano, medio y tardío, con cambios y continuidades en la evidencia arqueológica (ver López 2008;López y Restifo 2017; Orsi 2018; Restifo 2013). La excavación del sitio se realizó siguiendo la estratigrafía arqueosedimentaria reconocida en los trabajos de campo. Más claramente, los conjuntos arqueológicos se agruparon y analizaron a partir de la distinción estratigráfica hecha en la excavación. Cada capa, si bien presenta una matriz general limosa, se distinguió por las diferencias en la textura, el colory el contenido orgánico y arqueológico, entre otros indicadores. Las capas superiores corresponden a contextos del Holoceno tardío, con fechados entre ca. 2000 y 600 AP (López 2013). Por debajo se ubica una capa con evidencia arqueológica (F2), fechada entre ca. 5100 y 4200 AP, y otra capa pedregosa (F3) datada entre ca. 7500 y 6500 AP.
La capa más profunda, localizada al final de la estratigrafía distinguida, se apoya sobre la roca de caja del sitio, a casi un metro de profundidad, y con un espesor superior a los 20 cm. Se trata de la capa F4, caracterizada por ocupaciones intensas y recurrentes en el Holoceno temprano (López 2008, 2013). Al respecto, se obtuvieron cinco fechados radiocarbónicos de 9880 ( 100 años AP,9650 ( 100 años AP, 9280 ( 140 años AP, 8838 ( 52 años AP, y 8504 ( 52 años AP (López 2008, 2013; López y Restifo 2014, 2017). Desde el punto de vista estratigráfico, esta capa ha sido considerada como un componente, debido a que no pudo dividirse en estratos menores. Se trata de una capa de limo con abundante material orgánico, compuesta de una alta frecuencia de evidencia arqueológica producto de eventos recurrentes de ocupación.Similarmente a lo efectuado en los demás estratos, el agrupamiento de este conjunto fue una decisión adecuada y pertinente en función de la indivisibilidad arqueosedimentaria de la capa. Más allá de la extensión cronológica de F4, esta decisión metodológica fue relevante para distinguir el conjunto del Holoceno temprano de otros correspondientes a capas fechadas en el Holoceno medio y tardío. Por otra parte, si bien algunos procesos pudieron afectar en alguna medida la integridad del conjunto, se considera que sus efectos fueron limitados, tal es el caso de la influencia de roedores (ver más adelante)(López 2008). En síntesis, se espera que el estudio de este conjunto aporte nueva evidencia para discutir la explotación de camélidos y otros recursos faunísticos en Alero Cuevas en el Holoceno temprano, y contribuya también a la discusión de este tema en escala macrorregional.
En los últimos años, se profundizó el análisis de los restos óseos recuperados en este contexto a partir de las nuevas excavaciones realizadas en el sitio (ver Orsi 2018). Los análisis desarrolladoshasta el momento permitieron dar cuenta de un abundante registro arqueofaunístico (López 2013;Orsi 2018). Al mismo tiempo, también se recuperaron instrumentos y desechos de talla de material lítico, los cuales no son objeto de estudio en este trabajo (ver Restifo 2013). Solamente es importante mencionar la presencia de puntas triangulares apedunculadas de amplia replicación en los Andes, que constituyen los instrumentos más característicos de esta capa (López y Restifo 2017).
Análisis de las muestrasarqueofaunísticas
Como se señaló previamente, los conjuntos arqueofaunísticos analizados provienen del contexto estratigráfico más temprano del sitio Alero Cuevas, la capa F4. La determinaciónanatómica y taxonómica de los especímenes óseos se llevó a cabo con el uso de material de referencia del Instituto de Arqueología de la Facultad de Filosofía y Letras, Universidad de Buenos Aires, y el empleo de un Atlas Osteológico (Sierpe 2015).De acuerdo a la muestra estudiada, el grado de fragmentación del conjunto permitió determinar distintos niveles de clasificación taxonómica. En este sentido, los especímenes con rasgos diagnósticos se incluyeron a nivel Familia y las piezas óseas que no presentaron rasgos anatómicos precisos se ubicaron dentro de categorías taxonómicas más abarcativasa nivel de Orden, como por ejemplo Artiodactyla, Rodentia, etc. (Grayson y Frey 2004; MengoniGoñalons 1999;Reitz y Wing 2008). Por otra parte, también se contabilizaron los especímenes No Identificados, que por su alto grado de fragmentación no pudieron ser asignados a una categoría anatómica o taxonómica particular (MengoniGoñalons 2010).
Para la cuantificación del taxón Camelidaese consideraron las medidas de abundancia taxonómica y anatómicade NISP (número de especímenes identificados por taxón), su expresión porcentual NISP%, y el NUSP (número de especímenes no identificados). También se determinó el MNI (número mínimo de individuos), MNE (número mínimo de elementos), MAU (unidades anatómicas mínimas) y su expresión en MAU% para estudiar la representación de unidades anatómicas que ingresaron al sitio. Asimismo, se calculóel PEB (partes esqueletarias básicas) para comparar la frecuencia de unidades de trozamiento(Binford1978, 1984;Lyman1994a, 1994b,2008).
También se caracterizó el perfil etario de los camélidosa partir del estado de fusión de los huesos, aunque complementariamente se consideraron el tamaño y las características generales del tejido óseo (Grant 2014;MengoniGoñalons 1999;Orsi 2018;Reitz y Wing2008).En este sentido, para la determinación de las edades de fusión en camélidos se complementaron los perfiles de alpaca de Kent (1982) y de guanaco de Kaufmann (2009). Las categorías etarias usadas fueron tres:nonato/neonato (hasta 12 meses de edad), inmaduro/subadulto (hasta 36 meses) y maduro/adulto (a partir de 36 meses de edad).Asimismo, en anteriores trabajos se han presentado los resultadososteométricos para la diferenciación interespecífica de especímenes de camélidosa nivel especie de la capa F4 de Alero Cuevas (López y Orsi 2019). Por este motivo, de acuerdo a los objetivos de este trabajo se consideró trabajar a nivel familia.
Para evaluar la fragmentación total de la muestra se utilizó la relación entre el número total de especímenes y el número de especímenes identificados por taxón (NSP:NISP). Más específicamente, para evaluar la fragmentación del subconjunto de camélidos se tomó la razón entre el número de especímenes identificados por taxón y el número de elementos identificados (NISP:MNE) (Wolverton 2002). Los resultados de estos indicadores se expresan en valores del total del conjunto de camélidos, cada uno de los elementos anatómicos y en la media del esqueleto axial y apendicular. Al mismo tiempo, se incluyen en conjunto con el estudio de marcas antrópicas con el objetivo de discutir los patrones de procesamiento. Más adelante se espera ampliar estos análisis con la inclusión de los patrones de fractura y su rol en la fragmentación, incluyendo por ejemplo factores tales como el pisoteo.Por otra parte, se utilizó el rSpearmanen el software estadístico PAST4.3 para correlacionar el MAU% con los índices de utilidad, índice de médula y el índice de secado de carne. De esta manera, se pudo estudiar el rendimiento y aprovechamiento de las partes esqueletarias representadas en relación con el total del conjunto (Borrero 1990; De Nigris y MengoniGoñalons2004). Finalmente, para estudiar la integridad de los conjuntos óseos de camélidos se identificaron marcas antrópicas, marcas no antrópicas, especímenes termoalterados, estadios de meteorizaciónyse correlacionó el MAU% con los valores estándares de la densidad ósea de Sthal (1999)(ver también Behrensmeyer 1978; Benett 1999, Binford 1981;Lyman 1994b; Marean 1995; entre otros).
Resultados obtenidos
Diversidady abundancia taxónómica
El total de la muestra es n=7831 especímenes óseos contabilizados, de los cuales n=5763 corresponden a especímenes no identificados y n=2068 son especímenes identificados en el conjunto arqueofaunístico de la capa F4. La tabla 1 muestra la diversidad de los taxones a nivel de Orden y Familia;se observa que Camelidaees el taxón con mayor representación (NISP= 1084; 52,42%) y en menor porcentaje los artiodáctilos (NISP= 762; 36,85%) (Tabla 1). La categoría Artiodactyla incluye restos faunísticos óseos de cérvido y camélido, que al no tener características distintivas se asignaron a nivel de Orden. Asimismo, de acuerdo a la historia arqueofaunística de la Puna en general y la Puna de Salta en particular, se asume que estos restos pertenecientes a la categoría de artiodáctilos proporcionalmente podrían corresponder en su mayoría a camélidos y en menor medida a cérvidos (Elkin 1996;López 2008;Mondini y Elkin 2006;Yacobaccio 2017;Yacobaccio y Vilá 2016). Por otra parte, entre los taxones identificados que tienen menor representatividad en la muestra se observaa los chinchíllidos (NISP= 100; 4,84%) y a los roedores pequeños (NISP= 120; 5,8%). La representación de este último taxónen el registro arqueológico puede deberse a un posible ingreso tafonómico. De todas maneras, se considera que su influencia en la modificación del conjunto es limitada, dado que la mayoría de los indicadores arqueológicos (faunísticos y líticos) reflejan patrones recurrentes y concordantes con otros contextos similares a nivel macrorregional. En el caso del material lítico, esto se expresa en la presencia de las puntas triangulares apedunculadas como los instrumentos tiempo-sensitivos del Holoceno temprano (de amplia replicación en los Andes), sin evidencia de “intrusión” de materiales correspondientes a otras cronologías. A su vez, la fauna muestra una alta integridad general, con el agente humano como principal formador y modificador del conjunto.
Otros taxones fueron determinados en muy baja frecuencia. Este es el caso de un espécimen de cánido (NISP% = 0,05 %), posiblemente de un zorro puneño, y una falange tercera de cérvido (NISP= 0,05%). Estebajoporcentaje de especímenes de cérvido reflejaría una menor disponibilidad de este recurso para la caza (López 2008).
Tabla 1: Diversidad taxonómica considerando los especímenes de artiodáctilos y roedores pequeños en la capa F4.
En la figura 2, se puede observar la representación taxonómica al nivel de Familia. Al no incluirse las categorías taxonómicas por encima de este nivel, se identifica un mayor contraste entre los taxones. Esto eleva porcentualmente la representación de camélidos al 91,4% (Figura 2). En contraste, los chinchíllidos, cérvidos y cánidos son los taxones de menor y escasa representatividad en NISP% a nivel Familia (Figura 2). Esta baja frecuencia en el registro con respecto a los camélidos puede verse en que la sumatoria de los porcentajes de todos estos taxones es solo de 8,60%. Estos valores indican una predominancia casi absoluta de la familia Camelidae.
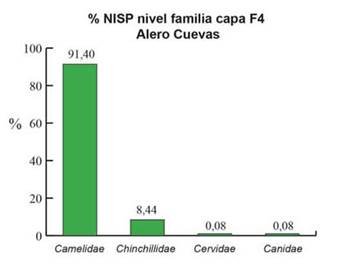
Figura 2: NISP porcentual del conjunto arqueofaunístico de la capa F4 del sitio Alero Cuevas considerando los taxones identificados a nivel Familia.
Representación de partes esqueletarias de camélidos
En la Tabla 2 se resume la totalidad de indicadores que fueron considerados en este trabajo para el análisis de las partes esqueletarias del taxón Camelidae. De acuerdo a los resultados, se puede observar la presencia de todos los elementos anatómicos que comprenden el esqueleto axial (NISP= 434; 40,1%)y apendicular (NISP= 650; 59,9%)en distintas frecuencias, siendo las costillas (NISP 141; 13%) y las falanges primeras (NISP= 162; 14,9%) los elementos más representativos en valores absolutos y porcentuales (Tabla 2). Otro indicador, el MNI, arrojó un valor de 10 individuos de camélidos. Con respecto a las partes esqueletarias medidas por MNE, se observa una baja representación de partes axiales(MNE= 74; 17,29%) y alta representación de partesapendiculares (MNE= 354; 82,71%). De acuerdo a estos valores, en el esqueleto axial se distinguen las costillas (MNE= 25; 5,8%) y las vértebras cervicales (MNE= 16; 3,7%). Asimismo, en los elementos apendiculares se detectauna mayor representatividad de las falanges primeras (MNE= 78; 18,2%), huesos carpianos (MNE= 52; 12,1%), falange segunda (MNE= 27; 6,3%) ymetapodio distal (MNE= 27; 6,3%) (Tabla 2).Por su parte, el MAU y MAU% muestran que las falanges primeras son las unidades anatómicas que tienen mayor representación, seguidas por el húmero y radioulna distal (Tabla 2). Del mismo modo, estas tendencias pueden distinguirse en la Figura 3 que muestra el agrupamiento de los elementos expresados en los porcentajes de las partesesqueletarias básicas (PEB).Estos valores indican una mayor representación de manos y pies (PEB= 37,38%) y falanges (PEB= 28,27%). Con menor representación porcentual se encuentran las patas delanteras (PEB= 10,75 %), patas traseras (PEB= 6,3%) ycolumna(PEB= 7,48%) (Figura 3).
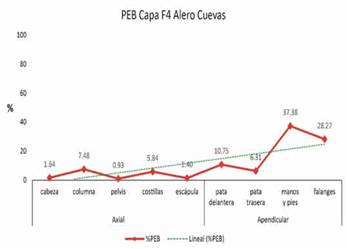
Figura 3: Agrupamiento del número mínimo de elementos en partes esqueletarias básicas (PEB) expresado en porcentajes con la línea de tendencia media.
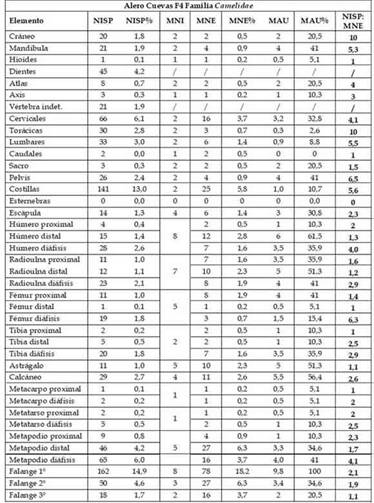
Tabla 2: Cuantificación de partes anatómicas de Camelidae en la capa F4 (9880-8504 AP), Alero Cuevas.
Perfil etario, índices de fragmentacióny utilidad e indicadores tafonómicos
De acuerdo al estado de fusión de los huesos de camélidos,elNISP% de especímenes fusionados es 57,7% y no fusionados 42,3%. Asimismo, el MNE dio por resultado 51,5% de fusionados y 42,5% de especímenes no fusionados.Por lo tanto, los resultados del NISP% y MNE% muestran mayor representación de especímenes fusionados. Por su parte, la determinación de edad dio por resultado los siguientes porcentajes: 57,5% maduros, 26,9% subadultos/inmaduros, y 15,5% neonatos. De esta manera, se distingue un alto porcentaje de la categoría adulto en comparación con subadultos y neonatos.
Por otra parte, los valores de la fragmentación total del conjunto dieron por resultado la razón NSP:NISP = 3,8. En el conjunto de camélidos, el valor de la fragmentación se expresa en la razón NISP:MNE = 2,5 (Tabla 2). De acuerdo a este último indicador en cada uno de los elementos anatómicos,en el esqueleto axial, los que presentan mayor fragmentación son la pelvis (6,5), las costillas (5,6) y las vértebras lumbares (5,5). En el esqueleto apendicular la mayor fragmentación se distingue en la diáfisis del fémur (6,3) y del húmero (4). Comparativamente, se observa que disminuyen los valores de fragmentación en los metatarsos, metacarpos, huesos tarsales, huesos carpiales y falanges (Tabla 2). De esta manera, se entiende que estos elementos habrían sido procesados con menor intensidad. En un nivel más agregativo, la media de la razón NISP:MNEfue variable entre el esqueleto axial (4,5) y apendicular (2). Estos valores de fragmentación indicarían una mayor intensidad de procesamiento en elementos de la parte axial y menor en los elementos que pertenecena la parte apendicular.
Para analizar el rendimiento de partes representadas en relación con la composiciónanatómica del conjunto, se correlacionó el MAU% con el índice de utilidad de carne, esto dio por resultado rho=-0,13598, p = 0,47369. De igual manera, se correlacionó el índice de médula rho=-0,17065,p= 0,36726 y el índice de secado de carne que dio una correlaciónnegativa moderada pero no significativa rho=-0,27548, p= 0,30175. A partir deestos resultados se entiende que el aprovechamiento de unidades anatómicas no presenta una selección de partes de acuerdo a su utilidad. En cambio, se estaría aprovechando la totalidad de las partes esqueletarias que constituyen a la estructura ósea de un camélido.
Por su parte, laFigura 4a muestra un alto contraste en los porcentajes de marcas antrópicas registradas en el esqueleto axial y apendicular, indicando una mayor cantidad de especímenes con marcas en las partes apendiculares (Figura 4a). Por otro lado, la Figura 4b discriminael porcentaje de especímenesde camélidos con distintas categorías de marcas antrópicas que fueron registradas (n= 237, 21,86% deltotal del conjunto). De esta manera, se reconoce que, de todas las categorías identificadas, las marcas de cortes son las que tienen mayor porcentaje (n = 169; 71,31%), luego los lascados y negativos de impacto, y en menor medida las marcas de machacado y raspado (Figura 4b).A nivel más específico, la Tabla 3 muestra la cantidad de unidades anatómicas con marcas distribuidasen cada categoría reconocida.En este sentido, puede identificarse quelas costillas (n= 48) y las falangesprimeras (n= 19) son las unidades principales que registran marcas de cortes (Tabla 3). En menor frecuencia, también se identificaronlas vértebras cervicales, diáfisis de húmeros, fémures,tibias y metapodios con marcas de cortes (Tabla 3). Los lascados, que es la siguiente categoría que presenta más cantidad de especímenes, se concentran puntualmente en la parte apendicular, especialmente en diáfisis de húmeros y falanges primeras. Por su parte, los
negativos de impacto tienen mayor distribución, pero tienden a concentrarse en las partes apendiculares (Tabla 3). Este registro de marcas estaría asociado a prácticas relacionadas con actividades de procesamiento para el consumo, patrones de descarne, desarticulación y extracción de la médula ósea.También se registraron escasas marcas antrópicas en los especímenes de chinchíllidos (0,01%).
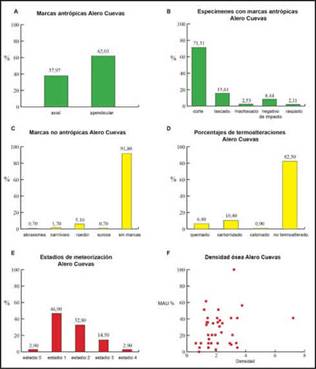
Figura 4: a) Porcentaje sobre el total de especímenes de camélidos con evidencias de marcas antrópicas identificadas en elementos del esqueleto axial y apendicular. b) Porcentaje sobre el total de los especímenes de camélidos con marcas antrópicas discriminadas por categorías. c) Porcentaje de especímenes de camélidos con marcas no antrópicas. d) Termoalteraciones en el conjunto de especímenes de camélidos. e) Estadios de meteorización del conjunto arqueofaunístico de la capa F4, Alero Cuevas. f) Gráfico de dispersión entre MAU% y la densidad ósea en Alero Cuevas capa F4.
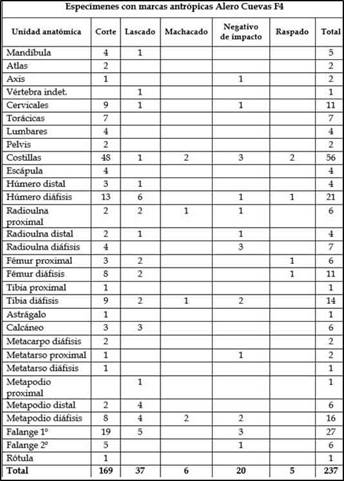
Tabla 3: Especímenes de camélidos con evidencia de marcas antrópicas en la capa F4 del sitio Alero Cuevas.
Por su parte,las marcas no antrópicastienenbaja frecuencia en el conjunto (8,20%) (Figura 4c).Entre ellas, predominan las marcas de roedor y en menor porcentaje las marcas de carnívoros (Figura 4c).En la figura 4d se observa que hay una baja representación de especímenes expuestos a termoalteraciones,un 10,40% de especímenes carbonizados y un 6,40% quemados (Figura 4d). Finalmente, la meteorización afectó de forma variable a los especímenes, con una predominancia de los estadios 1 (46,90%) y 2 (32,80%), aunque no es menor el porcentaje del estadio 3 (14,50%)(Figura 4e).A su vez, la densidad ósea y el MAU% muestran una correlación positiva moderada no significativa. Esto indicaría que la representación de partes esqueletarias no estuvo condicionada por la densidad global, tal como lo demuestra el resultado de la correlación con el MAU% (rho= 0, 24617, p= 0,13627) (Figura 4f).
Discusión y conclusiones
A nivel macrorregional la historia paleoambiental de la Puna indica que la distribución de parches de recursos durante el Holoceno temprano fue menos heterogénea que en la actualidad (Yacobaccio 2017;Yacobaccioet al. 2016). Esto significa que los diferentesgeoambienteshabrían estado menos separados y posiblemente más cercanos por la presencia de cuerpos de agua abundantes que permitieron la existencia de cubiertas vegetales extensas y permanentes (Morales 2011). Asimismo, se ha demostrado que en zonas más puntuales existió una variabilidad ambientalreconocida por las condiciones de mayor circulación de agua entre ca. 10600-9300 años AP y otros momentos de presencia de cuerpos de agua con menor circulación entre ca. 9300-8400 años AP (Oxman 2015;Yacobaccio y Morales 2005). De manera análoga, en las distintas escalas de análisis espacio-temporales se pueden reconocer la presencia de diversos cuerpos de agua con distintas dinámicas en el transcurso del Holoceno temprano que podrían haber generado diferentes cursos fluviales expandiendo los sectores fértiles. De esta manera, se interpreta que la distribución de animales sería más abundante y su disponibilidadmás frecuente y estable, pudiendo aprovecharse todos los espacios donde existían recursos. En este contexto ambiental, los camélidos, cérvidos, roedores y aves estarían distribuidos de manera más homogéneaen la Puna, formando parches de recursos con la suficiente biomasa para ser ocupados recurrentemente por sociedades cazadoras recolectoras tempranas (Muscio y López 2011;Yacobaccio 2017).
Específicamente, en la Puna de Salta, de acuerdo a los antecedentes paleoambientales de la Puna en general, se interpreta quelas quebradas de Pastos Grandes también habrían tenido una mayor presencia de cubierta vegetal, configurando parches de recursos más homogéneos y con mayor abundancia de animales silvestres. Concretamente, se reconoce que las quebradas de altura son sectores óptimos para la ocupación humana dentro del área debido a la presencia y circulación de agua permanente, pasturas, abundancia de animales para la caza y abrigos rocosos que aportan refugios seguros(López 2008). Estas característicashabrían estado disponibles para las sociedades cazadoras recolectoras del Holoceno temprano. En este sentido, la alta frecuencia de material faunístico y lítico demuestra la alta intensidad delas ocupaciones humanas en Alero Cuevas durante este período (López 2008; Muscio y López 2016;Restifo 2011 2012, 2013). Por consiguiente, en el marco de una dinámica de alta movilidad humana con uso de determinados parches, sumado a la recurrencia constante de ocupaciones, puede plantearse que este sector de quebradas altas constituyó un espacio de aprovechamiento óptimo en términos de calidad global del ambiente (Muscio y López 2011; Yacobaccio 2017).
Concretamente, para la dispersión y ocupación humana de la Puna habría sido clave la disponibilidad de recursos de alto ranquin comoson los camélidos (Muscio y López 2016).En la Puna de Salta y particularmente en las quebradas de Pastos Grandes,la abundancia y disponibilidad de camélidos es fundamental para comprender la intensidad y recurrencia de las ocupaciones humanas en Alero Cuevas, principalmente porque su procesamiento permite obtener recursos alimenticios como carne, médula, grasa y utilitarios materiales como el cuero, tendones y lana para tecnofacturas(MengoniGoñalonsy Yacobaccio 2006;Muscio y López 2011; Olivera 1997). Asimismo, esta importancia de los camélidos se extiende a nivel regional y macrorregional (Olivera 1997;Yacobaccio 2012).
La evidenciaarqueofaunística del sitio Alero Cuevas indica que los camélidos fueron el principal recurso alimenticio animal en el Holoceno temprano. Los resultados de la diversidad taxonómica de la capa F4 (ca. 9800-8500 años AP) muestran un predominio casi absoluto de estos recursos por encima del 90% en NISP. Este resultado refleja un aprovechamiento alimenticio concentrado en camélidos en contraste con fauna pequeña. Con respecto a esto, debe destacarse que en el presente en la quebrada de las Cuevas, donde se encuentra el sitio,hay abundancia de vizcachas en los roquedales. De manera análoga, puede plantearse la disponibilidad de estos recursos en las cercanías del alero durante el Holoceno temprano, pero aun así su frecuencia en los conjuntos arqueofaunísticos es reducida. Esta particularidad podría estar relacionada con la alta disponibilidad de los recursos de mayor rendimiento (camélidos) en las quebradas, lo cual no haría necesario recurrir a fauna de menor calidad (chinchíllidos). De esta manera, el aprovechamiento de camélidos explicaríael uso recurrente del alero y condice con lo esperado para ocupaciones humanas estables a lo largo del Holoceno temprano. Asimismo, de acuerdo al estado de fusión de los huesos y la determinación de edad,los resultados sugieren un patrón centrado en animales adultos, mientras que la presencia de neonatos indicaría una probable ocupación estacional del sitio.
Por su parte, la figura 5 muestra el NISP% de taxones a nivel familia en una escala macrorregional y puede observarse el contraste entre la fauna de alto retorno energético (camélidos) con la fauna pequeña de menor rendimiento (Figura 5).Esta comparación indica una alta diversidad relativa de animales silvestres explotados durante el Holoceno temprano, considerando el número y la representaciónde taxa registrada en diferentes sitios arqueológicos. Los resultados de Alero Cuevasindican una menor diversidad relativa, debido a la concentración en el consumo de camélidos, en contraste con otros sitios que presentan mayor variabilidad de taxonesaprovechados. Para el Holoceno tempranose ha sugerido un comportamiento macrorregional fragmentado en el consumo y explotación de animales silvestres (Yacobaccio 2017). En consecuencia, las tendencias en Alero Cuevas se relacionan más con Huachichocana III, Quebrada Seca 3 y Punta de la Peña 4 (Elkin 1996;Urquiza y Aschero 2014; Yacobaccio 2001) quecon sitios como Pintoscayoc 1 o Inca Cueva 4 (Elkin y Rosenfeld 2001; Hernández Llosas 2000; Yacobaccio 2001), estos últimos con alta presencia de roedores. También se observa un contraste con los sitios del norte de Chile donde hay mayor aprovechamiento de fauna pequeña. Asimismo, se relaciona con la evidencia considerable de camélidos en los sitios de Tuina1 y 5 (Núñez et al. 2005). Estas similitudes y diferencias podrían deberse a variaciones locales en la disponibilidad de fauna en general y de camélidos en particular. Más precisamente,se debe profundizar el análisis comparativo de diversos ambientes locales de la Puna argentina y el norte de Chile, cuyas particularidades pudieron haber afectado diferencialmente la disponibilidad de recursos, principalmente los camélidos.
Por otra parte, a nivel más específico, se interpreta que existió un aprovechamiento completo de los camélidos como presas, así lo indican los altos porcentajes del esqueleto axial y apendicular, sumado a la presencia de todas las partes que componen el esqueleto de los camélidos. Asimismo, de todas las unidades identificadas se destaca principalmente la predominanciade falanges primeras y costillas. De igual manera, a nivel más agregativo se observó la mayor presencia de patas delanteras y traseras. Sin embargo, se identificó un menor estado de fragmentación en los huesos que componen las patas delanteras y traseras que en los elementos que componen la parte axial. Esto estaría indicando que la fragmentación fue más intensa en los huesos de la parte axial. Por otra parte, se identificaron mayor cantidad de marcas en la parte apendicular, principalmentemarcas de corte en las falanges. Esta evidencia es coincidente con la alta presencia de estos elementos anatómicos en el registro y se asocia con las prácticas de procesamiento y consumo (descarne, desarticulación y extracción de médula). Por lo tanto, estos patrones indican que las partes apendiculares tienen mayor presencia en el registro, mayor cantidad de marcas antrópicas y un índice de fragmentación menor que las partes esqueletarias axiales. En este sentido, se interpreta que la concentración del aprovechamiento alimenticio estaría orientado con mayor intensidad de procesamiento a todos los sectores que componen alto contenido de carne, masa muscular y vísceras. También se estarían procesando y aprovechandolas unidades anatómicas que tienen menor cantidad de carney mayor rendimiento de médula y grasa ósea (huesos de patas delanteras y traseras), pero con una baja intensidad que dejaría una menor fragmentación en este registro. En trabajos anteriores, se han presentado indicadores de fragmentación que muestran una menor intensidad de procesamiento en estos elementos del conjunto de arqueofaunas de F4 (López y Orsi 2019).
Si bien se espera que el consumo completo de una presa para la alimentación se oriente a la obtención de carne y masa muscular, y al acceso a médula y grasa ósea con una alta fragmentación de las partes esqueletarias apendiculares (Wolverton 2002), en el caso de la capa F4 de Alero Cuevas, los resultados no indican una alta intensidad de fragmentación y procesamiento. Por el contrario, la intensidad de procesamiento parece haber aumentado en el conjunto del Holoceno medio final (ver López y Orsi 2019;Orsi 2018;Orsi y López 2019). Se ha interpretado que estos cambios se habrían relacionado con mayores presiones ecológicas y sociales que llevaron al aumento de la demanda energética en ese período, al contrario delo ocurrido en el Holoceno temprano (López y Orsi 2019).

Figura 5: Gráfico de comparación macrorregional del NISP% de los taxones principales registrados en sitios con fechados del Holoceno temprano de la Puna Jujuy, Salta, Catamarca y Atacama en el norte de Chile
Por último, es importante destacar que la alta presencia de camélidos en el conjunto del Holoceno temprano de Alero Cuevas es coincidente con un ambiente que tendría una mayor disponibilidad y oportunidad de encuentro de estos recursos animales. En consecuencia, la concentración en el aprovechamiento de camélidos es esperable en estos contextos.
Agradecimientos: Agradezco a CONICET, al equipo editorial de la revista, a los evaluadores por lascorrecciones para mejorar la calidad del trabajo y a mi director Dr. Gabriel López por los comentarios y sugerencias.

