











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. INTRODUCCIÓN
La provincia de Entre Ríos ha sufrido históricamente un intenso proceso de transformación de sus bosques nativos. Las elevadas tasas de deforestación obedecen principalmente al avance de la frontera agrícola y al aprovechamiento forestal selectivo de los bosques, desencadenando procesos destructivos, áreas abandonadas y desarrollo de bosques secundarios. Los sistemas de bosques constituyen el recurso básico sobre el que se sustenta la ganadería basada en el aprovechamiento de los pastizales naturales.
Actualmente quedan escasos relictos que conservan la vegetación autóctona, generalmente muy degradados y expuestos a grandes presiones, tales como el sobrepastoreo y las quemas descontroladas (Rainer Cinti, 1998). La deforestación constituye un disturbio intenso en áreas de bosques nativos, desencadenando procesos de sucesión secundaria en los que se van reemplazando las comunidades vegetales originales por otras de inferior valor, y con tendencia a restituir en la etapa final, la vegetación original. Estudios de zonificación de bosques nativos de Entre Ríos (Sabattini et al., 2009) han detectado modificación en la estructura original, debido al enmalezamiento, tala e invasión de especies exóticas.
La recuperación de la cobertura vegetal en cualquier terreno, a partir del proceso sucesional, depende de una serie de factores bióticos y abióticos, y de procesos ecológicos. Entre los factores bióticos, están la presencia del banco y la lluvia de semillas, como los principales dinamizadores naturales de la sucesión (Bedoya- Patiño et al., 2010). El banco de semillas del suelo (BSS), entendido como la reserva de semillas viables que potencialmente son capaces de germinar y establecerse (Cano-Salgado et al., 2012), constituye el medio más importante para la regeneración de la vegetación con especies pioneras, en terrenos donde el uso del suelo no ha sido muy intenso (Garwood, 1989; Dalling y Hubbell, 2002). Sin embargo, Tekle y Bekele (2000) sostienen que en áreas degradadas, la sola presencia del banco de semillas no es suficiente para la recuperación de la vegetación, en especial aquella de hábito arborescente.
Los BSS son muy dinámicos en una comunidad, y presentan alta heterogeneidad en composición y abundancia, tanto espacial como temporal (Paluch, 2011). Garwood (1989) destaca la importancia de la composición y dinámica del banco de semillas del suelo, para predecir el curso de la sucesión secundaria, cuando se presentan perturbaciones en los bosques nativos. El banco de semillas del suelo representa el potencial regenerativo de las comunidades vegetales y cumple un papel preponderante en la recuperación de áreas con drásticos procesos de disturbios (Henderson et al., 1988; De Souza et al., 2006). El BSS puede cumplir un papel fundamental en la recuperación de áreas disturbadas, requiriéndose la implementación de prácticas de manejo y conservación de estos BSS para el mantenimiento de la diversidad florística y la sustentabilidad social y económica de estos ecosistemas (De Souza et al., 2006).
En Argentina diversos trabajos abordan la problemática de la composición florística del BSS en suelos agrícolas bajo diferentes tipos de laboreo (Leguizamón y Roberts, 1982; Leguizamón, 1983; Díaz, et al. 1994, Pucheta et al., 1998), en relación con disturbios como el fuego o el pastoreo (Suarez et al., 2009; Ernst y Morici, 2013) y la similitud entre la vegetación emergente y el BSS (Haretche y Rodriguez, 2006; Etchepare y Bocanelli, 2007), pero son escasos los estudios en bosques nativos. Por lo expuesto, se considera prioritario estudiar el BSS en agroecosistemas boscosos disturbados , a efectos de inferir la posibilidad de restauración de las comunidades vegetales originales a partir del mismo.
El objetivo de este trabajo fue realizar un análisis comparativo del banco de semillas del suelo en dos etapas sucesionales del bosque nativo.
2. MATERIALES Y MÉTODOS
Caracterización del área de estudio
El estudio se realizó en un establecimiento agropecuario del Departamento La Paz, en la provincia de Entre Ríos. El área pertenece a la Provincia Fitogeográfica del Espinal, Dominio Chaqueño (Cabrera, 1976). La vegetación está caracterizada por bosques semixerófilos dominados por ñandubay (Prosopis affinis Sprengel.) y espinillo (Acacia caven Mol, Molina.). Son bosques generalmente abiertos, compuestos por un estrato arbóreo bajo, menor a 6 m, con algunos ejemplares aislados que excepcionalmente sobrepasan los 12 m de altura.
En el establecimiento agropecuario en estudio predominan los suelos Alfisoles (Ocracualfes vérticos), seguidos por Molisoles (Argiudoles vérticos y Argiudoles ácuicos). Las precipitaciones registradas en la localidad de San Gustavo (próxima al establecimiento) alcanzaron un total de 692 mm para el período Julio a Diciembre de 2012, excediendo en 190 mm a la media histórica para ese lapso de tiempo, traduciéndose en un año con excedente hídrico, coincidente con el período de floración de Prosopis spp. Para el período enero - abril del año 2013 se registró un total acumulado de 248,6 mm, resultando un déficit hídrico de 243 mm respecto a la media histórica (SIBER, 2014). Por su parte, las temperaturas máximas mensuales medias superaron durante casi todo el período los valores históricos en aproximadamente 2 ó 3 °C; mientras que las mínimas mensuales se mantuvieron dentro de los valores que indican los promedios históricos para esa zona (http://siga2.inta.gov.ar/en/ datoshistoricos/ y http://www.mineria.gob.ar/estudios/irn/entrerios), siendo julio el mes más frío (mínimas medias históricas de 5.5 ºC) y enero el más cálido (máximas medias históricas de 31 ºC).
Los ensayos fueron instalados en lotes que presentan el mismo tipo de suelo, Argiudoles vérticos (Molisoles), correspondiente a la Serie Banderas. Son suelos desarrollados sobre materiales limo-loessoides, poco profundos, con un epipedón de textura franca a franca limosa, seguido de un horizonte argílico fuertemente textural (Plan Mapa de Suelos, 1986).
Ensayo y diseño experimental
Se caracterizó el banco de semillas del suelo, evaluándose dos tratamientos: T1) suelo de bosque nativo considerado testigo (BN); y T2) suelo de bosque en regeneración (BREN), con tres repeticiones cada uno. Para la diferenciación de los tratamientos, se siguió el criterio de Sabattini et al. (2009) y Muracciole (2008), quienes establecen que el bosque nativo corresponde a una categoría de bosques que se encuentran en la etapa final de la sucesión vegetal (clímax) con modificaciones antrópicas generadas por las actividades productivas.
El bosque nativo evaluado (T1) presenta características de bosque bajo, inferior a 6 m de altura (Sabattini et al. , 2009), con dominancia de P. affinis (30 % de cobertura), acompañado de Celtis tala Gillies ex Planch. y A. caven. Se registró un enmalezamiento con especies arbustivas, dado principalmente por un 37,5 % de cobertura de Baccharis punctulata DC., acompañado por Aloyssia grattíssima y especies de malezas herbáceas de gran porte como Melica macra Nees. y Eryngium horridum Malme. El estrato herbáceo es continuo, constituido fundamentalmente por especies de Poáceas cespitosas que dan lugar a una comunidad herbácea de alto valor forrajero.
Por su parte, el T2 consiste en suelos con bosques nativos en regeneración desencadenada a partir de la deforestación. Esta área sufrió un desmonte en el año 2000, para su incorporación a la agricultura, básicamente cultivo de soja en rotación con trigo, actividad que se mantuvo durante un período de 6 años. La vegetación actual es un bosque bajo (inferior a 6 m de altura) dominado por A. caven, con un valor de cobertura de un 30 %. En este tratamiento se observó un alto grado de enmalezamiento (75 %) con especies invasoras arbustivas tales como Baccharis notosergila Griseb., Baccharis punctulata y Baccharis coridifolia DC. Entre las especies herbáceas dominantes se observó la presencia de Eryngium horridum y Polygonum aviculare como malezas herbáceas y especies de Poáceas tales como Stipa neesiana, Piptochaetium montevidensis y Lolium multiflorum.
Tanto el BN como el BREN se destinan desde el año 2008, a la actividad de cría y recría de ganado vacuno, con una carga promedio anual de 1,2 equivalente vaca/ha.
A fines de abril de 2013, se efectuó la extracción de muestras de suelo para la evaluación del BSS, siguiendo un método de muestreo sistemático (Cochran, 1984). En cada tratamiento, se delimitaron tres parcelas de 100 m x 100 m, en cada una de las cuales se dispuso una transecta diagonal, con 10 puntos de muestreo (unidades muestrales) de suelo a intervalos regulares, alcanzando un total de 30 puntos por tratamiento. Las muestras de suelo fueron extraídas con un barreno de 10 cm de diámetro, a dos profundidades: 00-05 cm y 05-10 cm. El procesamiento se realizó por el método de la separación física de las semillas (Leguizamón, 1983), a través del tamizado y lavado con agua, empleándose tres tamices de aberturas de malla Nº 10, 18 y 35. En la etapa de tamizado y lavado se aplicó el efecto del “estallido” (humectación rápida por inmersión en agua) para lograr la separación de los agregados de suelo por sus planos naturales de ruptura, minimizando posibles daños sobre las semillas presentes en las muestras (Wilson y Valenzuela, 1998). Por cada muestra procesada fueron obtenidas tres fracciones de semillas según tamaño, resultantes de su tamizado. Se separaron las semillas potencialmente viables. La viabilidad fue determinada ejerciendo una ligera presión con una pinza, considerándose que aquellas que resistían la presión eran viables (Tuesca et al., 2004).
El reconocimiento de las semillas se realizó bajo lupa binocular, utilizando claves de reconocimiento de semillas (Petetin y Molinari, 1982; Rodriguez, 1992).
Caracterización del Banco de semillas del suelo
La clasificación del BSS se efectuó de acuerdo a los criterios de Thompson (1992), quien propone: a) la presencia de un BST (banco de semillas transitorio) para aquellas especies cuyas semillas se presentan sólo en las capas superiores del suelo (00-05 cm de profundidad); b) BSPcp (banco de semillas persistente a corto plazo) para especies cuyas semillas son más frecuentes en capas superiores, pero también están presentes en capas inferiores (05-10 cm profundidad) y c) BSPlp (banco de semillas persistente a largo plazo) si las semillas son más frecuentes en las capas inferiores, o al menos tan frecuentes como en capas superiores.
Se determinó la composición específica del BSS, la densidad total y por especie (número de semillas por m2), y la frecuencia por especie. La frecuencia por especie se calculó como el porcentaje de unidades muestrales en las que la especie estuvo presente. Se estimó la diversidad específica del BSS en cada tratamiento, evaluado a partir del índice de Simpson (Braun Blanquet, 1979).
Análisis estadístico de datos
Los datos de densidad del BSS obtenidos fueron procesados estadísticamente aplicando, una vez comprobados los supuestos del modelo, comparaciones múltiples de medias (Test de Duncan al 5 %) para detectar diferencias entre los tratamientos evaluados. Se trabajó con el Software Estadístico InfoStat (2005). Los datos del BSS (densidad y composición) se analizaron a través de métodos multivariados (análisis de conglomerados). Las matrices de datos se clasificaron con el método de Ward y como medida de distancia se usó la Distancia Euclídea Promedio. Se obtuvieron grupos florísticos por tratamiento y por profundidad evaluada. Para estos análisis, fueron consideradas aquellas especies que se presentaron con valores de frecuencia superiores a 5 %.
3. RESULTADOS
Caracterización del banco de semillas del suelo
Densidad de semillas
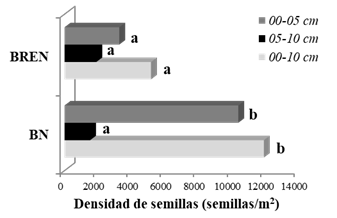
La densidad total de semillas del banco en los primeros 10 cm de profundidad resultó significativamente superior (p<=0,05) en el BN respecto al BREN, con un total de 11.917 semillas/m2 y 5.169 semillas/m2, respectivamente (Figura 1). La diferencia de densidad de semillas también resultó significativa entre ambos tratamientos al analizar la capa superficial del suelo (00-05 cm). En la profundidad de 05-10 cm los valores de densidad resultaron estadísticamente similares (Figura 1). En ambos tratamientos, las mayores densidades de semillas se registraron en los primeros 5 cm del suelo (Figura 1).
Composición florística del banco de semillas
T1. Bosque Nativo (BN)
Se registró un total de 57 especies en la capa 00-10 cm, pertenecientes a 19 familias, de las cuales las Poáceas y las Ciperáceas resultaron las más representadas, con 11 y 10 especies, respectivamente (Tabla 1). Del total de especies registradas, 5 no han podido ser identificadas a la fecha.
En la capa superficial del suelo (00-05 cm), las especies más frecuentes fueron Carex rostrata (86,7 %), Cyperus eragrostis (73,3 %), Cyperus polystachyos (66,7 %), Setaria geniculata (66,7 %) y Sida spinosa (63,3 %). Las especies que aportaron mayor densidad de semillas fueron Setaria geniculata (3032 semillas/m2), C. rostrata (920 semillas/m2), Brassica rapa (861 semillas/m2), Sida spinosa (768 semillas/m2) y P. montevidensis (550 semillas/m2), aportando densidades relativas de 29,2 %, 8,8 %, 8,2 %, 7,4 % y 5,3 %, respectivamente (Figura 2 a). La única especie arbórea registrada en el BSS fue A. caven, con una frecuencia promedio de 13,3 % y un aporte de 17 semillas/m2 al banco, habiéndose hallado sólo en los primeros 5 cm de suelo.
Tabla 1 Especies presentes en el banco de semillas del suelo (00-10 cm) en el bosque nativo (T1). Entre Ríos, ArgentinaTipo de banco: transitorio (BT), persistente a corto plazo (BPcp) y persistente a largo plazo (BPlp). Fr.: frecuencia
En la capa de suelo de 5 a 10 cm, para el mismo tratamiento (T1), las especies más frecuentes fueron Cyperus polystachyos (43,3 %), Setaria geniculata (33,3 %) y una especie incógnita (33,3 %). Los mayores valores de densidad correspondieron a Carex rostrata (259 semillas/m2), C. polystachyos (200 semillas/m2) y Setaria geniculata (147 semillas/m2), correspondiéndoles densidades relativas de 17 %, 13,7 % y 12 %, respectivamente (Figura 2b).
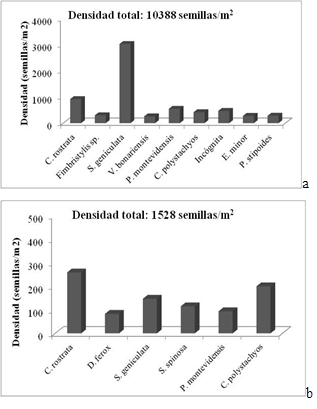
Figura 2 Densidad de semillas (semillas/m2) por especie y por profundidad para las especies de mayor densidad relativa.T1 (Bosque Nativo). a. 00-05 cm; b. 05-10 cm
En el BN, 20 especies forman un banco de tipo transitorio, 24 un banco persistente a corto plazo y 13 adoptan una estrategia de banco persistente a largo plazo (Tabla 1). La única especie arbórea registrada en el BSS (A. caven) forma un banco transitorio.
En este tratamiento, la clasificación de las muestras del BSS para los primeros 5 cm permitió distinguir tres grupos florísticos (análisis al 50 % de la distancia máxima) que se corresponden con la importancia relativa, dada por los valores de densidad relativa de las diferentes especies registradas (Figura 3a). El grupo I resultó integrado sólo por S. geniculata, siendo la especie de mayor contribución porcentual a la densidad total del banco (29,7 %). El grupo II, incluyó a 12 especies, que fueron aquellas que aportaron densidades relativas comprendidas entre 2 y 9 %, siendo C. rostrata, S. spinosa y P. montevidensis, las especies con valores mayores de densidad relativa (8,9 %, 7,4 % y 5,3 %, respectivamente). El tercer grupo quedó conformado por especies que contribuyeron con menos del 2 % a la densidad total del banco.
Respecto a la profundidad de 5 a 10 cm, se distinguieron sólo 2 grupos de especies (Figura 3b). El primero de ellos quedó conformado por 2 subgrupos, uno de los cuales está representado sólo por P. montevidensis, especie que se presentó con menor frecuencia y menor densidad relativa (5 %) respecto a las especies del otro subgrupo. Este segundo subgrupo incluyó a especies que tuvieron mayores densidades relativas en el BSS (C. rostrata, C. polystachyos, S. spinosa y S. geniculata). El grupo II quedó conformado por 15 especies que fueron aquellas con menor contribución porcentual al BSS.

T2. Bosque en regeneración (BREN)
Se registró una riqueza total de 45 especies, pertenecientes a 17 familias, de las cuales Poáceas resultó la más representada (8 especies), seguida por las Ciperáceas (5 especies), Fabáceas y Euforbiaceas con 4 especies cada una (Tabla 2). Del total de especies registradas, 5 no han podido ser identificadas a la fecha.
Tabla 2 Especies presentes en el banco de semillas del suelo (00-10 cm) en el bosque en regeneración (T2). Entre Ríos, Argentina.Tipo de banco: transitorio (BT), persistente a corto plazo (BPcp) y persistente a largo plazo (BPlp). Fr.: frecuencia
En la capa superficial del suelo (00-05 cm), las especies más frecuentes fueron S. sisymbriifolium (96 %), P. stipoides (76 %) y Verbena bonariensis (64 %). Las especies que aportaron mayor densidad de semillas fueron S. sisymbriifolium (986 semillas/m2), P. stipoides (709 semillas/m2), V. bonariensis (459 semillas/m2) y C. eragrostis (202 semillas/m2), contribuyendo a la densidad total BSS con 30 %, 21,6 %, 14 %, y 6,1 %, respectivamente (Figura 5a).
En la capa de suelo de 5 a 10 cm, las especies más frecuentes fueron S. sisymbriifolium (88 %), P. stipoides (72 %) y Verbena bonariensis (56 %). S. sisymbriifolium aportó el 48,4% de la densidad (919 semillas/m2). Le siguieron P. stipoides (236 semillas/m2) y V. bonariensis (216 semillas/m2), correspondiéndoles densidades relativas de 13,7 % y 12 %, respectivamente (Figura 5b). La única especie arbórea registrada en el BSS fue A. caven, con una frecuencia promedio de 6 % y un aporte de 27 semillas/m2 al banco (20 sem/m2 en la capa de 00-05 cm y 7 sem/m2 en la capa de 05-10 cm).
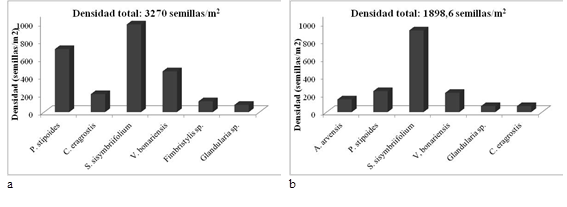
Figura 5 Densidad de semillas (semillas/m2) por especie y por profundidad para las especies de mayor densidad relativaT2 (Bosque en regeneración). a. 00-05 cm; b. 05-10 cm
En el BREN, 18 especies constituyen un banco transitorio, en tanto que 27 especies forman un banco persistente, 16 de corto plazo y 11 de largo plazo. En esta situación las semillas de la única especie arbórea determinada constituyen un banco persistente de corto plazo, habiéndose encontrado tanto de 00-05 cm como de 05-10 cm de profundidad, aunque con bajos valores de densidad media y frecuencia (Tabla 2)
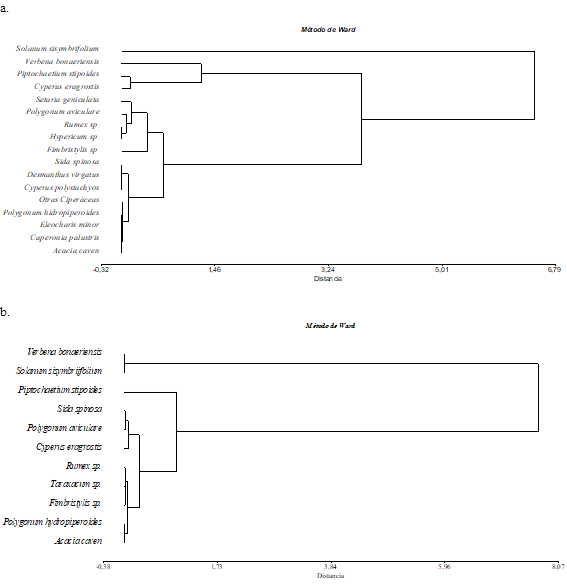
La clasificación de las muestras del BSS para los primeros 5 cm de BREN permitió distinguir tres grupos florísticos, que quedaron conformados de acuerdo a la importancia relativa, dada por los valores de densidad relativa de las diferentes especies registradas (Figura 6a). El primer grupo quedó integrado sólo por una especie (S. sisymbriifolium), que aportó el 30 % de la densidad total del BSS. El grupo II incluyó a un total de 3 especies (P. stipoides, V. bonariensis y una especie incógnita) que contribuyeron a la densidad total con el 21 %, 14 % y 6 % respectivamente. Por su parte, el resto de las especies quedaron in cluidas en el tercer grupo florístico, grupo que reunió a aquellas especies con valores de densidad relativa inferior a 4%. De estas especies, Fimbristylis sp. fue la especie de mayor contribución al BSS (3,7 %).
Del análisis a la profundidad 05-10 cm, la clasificación de las muestras arrojó un total de 2 grupos de especies (Figura 6b). El primer grupo estuvo formado por V. bonariensis y S. sisymbriifolium, que aportaron respectivamente el 48,3 % y 11,3 % de la densidad total del BS. El grupo II reunió al resto de las especies, que aportaron menores densidades relativas.
Diversidad específica
La diversidad específica determinada del BSS de 00-05 cm resultó estadísticamente similar (p<=0,05) entre los tratamientos evaluados, con valores que oscilaron entre 0.819 (BREN) y 0.824 (BN). En la capa de suelo de 05-10 cm de BREN, la diversidad específica fue de 0.705, estadísticamente inferior (p<=0,05) respecto a la determinada en BN (0.905).
4. DISCUSIÓN
Las densidades de semillas del BSS en el BN resultaron similares a las encontradas por Sabattini et al. (2011) en un bosque nativo del Espinal entrerriano. Estos autores determinaron en el BSS, densidades comprendidas entre 2.853 semillas/m2 y 11.076 semillas/m2. Por su parte, Ferri et al. (2009) en una investigación sobre evaluación del banco de semillas de un relicto del Espinal (Córdoba), determinaron densidades medias de 25.525 semillas/m2. Esta amplia variación en las densidades de semillas, producto de tan diversos factores, hace muy difícil las generalizaciones en relación a densidad de semillas del BSS.
La mayor parte de las especies identificadas en el banco de semillas en ambas comunidades evaluadas fueron formadoras de bancos persistentes, de las cuales las persistentes a corto plazo en el suelo (de uno a cinco años), resultaron las más abundantes. En el BREN se observó una menor participación de especies nativas al BSS, si bien la diferencia respecto al BN no resultó significativa. En los dos tratamientos, las familias botánicas más representadas en el BSS fueron las Poáceas y Ciperáceas. En BN, las semillas de especies forrajeras tales como Setaria geniculata y especies del género Piptochaetium contribuyeron con un 35 % de la densidad total de semillas del BSS, representando un potencial para la regeneración del pastizal natural.
El BREN, desencadenado a partir de la deforestación, presentó un BSS de menor riqueza específica, menor tamaño y menor proporción de especies nativas, respecto al bosque nativo estable. En este tratamiento se observó una participación porcentual de semillas de especies forrajeras en el BSS, un 42 % inferior respecto a BN. Resultados similares han sido reportados por otros investigadores. Gianaccini et al. (2009) determinaron una reducción de la riqueza específica y del tamaño del BSS en agrosistemas que reemplazan a la vegetación nativa característica de pastizales. Estos autores afirman que la sensibilidad mostrada por el banco de semillas frente al cambio en el uso de la tierra, lo convierte en un muy buen indicador del impacto ambiental provocado por la expansión de la actividad agrícola. De igual modo Latorre et al. (2001) consideran que la cantidad de propágulos del BSS podría constituir un indicador eficaz de la intensidad del disturbio experimentado en el área y de su capacidad de recuperación.
Respecto al aporte de semillas de especies malezas al BSS, en el BN se determinó aproximadamente un 25 % mientras que en el BREN este valor resultó próximo al 50 %, constituyendo un potencial de enmalezamiento del pastizal en detrimento de su calidad forrajera. Al respecto, Abella and Springer (2012) afirman que los incrementos en las abundancias de especies de malezas en el BSS indican disturbios antrópicos por deforestación, actividades agrícolas e incendios. Por su parte, López Toledo y Martínez Ramos (2011) alertan sobre los altos porcentajes de especies invasoras no nativas halladas en bancos de semillas de pastizales.
Acacia caven constituyó la única especie arbórea presente en el BSS de ambos tratamientos, presentándose con bajos valores de frecuencia y densidad relativa. En el BN, esta especie formó un banco transitorio, mientras que en el BREN se presentó formando un banco persistente a corto plazo. Diversos autores han reportado que las especies arbóreas están poco representadas en el BSS (Thompson, 1992; Carrillo-Anzures et al., 2009; Martinez Orea, 2013). Al respecto, en estudios realizados en sistemas de bosques nativos, Thompson (1992) observó que las semillas de los árboles presentes en la vegetación implantada suelen estar ausentes en el suelo, o si están presentes forman bancos transitorios o persistentes a corto plazo. En una investigación realizada en un relicto del Espinal (Córdoba, Argentina), sólo tres especies leñosas aparecieron en el banco del suelo, mientras que el resto de las semillas correspondió a especies herbáceas (Ferri et al., 2009). Sin embargo en el presente estudio, la ausencia de semillas de las especies arbóreas dominantes de BN (P. affinis) podría estar asociado a los problemas de fructificación que experimentó esta especie en el período en estudio, dado por los excedentes de precipitaciones registrados. La floración del algarrobo se ve especialmente afectada por las lluvias intensas y abundantes, que en esta zona ocurren en octubre-noviembre y producen el lavado de las flores. Esta podría ser una de las causas de la escasa producción de frutos de P. affinis en el sitio evaluado (Lallana et al., 1999). Otro posible factor que podría explicar la ausencia de semillas en el BSS de las especies arbóreas dominantes de los bosques sería la posibilidad de que las mismas formaran un BT de corta duración, con una inmediata germinación. A su vez, no se han observado plántulas de Prosopis spp. en el momento del muestreo, por lo que se podría inferir que una vez germinadas las semillas, la etapa de establecimiento de las plántulas constituiría una etapa crítica de la especie.
En el caso de bosques en pastoreo, el pisoteo del ganado podría constituir un factor clave, dificultando el establecimiento de las plántulas. Al respecto, Galera (1996) sostiene que P. nigra se regenera bien por vía de semillas y coloniza áreas degradadas. Este autor ha demostrado que en P. nigra, la capacidad de rebrote a partir del sistema radical y de tocones constituye otro medio que facilita la perpetuación de esta especie. Lallana et al. (1999) determinaron una buena capacidad de rebrote, y tasa de crecimiento en individuos talados de P. affinis. Luego de una abundante brotación inicial, los individuos no produjeron más brotes hasta el próximo período de crecimiento, comprendido entre la primavera y el verano de cada año, coincidiendo además con el período inmediatamente posterior a las copiosas lluvias de temporada. Por su parte, Rufini et al. (2006) evaluaron, en bosques nativos de Entre Ríos, el crecimiento de rebrotes de cepas de P. affinis, registrando mayores valores de incrementos corriente anual y medio anual de rebrotes, en clases diamétricas comprendidas entre 21 cm y 30 cm. Por lo expuesto sería factible aseverar que la capacidad de rebrote de P. affinis a partir del sistema radical y de tocones constituiría un medio de mayor eficacia respecto al BSS, para asegurar la regeneración natural de las especies arbóreas dominantes de los bosques evaluados.
5. CONCLUSIÓN
A partir de la información generada en el presente trabajo, es posible afirmar que el aporte de semillas de especies herbáceas forrajeras al BSS, aseguraría la persistencia y regeneración del pastizal natural, recurso básico sobre el que se sustenta la actividad ganadera de cría y recría.
Sin embargo, los resultados obtenidos indican una importancia relativamente baja del BSS en los procesos de regeneración y mantenimiento del estrato arbóreo de los bosques nativos, dada la baja densidad de semillas de las especies dominantes, aún en el banco del BN. Se destaca la necesidad de profundizar en investigaciones de esta índole, poniendo énfasis en estudios de viabilidad y poder germinativo de semillas de las especies arbóreas más conspicuas de los bosques nativos, como así también otras posibles formas de reproducción de estas especies.

