











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El pelo es una estructura de gran importancia ya que forma parte de la identidad, características étnicas y la imagen de cada persona. Los seres humanos cuentanen promedio con 100.000 cabellos cada uno producido por un apéndiceespecializado de la piel denominado folículo piloso el cual es formado a partir deltercer mes de vida embrionaria derivado de la placoda ectodérmica.1-3. Cadafolículo piloso presenta diversas características morfológicas e inmunológicasque permiten el desarrollo y crecimiento normal de los tallos pilosos. Al presentarse alguna alteración en los mecanismos reguladores del ciclo fisiológico del folículo piloso se impide el crecimiento del tallo piloso y la consecuente aparición de áreas alopécicas.
Reporte de casos
Caso 1:Paciente femenina de 23 años estudiante de arquitectura, con cuadro clínico de 3 meses de evolución consistente en aumento decaída decabello en algunas zonas del cuero cabelludo, niega antecedentes médicos de importancia, refiere notar aumento de la caída posterior a evaluaciones finales del semestre. Al examen físico se evidencia en región coronal área alopécica numular de 2 cm de diámetro aproximadamente con escasos tallos pilosos en su superficie, algunos miniaturizados y con disminución en su grosor. (Ver figura 1a)Test de pilotracción positivo sinpoliosis. Se toma biopsia de área alopécica donde se evidencia infiltrado linfocitario peribulbar confirmando el diagnóstico de alopecia areata (Ver figura 1b). Tomando en cuenta los hallazgos clínicos e histológicos se inicia manejo con infiltraciones de triamcinolona, presentando mejoría y repoblamiento a los dos meses de tratamiento
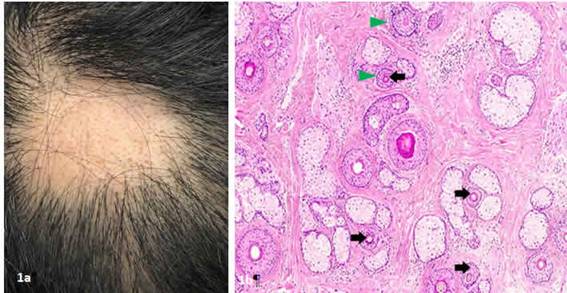
Figura 1: a. Área alopécica en región coronal con escasos tallos pilosos y pelos miniaturizados. Figura 1b. Corte transversal. Múltiples folículos pilosos miniaturizados (flechas negras) asociados a leve infiltrado linfocitario peribulbar (triángulos verdes) Hematoxilina-eosina 10x (Fotografía Cortesía Dr. Jairo Alfonso Sierra Avedaño y Dr. Nabor Gallo Gómez)
Caso 2: Paciente femenina de 17 años con cuadro clínico de 5 meses de evolución consistente en aparición de zonas sin cabello que han aumentado de tamaño y persisten a pesar de automedicarse betametasona en crema dos veces al día por un mes (Ver figura 2a). Sin antecedentes médicos de importancia. Al examen físico se evidencia en región biparietal áreas alopécicas numulares que confluyen con escasos tallos pilosos en su superficie, test de pilotracción positivo. Se inicia manejo con infiltraciones detriamcinolona y minoxidil en espuma, presentando repoblamiento a los tres meses.
Caso 3: Paciente femenina de 34 años con cuadro clínico de 6 meses de evolución consistente en caída de cabello progresiva posterior a quedar desempleada, niega otra sintomatología, niega antecedentes de importancia. Al examen físico múltiples áreas alopécicas numulares en región temporoparietal y disminución de la población pilosa, a la dermatoscopia se observan puntos negros, pelos afinados y pelos en signo de admiración. (Ver figura 2b) en cuarto dedo mano derecha y segundo dedo de mano izquierda múltiples pits en lámina ungueal(ver figura 2c).
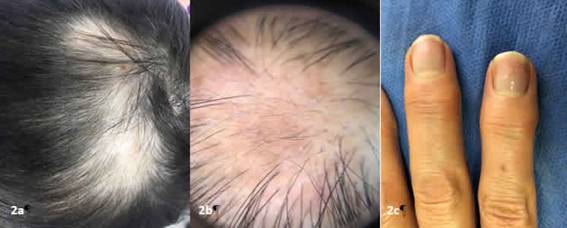
Figura 2:a Hallazgos al examen físico del caso 2, Figura 2b. Hallazgos dermatoscópicos del caso 3, Figura 3c. Múltiples pits ungueales en paciente del caso 3
Inmunobiología del folículo piloso
El folículo piloso es un sitio de gran interacción inmune, al realizarse estudios histológicos es posible evidenciar aumento en la cantidad de células presentadoras de antígeno en el segmento superior del folículo con disminución en su número a medida que se distribuyen por el segmento inferior.4,5 Esta distribución particular, predominante en la fase anágena, le concede al folículo pilosolas propiedades de una estructura con privilegio inmune(PI). 6,8
Privilegio inmune
Son varios los mecanismos que permiten evitar un ataque citotóxico sobre las células de algunos tejidos, la función del privilegio inmune esasegurar un ambiente óptimo para la realización del ciclo piloso y secuestrar autoantígenos asociados a melanocitos generando un mecanismo de protección ante una potencial respuesta auto agresiva nociva en la fase anágeno o hacia la melanogénesis.4,9
En el folículo piloso esta habilidad protectora es dinámica, ya que se presenta solo en el epitelio proximal del tallo piloso en fase de anágeno y se da, gracias a varios mecanismos que logran mantener el equilibrio funcional.10,11
Algunos de estos mecanismos sonla disminución o ausencia en la expresión del complejo mayor de histocompatibilidad de clase I, que secuestra autoantígenos en los tejidos y dificulta su presentación a linfocitos TCD8+ autoreactivos6, la producción local de inmunosupresores como el factor transformante de crecimiento beta 1 (TGF-B1), IL-10 y hormona estimulante de melanocitos (MSH-alfa)9,la ausencia de vasos linfáticos12,el establecimiento de una matriz extracelular de barrera que impide el paso de células del sistema inmunológico13 y la expresión de moléculas del MHC clase Ib y de Fas ligando (FasL, CD95l) para eliminar las células T autoreactivas.7
El inmunoprivilegio del bulbo solo existe en fase anágena en gran parte, ya que la mayoría de queratinocitos CMH clase Ia/B2 microglobulina negativos se eliminan vía apoptosis durante el catágeno7.
Complejo Mayor de Histocompatibilidad
Los locus genéticos de la región CMH se agrupan en tres clases (I-III), las moléculas del CMH I se dividen en clásicas (CMHIa) y no clásicas (CMHIb). Las moléculas clásicas a su vez se subdividen en HLA-A, HLA-B y HLA-C en seres humanos y se encargan de presentar péptidos antigénicos endógenos a linfocitos T CD8+. Las moléculas no clásicas se subdividen en HLA-E, HLA-F, HLA-G y HLA-H y tienen como función inhibir las células Natural Killer (NK). A pesar de que el CMH I se expresa en células nucleadas, existe una carencia de expresión de moléculas de clase I clásicas en aquellos tejidos inmunoprivilegiados. Las moléculas del CMH clase II tienen como función presentar péptidos antigénicos exógenos a linfocitos TCD4+, sin embargo en los tejidos inmunoprivilegiados su expresión se encuentra reducida.9
Así como se encuentra disminuida la expresión de moléculas del CMH en la porción proximal del folículo piloso, también existe una reducción en la expresión de moléculas asociadas a esta vía de presentación, tales como el transportador asociado con el procesamiento de antígeno (TAP) y la subunidad B2 microglobulina de las moléculas del CMH I. Para efectuar la presentación de antígenos a Linfocitos T CD8+ se requiere de TAP ( subunidades TAP1 y TAP2) que ejerce su función al interior del retículo endoplásmico rugoso, donde se une a la molécula de clase I compuesta por la subunidad β2 microglobulina y la subunidad a, para luego plegarse, liberarse y expresarse en la superficie celular. 9,14
Linfocitos T y células dendríticas
En condiciones normales los linfocitos TCD4+ se encuentran disminuidos en los folículos pilosos en fase de anágeno, y en la región proximal del folículo piloso se puede apreciar una ausencia de TCD8+ y células CD1a+, con reducción del número de células de Langerhans.9,14 En la alopecia areata se puede observar un infiltrado de TCD8+ y TCD4+ alrededor del folículo piloso en fases tempranas y una disminución del infiltrado de TCD4+ pero persistencia o ligero aumento en la cantidad de TCD8+ con inducción de respuesta de tipo TH1 autoinmune en fase anágena.15
Mastocitos
Los mastocitos son células efectoras de respuesta inmediata que pueden liberar mediadores pro-inflamatorios e interactuar con linfocitos por medio de la expresión de CMH clase I , en alopecia areata se ve aumentada la cantidad de mastocitos en comparación con áreas sin afectación, lo que sugiere que presentan un papel co-estimulador de señales para linfocitos T CD8. Los mastocitos también pueden liberar mediadores proinflamatorios sin generar degranulación tal como IL-6 y factor de crecimiento endotelial (VEGF). 14
Natural Killer
Las células NK poseen receptores inhibitorios que reconocen las moléculas de CMH I que se expresan en células sanas del individuo. En estado activador pueden reconocer estructuras presentes en célulassusceptiblesasí como de células normales, esta influencia predomina cuando se reconocen las moléculas del CMH clase I. Las células NK no se encuentran usualmente alrededor de los folículos pilosos en fase anágena, en cambio se pueden apreciar infiltrando el folículo pilosos en la alopecia areata (AA). 6,9,12
Los folículos pilosos normales en anágeno logran escapar del ataque de células NK mediante el aumento de receptores y factores de inhibición que suprimen la actividad de los mismos, tales como el factor inhibidor de la migración de macrófagos (MIF). Otro mecanismo de evasión del ataque por parte los NK es reduciendo la recepción de señales de estimulación mediante la reducción en la expresión del receptor activador NKG2D y de su ligando MICA. En pacientes con AA se encuentra una generalizada inmnorreactividad para el ligando MICA y una mayor expresión en la vaina radicular externa proximal, la papila dérmica y la vaina de tejido conectivo del folículo piloso, sugiriendo una disminución en la capacidad para la supresión de la función no deseada en células NK en pacientes con alopecia areata. 6,12
Drenaje linfático y matriz extracelular
La ausencia de vasos linfáticos de drenaje y la presencia de una barrera de matriz extracelular compuesta por tejido conectivo alrededor del folículo piloso impiden el traslado de células del sistema inmune a estas estructuras favoreciendo el carácter de inmunoprivilegio en esta zona. 4
Otros inmunosupresores locales
Se ha encontrado expresión de una molécula inmunosupresora capaz de disminuir la expresión ectópica de las moléculas del MHC I, que se denomina MSH-α, esta se encuentra en losqueratinocitos de la VRE y la matriz durante lafase anágena. También existen moléculas como TGF-β que es uno de los inmunosupresores más potentes y es expresado durante el anágeno tardío y el inicio de la fase catágeno en las células de la VRE.Este factor induce el catágeno, mientras que otras moléculas como IGF-I, prolongan el anágeno e inhibe el catágeno, llevando a pensar que las moléculas IGF-I, TGF-β 1 y MSH-α son reclutados por el folículo piloso en anágeno para efectuar un papel protector del inmunoprivilegio en esta zona. 9,14
Colapso del privilegio inmune y alopecia areata
Inicialmente se puede evidenciar en la alopecia areata una perdida focal de cabello en piel no inflamada que puede llegar a progresar a perdida universal del cabello en algunos pacientes. Histologicamente se evidencia un denso infiltrado inflamatorio autoagresivo alrededor del bulbo piloso que causa la terminación prematura de la fase de anágeno y el inicio temprano de catágeno produciendo distrofia mayor del folículo piloso. Estos cambios inducen un colapso del PI generando una disrupción en el ciclo y el crecimiento del pelo. 6,10
El PI puede colapsar por múltiples causas como infecciones, estrés emocional, daño o trauma, produciendo un aumento en la expresión de IFN-gamma que incrementa las moléculas CMH de clase I en la matriz del pelo. Al existir un fallo en el IP de los folículos en anágeno existe una actividad insuficiente de los inmunosupresores locales y las actividades represoras de NK que inducen un ataque mediado por células TCD8+ al epitelio del folículo16-18
El carácter reversible de este proceso ha permitido la investigación de nuevas moléculas como los inhibidores de la JAK/STAT, que muestran su efectividad en el manejo de la AA por medio del bloqueo de la señalización mediada por INF-gamma inhibiendo la activación tanto de células TCD8+ como NK6,7
La activación de células TCD8+ genera aumento de la respuesta inmune y destrucción celular que produce liberación de ácidos nucleicos y autoantígenos que activan células presentadoras de antígenos y promueven el reclutamiento de más linfocitos. (Ver figura 3) Se genera una infiltración celular inflamatoria intra y perinfundibular que promueve la formación de un estadio anágeno distrófico en estadios tempranos de la alopecia areata y en estadios más tardíos una fase telogénica inactiva crónica. 6,14,19,20
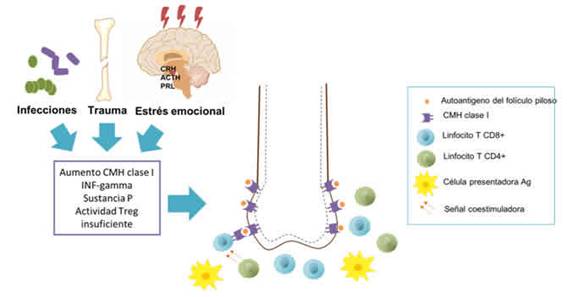
Figura 3: Factores desencadenantes de respuesta autoagresora y moléculas implicadas en la pérdida del privilegio inmune.
Se ha encontrado una preferencia en la aparición de la AA en folículos pilosos en anágeno con pigmentación completa en estadio IV lo que favorecería la evidencia de poliosis al examen clínico en estas áreas y relaciona la hipótesis de autoantígenos asociados a la melanogénesis presentados por CMH clase Ia como objetivos principales en el desarrollo de la enfermedad. 6,13
Sin embargo, es posible que se genere la restauración del PI en el siguiente ciclo si no se produce un nuevo ataque, por medio de la promoción intrafolicular de guardianes del PI endógenos por los queratinocitos induciendo la liberación de neuropéptidos inmunoinhibitorios a partir de las fibras nerviosas sensoriales. 6,21
Estrés emocional, colapso del PI y alopecia areata
La activación de neurohormonas por el estrés emocional ocurre a través del eje Hipotálamo-Pituitaria-Adrenal ocasionado por la regulación positiva de hormonas del estrés como la CRH (hormona liberadora de corticotropina), ACTH y los glucocorticoides. Estas hormonas de estrés se acompañan de una respuesta adicional mediada por neuropéptidos. 22,23
Ante situaciones de estrés psicológico se genera una respuesta central que conduce a la activación del eje hipotalámico-pituitario-adrenal, causando la liberación de la hormona liberadora de corticotropina (CRH), ACTH y prolactina (PRL). La liberación de CRH desencadena un aumento en la expresión del gen propiomelanocortina(POMC) en la porción anterior de la glándula pituitaria.POMC se convierte en hormona adrenocorticotropica(ACTH) que se une a los receptores de melanocortina tipo 2 (MC) de la corteza adrenal y estimula la síntesis y excreción a la circulación sanguínea de glucocorticoides para generar diversas funciones fisiológicas en el organismo. 21,22,24,25
El estrés emocional también induce liberación de catecolaminas por la vía medular-simpático-adrenal con liberación de epinefrina y norepinefrina que se unen a diferentes tipos de receptores adrenérgicos, produciendo una disminución en el flujo sanguíneo y alteraciones en el sistema inmunológico tales como activación de funciones inflamatorias, tránsito y proliferación de linfocitos y producción de citoquinas. 18,24
Un mediador clave en la respuesta al estrés por parte del folículo piloso es la sustancia P que estimula la degranulación de mastocitos desencadenando inflamación neurogénica, promoción de inmunidad celular, crecimiento de fibras nerviosas sustancia P +, activación de células endoteliales y receptores activados de proteína (PARs). (Ver figuras 4 y 5)La activación de mastocitos permite que se produzca supresión de la respuesta TH1 y cambio a TH2 promoviendo activación de IL-4.Por tanto en la alopecia areata se observa regulación positiva en folículos pilosos de sustancia P que pueden inhibir el crecimiento de pelo al interrumpir el mecanismo de privilegio inmune de la porción inferior en fase anágena. 22,24
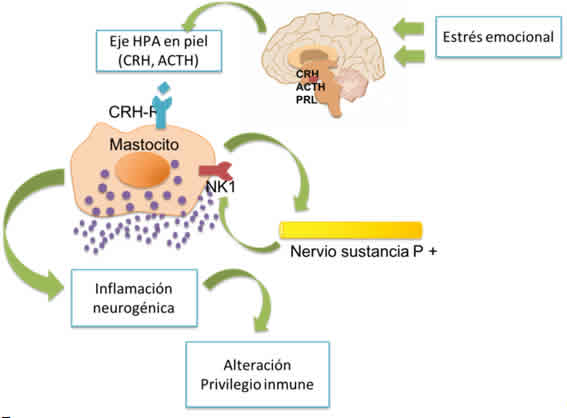
Figura 4: Estrés emocional como desencadenante de la activación del eje HPA e inducción de inflamación neurogénica potenciadora de la disrupción del privilegio inmune.
Figura 5: Asociación entre estrés emocional y colapso del inmunoprivilegio. Interacción entre linfocitos TCD8+ y mastocitos, aumento de INF-gamma y CMH clase I.
Conclusión
El folículo piloso es una estructura compleja ydinámica en la cual se llevan a cabo diferentes procesos fisiológicos, gracias al beneficio de ser una de las pocas estructuras anatómicas en la que se presenta un mecanismo protector de estas funciones como lo es el privilegio inmune. Este depende de diversos factores tales como la producción de inmunosupresores locales que inhiben y controlan la población celular de defensa en fase anágeno y las características propias de la matriz extracelular del folículo piloso que le brindan un ambiente aislado de posibles células autoagresoras. Al presentarse un desequilibro en alguno de los mecanismos protectores se produce una respuesta autoreactiva, característica de la alopecia areata que puede verse potenciada por el estrés emocional, activando el eje hipotálamo-hipófisis-adrenal con la inducción de inflamación neurogénica y aumento en la activación ydegranulación de mastocitos,que provoca un mayor reclutamiento de linfocitos T agresores del folículo piloso, reflejando la posible asociación entre el colapso del inmunoprivilegio y el estrés emocional en la alopecia areata, sugiriéndonos posibles manejos terapéuticos futuros enfocados en reducir esta respuesta potenciadora de autoagresión,mostrando la importancia del acompañamiento terapéutico profesional de estos pacientes a los distintos cambios emocionales causantes de estrés.

