











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Estudios que aborden la reproducción de las plantas y su relación con los insectos polinizadores contribuyen a un mejor entendimiento del funcionamiento de los ecosistemas regionales ya que las interacciones planta-polinizadores son críticas no sólo para el flujo génico en la población de plantas (Hall et al., 1996), sino también para el mantenimiento de la biodiversidad y la integridad de los ecosistemas (Aizen & Vázquez, 2006; Kearns & Innouye, 1997). Además, esta información es básica para el desarrollo de estrategias de conservación tanto de la diversidad genética como taxonómica de una comunidad biológica (Tadey et al., 2009).
Chuquiraga avellanedae Lorentz (Asteraceae) es una especie endémica de la estepa patagónica y representa un componente fundamental en los ecosistemas terrestres de la región. Habita desde el sur de la provincia de Mendoza hasta el norte de Santa Cruz (Ezcurra, 1985). Es dominante en la Provincia Fitogeográfica Patagónica (Bisigato et al., 2016) y posee la capacidad de modelar el ambiente al fijar el suelo (Feijóo, 2006). Es utilizada como refugio, sitio de nidificación y alimentación de diversos taxones de artrópodos (Martínez et al., 2021) y posee propiedades de interés medicinal (Forcone, 2004; Tapparaci & González, 2009). A pesar de la relevancia de esta especie arbustiva, se desconocen el grado de autoincompatibilidad, la dependencia a los agentes polinizadores y la composición del ensamble de visitantes florales.
Considerando que es un arbusto longevo con flores hermafroditas, y que una especie congénere (C. oppositifolia D. Don) es autoincompatible (Muñoz & Arroyo, 2004), se espera que C. avellanedae presente autoincompatibilidad. Además, los caracteres florales de C. avellanedae (i.e. tamaño, color y oferta simultánea de flores) sugieren que el traslado del polen es realizado por insectos (Tadey et al., 2009; Torres & Galetto, 2008). Por otro lado, el período de floración (Campanella & Bertiller, 2008) coincide con los fuertes vientos del norte predominantes en el verano (6 m/s velocidad media máxima) (Palacio et al., 2014). Entonces, considerando que el viento es un componente característico en la Patagonia (Paruelo et al., 1998) y que la polinización por viento puede ser más importante en sitios abiertos que en los hábitats cerrados y boscosos (Medan & Devoto, 2017), es probable que, además de los insectos, el viento tenga un papel importante en la polinización de C. avellanedae. Este fenómeno que presentan las especies vegetales polinizadas tanto por los insectos como por el viento se denomina ambofilia (Culley et al., 2002; Rech et al., 2016). Por ello, los objetivos de este estudio fueron determinar el grado de autoincompatibilidad de C. avellanedae y evaluar si es una especie ambófila.
Materiales y Métodos
Area de estudio y especie focal
El trabajo se realizó en la proximidad de la ciudad de Puerto Madryn, sitio ubicado en el ecotono entre las Provincias Fitogeográficas del Monte y Patagónica (-42,84 S; -65,02 O). En el ecotono coexisten comunidades vegetales, donde se destacan la de Larrea divaricada Cav. y Stipa spp., típica del Monte Austral y la de C. avellanedae, característica de la Provincia Fitogeográfica Patagónica (Bisigato et al., 2016). El clima es árido, templado y ventoso. La temperatura media anual es 13,5 °C y la precipitación media anual es 233,8 mm, con alta variación entre años y sin un patrón claro de estacionalidad (Casalini, 2016).
Chuquiraga avellanedae es un arbusto siempre-verde de 0,5 a 1,5 m de altura de estructura ramosa y forma hemisférica. Las flores son tubulosas, amarillas y hermafroditas, se hallan reunidas de 8 a 15 en capítulos sésiles. Éstos se disponen de 1 a 3 en el extremo de los tallos o las axilas de las hojas. Las flores son amarillas, con papus pardusco tan largo como la corola. Es una especie protándrica (observación personal) y el polen tiene un diámetro de 30 pm (Forcone, 2008). Su fruto es un aquenio, el cual contiene una sola semilla (Correa, 1971). El período de floración se extiende desde Noviembre hasta Marzo (Kropfl et al., 2012).
Tipo de sistema reproductivo y dependencia a los agentes polinizadores
Con el fin de caracterizar el sistema reproductivo de C. avellanedae se realizó un experimento de cruces manuales. Se seleccionaron 30 individuos de C. avellanedae al comienzo del período reproductivo (Noviembre - Diciembre 2018). En cada arbusto se escogieron 5 ramas al azar y se retiraron algunos capítulos hasta alcanzar un único capítulo por rama. A cada rama se le asignó uno de los siguientes tratamientos (n=30 capítulos por tratamiento) (Tadey, 2007): 1) autopolinización forzada: flores embolsadas en pimpollo con friselina (excluye al polen) y polinizadas manualmente con polen de la misma planta, 2) autopolinización espontánea: flores embolsadas en pimpollo con friselina y sin intervención manual, 3) polinización cruzada: flores embolsadas en pimpollo con friselina y polinizadas manualmente con una mezcla de polen con-específico proveniente de otras plantas, 4) polinización por viento: flores embolsadas en pimpollo con voile de 0,2 mm de apertura de malla (excluye a los insectos y permite el paso del polen bajo la acción del viento) y sin intervención manual y 5) control: se dejaron flores sin embolsar expuestas a los agentes polinizadores naturales y sin intervención manual.
La intervención manual se realizó cuando los estigmas se encontraron abiertos (Kearns & Innouye, 1993). La superficie de los estigmas se cubrió de polen pincelando suavemente con las anteras (Muñoz & Arroyo, 2006). En cada capítulo se realizaron dos intervenciones manuales, la primera cuando se observó cuatro flores del capítulo con los estigmas bifurcados y la segunda luego de dos días posteriores a la primera intervención. Realizados los cruces, las flores fueron monitoreadas hasta la formación de los aquenios. En el caso del tratamiento control, las flores se embolsaron con friselina una vez pasado el período de receptividad de los estigmas florales para evitar pérdidas por dispersión. Los capítulos fueron colectados en Marzo de 2019, donde se contabilizó la cantidad de flores y de aquenios por capítulo. Además, se calculó el índice de auto-compatibilidad (ISI), que resulta de la razón entre la producción de frutos por autopolinización forzada (tratamiento 1) y la producción de frutos por polinización cruzada (tratamiento 3). Este índice va de 0 (totalmente autoincompatible) a 1 (autocompatible). Se considera a la planta como autoincompatible con valores del ISI por debajo de 0,2 (Zapata & Arroyo, 1978).
Ensamble de insectos polinizadores
Para describir el ensamble de insectos polinizadores se realizaron observaciones directas de los insectos que visitaron a individuos de C. avellanedae durante el período de floración (Diciembre - Febrero) en las temporadas 2017/18 y 2018/19. Los muestreos se realizaron en intervalos de 10 min por planta (Muñoz & Arroyo, 2004) en el período de mayor actividad de los insectos, entre las 11:00 y 14:00 hs (Tadey et al., 2009). Durante ambas temporadas se realizaron 109 períodos de observación que equivalen a un total de 18,2 horas de observación directa. Se consideraron como polinizadores a aquellos insectos que tuvieron contacto con los órganos sexuales de las flores. Se registró a cada individuo y la cantidad de capítulos visitados. Además, cuando fue factible, se realizó la captura manual de los polinizadores. Los ejemplares fueron determinados al nivel taxonómico más bajo posible por medio de bibliografía especializada (Nieves-Aldrey et al., 2006; Pyrcz et al., 2016; Triplehorn et al., 2005). Finalmente, se estimó la abundancia relativa y la frecuencia de visitas de los distintos taxones que componen el ensamble de polinizadores de C. avellanedae. Los insectos fueron depositados en la colección entomológica del Instituto Patagónico para el Estudio de los Ecosistemas Continentales (IPEEC-CONICET).
Análisis estadísticos
Para evaluar la variación entre los efectos de los tratamientos se utilizaron modelos lineales generalizados mixtos (GLMM). Se consideró una variable respuesta binomial, con el número de aquenios producidos como “número de éxitos” y el número de flores vacías como “número de fracasos”. Se incluyó a los tipos de cruzamiento como efecto fijo y a los individuos de C. avellanedae como efecto aleatorio (bloques), controlando de esta manera la variabilidad asociada a cada individuo al que se le aplicaron los tratamientos (Zuur et al., 2009). Se utilizó una distribución de error binomial (función link = logit) y la validación del modelo se realizó a través de la inspección de los gráficos de residuales. Finalmente, se hicieron contrastes de Tukey (a posteriori) para analizar la variación entre tratamientos. Los análisis se realizaron utilizando los paquetes lme4 (Bates et al., 2015), multcomp (Hothorn et al., 2008) y DHARMa (Hartig, 2020) del software R (R Development Core Team, 2020). La rutina del análisis estadístico se encuentra en el material suplementario.
Resultados
Tipo de sistema reproductivo y dependencia a los agentes polinizadores
La cantidad de semillas producidas por capítulo varió entre los tratamientos (x2=43,345; p<0,0001; g.l.=3; Tabla A, material suplementario). En el tratamiento de autopolinización espontánea (n=27) no hubo producción de semillas, mientras que para la autopolinización forzada (n=26) y la polinización por viento (n=28) se obtuvieron valores bajos (Fig. 1). La cantidad de semillas producidas por capítulo fue significativamente mayor en los tratamientos control (n=30) y de polinización cruzada (n=27) (Fig. 1; Tabla B, material suplementario). El número de flores por capítulo fue de 9,78 ± 0,13 (media y error estándar).
El índice de auto-compatibilidad ISI fue de 0,14, por lo cual el sistema reproductivo de C. avellanedae se clasificó como autoincompatible.
Ensamble de insectos polinizadores
El ensamble estuvo compuesto por 49 morfo-especies/especies pertenecientes a seis ordenes (Tabla 1). Hymenoptera y Diptera fueron los ordenes que presentaron la mayor riqueza relativa de morfo-especies/especies y mayor registro de visitas (Tabla 1).
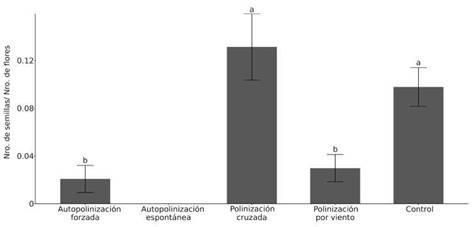
Fig. 1: Media y error estándar del número de semillas por flores del capítulo producidas en cada tratamiento. Letras distintas indican variación entre los tipos de cruzamiento.
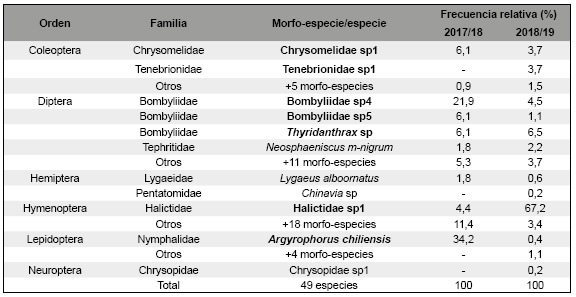
Tabla 1: Polinizadores registrados durante los períodos de observación directa (N=578). En negrita se indican las siete morfo-especies/especies más frecuentes. En la categoría “otros” se agruparon los polinizadores que presentaron menos del 1% del total de las visitas. Las temporadas en la que no hubo registros se simboliza con
Durante las dos temporadas se registraron 578 visitas. Argyrophorus chiliensis Bryk (Lepidoptera: Nymphalidae) fue el polinizador más frecuente en la primera temporada, en cambio para la segunda temporada lo fue Halictidae sp1 (Tabla 1).
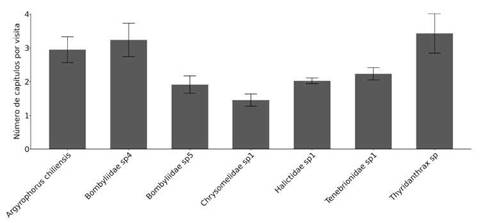
Entre los taxones más frecuentes, la cantidad promedio de capítulos visitados por visita varió entre 1,46 y 3,42 correspondiendo a Chrysomelidae sp1 y Thyridanthrax sp (Diptera:
Bombyliidae) respectivamente. Chrysomelidae sp1 visitó en promedio 1,46 capítulos por visita (rango: 1-2 capítulos). Bombyliidae sp5 visitó en promedio 1,90 capítulos por visita (rango: 1-12). Halictidae sp1 visitó en promedio 2,03 capítulos por visita (rango: 1-13). Tenebrionidae sp1 visitó en promedio 2,24 capítulos por visita (rango: 1-2). A chiliensis visitó en promedio 2,95 capítulos por visita (rango: 1-12). Bombyliidae sp4 visitó en promedio 3,24 capítulos por visita (rango: 1-15). Finalmente, Thyridanthrax sp visitó un promedio de 3,42 capítulos por visita (rango: 1-14) (Fig. 2).
Discusión
Los resultados encontrados indican que C. avellanedae es autoincompatible. Por lo tanto, este arbusto necesita la presencia de otros individuos y de agentes polinizadores para reproducirse sexualmente de manera exitosa. Este resultado concuerda con el descripto para otra especie cercanamente emparentada al nivel taxonómico de género, C. oppositifolia, la cual es autoincompatible (Muñoz & Arroyo, 2004). En este sentido, la polinización cruzada es considerada ventajosa respecto de la autofecundación porque produce una progenie genéticamente más variable y evita la depresión por endogamia, incrementando el potencial de adaptación ante eventuales cambios ambientales (Barrett, 2003).
En este trabajo se evidenció que la población de C. avellanedae estudiada depende principalmente de los insectos para reproducirse. En América del Sur se ha descripto polinización por insectos y colibríes para el género Chuquiraga (Abrahamczyk et al, 2017; Ezcurra, 2002). En consonancia con estos resultados, se describe que las especies del género que habitan en las Provincias Fitogeográficas del Monte y Patagónica son polinizadas por insectos (Ezcurra, 2002). Además, otros estudios mencionan que las especies de zonas áridas en Argentina también son polinizadas por insectos (Debandi et al., 2002; Tadey et al., 2009; Tadey, 2011). Con respecto a la ambofilia estos resultados sugieren que el viento no juega un rol importante como agente polinizador de C. avellanedae. Sin embargo, descartar la posibilidad de que presente ambofilia a lo largo de su rango de distribución (Mendoza - Santa Cruz) podría ser apresurado, ya que existen registros de variación entre polinización exclusiva por insectos y ambofilia para otro arbusto de Patagonia, Discaria chacaye (G. Don) Tortosa (Medan & Devoto, 2017).
Las flores de C. avellanedae fueron visitadas por 49 morfo-especies pertenecientes a 6 ordenes de insectos, por lo que el ensamble de polinizadores se considera generalista. También, se observó variabilidad interanual en la composición específica y en la abundancia relativa entre los taxones que componen el ensamble. Estos resultados coinciden con lo encontrado en otros estudios (Chacoff et al., 2018; Vázquez & Simberloff, 2004) donde se destaca que las interacciones son altamente dinámicas y varían mucho en el tiempo (Poisot et al., 2015). Frente a esta variabilidad temporal, la presencia de un ensamble generalista resultaría ventajosa. En este sentido, se ha encontrado que especies de polinizadores funcionalmente redundantes favorecen el éxito reproductivo de las plantas ya que, cuando alguno de los polinizadores está ausente o la abundancia es relativamente baja, la función como polinizador es reemplazada por otra especie del ensamble (Balvanera et al., 2005, 2006; Fenster et al., 2004; Morris, 2003).
Este trabajo aporta información novedosa sobre el sistema reproductivo y los visitantes florales de C. avellanedae, una especie endémica de la Patagonia y relevante en el ecosistema. Lo hallado indicaría que este arbusto es autoincompatible y depende principalmente de los insectos polinizadores para reproducirse de manera exitosa. También se encontró que C. avellanedae es una fuente de recursos para una elevada cantidad de insectos que visitan sus flores, y, por lo tanto, una especie vegetal relevante para los ensambles de polinizadores de la zona. Además, para la misma región, se ha encontrado que C. avellanedae es un hábitat importante para otros grupos funcionales de insectos (e.g. detritívoros y granívoros) (Dellape & Cheli, 2007; Martínez et al., 2021) sugiriendo que la interacción C. avellanedae-insectos representa un componente relevante en procesos y servicios ecosistémicos claves como el ciclado de nutrientes y la polinización.
Contribución de los autoresTodos los autores han realizado conjuntamente y a partes iguales la colecta de datos, su interpretación y redacción del manuscrito.
Agradecimientos
Al Consejo Nacional de Investigaciones Científicas y Técnica (PIP 11220170100981 y PUE-IPEEC-2016 22920160100044) y a la Universidad Nacional de la Patagonia San Juan Bosco (PI-1491) por proveer los fondos. A la Dirección de Flora y Fauna Silvestre de la Provincia de Chubut por los permisos de colecta (Disposición N° 09/18). A la Dr. A. Zapata y al Dr. M. Devoto por la ayuda en la determinación de los especímenes.

