











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
A partir del análisis y comparación de la dieta de las poblaciones prehispánicas y de los individuos nativos reducidos en instalaciones coloniales tempranas, como los pueblos de indios y las encomiendas del Paraná inferior, resulta posible indagar sobre las diversas consecuencias que generó la colonización hispánica en esta área (González Lebrero, 2002; Tapia, 2002, 2014). En especial, el estudio de esta cuestión puede proporcionar información sobre el impacto que generó en el acceso de las poblaciones nativas a los recursos tradicionales y a los nuevos alimentos europeos introducidos, así como también responder a interrogantes tales como: ¿continuaron utilizando algunos recursos básicos de la subsistencia tradicional? ¿se produjeron cambios significativos en la alimentación de los nativos y por ende en sus prácticas sociales?
Al respecto, diferentes estudios históricos y etnohistóricos han mostrado cómo se habrían desestructurado los modos de vida de las poblaciones nativas a partir del establecimiento de los distintos asentamientos coloniales (Guerín, 2000; González Lebrero, 2002; Fradkin y Garavaglia, 2009; Tapia et al., 2015). Entre ellos, las reducciones fueron una de las instalaciones más estables y de mayor impacto dentro del sistema de trabajo de las encomiendas, una empresa que buscaba controlar los levantamientos indígenas, utilizar su fuerza de trabajo y transformar sus prácticas sociales, económicas, políticas y religiosas (Assadourian, 1983; Stern, 1986; Bethell, 1990). Los hombres, mujeres y niños indígenas debían permanecer en esas instalaciones, brindando su mano de obra a cambio del adoctrinamiento religioso, seguridad y una extensión de tierra para cultivar y sustentarse, lo cual implicaba modificar diametralmente sus modos de vida, entre ellos sus patrones alimenticios (Lightfoot, 2005; Tapia, 2002, 2014).
Dentro del Humedal del Paraná Inferior (HPI), Santiago del Baradero fue una de las diferentes reducciones que fundó Hernando Arias de Saavedra en 1615, pero de todas ellas fue la única que perduró en el tiempo, pasando a ser un pueblo de indios y luego a convertirse en pueblo de criollos (siendo actualmente la ciudad de Bardero). De estas primeras instalaciones se cuenta con las fuentes escritas del Archivo General de Indias (AGI) que brindan información de relevancia acerca de los modos de vida en la reducción, incluyendo la referencia sobre algunos recursos alimenticios que se consumían. Por otra parte, existen numerosos estudios paleodietarios, basados en el análisis de isótopos estables (AIE) que han indagado sobre los alimentos consumidos por los grupos nativos que habitaron en la región del Paraná inferior durante el periodo prehispánico (Loponte y Acosta, 2003-2005; Rizzo et al., 2016; Bonomo et al., 2017; Salazar-García et al., 2018; Loponte, 2020; Loponte y Mazza, 2021; entre otros). Por el contrario, son escasos los datos isotópicos que permitirían inferir la dieta de los individuos reducidos y encomendados, que habitaron en los distintos pueblos de indios del área y de otras regiones del país durante el periodo colonial temprano (Tafuri et al., 2017; Cardozo et al., 2018; Valenzuela et al., 2019).
En este trabajo nos proponemos contrastar la información referida a la dieta proveniente de las fuentes documentales mediante el análisis de la composición isotópica del carbono y nitrógeno (δ 13C, δ 15N) sobre restos esqueletales provenientes del sitio arqueológico Cementerio Indígena (Baradero, Provincia de Buenos Aires) (Figura 1). Adicionalmente, se presentan dos modelos inferenciales multivariados, el de grupos dietarios propuesto por Froehle et al. (2012) y FRUITS (Food Reconstruction Using Isotopic Transferred Signals) (Fernandes et al., 2014). En este último se integra a su vez, información para la reconstrucción de la ecología isotópica de la región y para determinar el aporte que podría haber tenido cada grupo alimenticio. Esta información permitirá realizar un análisis comparativo con las poblaciones prehispánicas que habitaron el área, analizar el impacto que produjo la conquista hispánica sobre las formas de subsistencia tradicional y evaluar las causas y consecuencias nutricionales de los cambios observados.
El análisis de la alimentación tradicional de los nativos a partir de las fuentes documentales
Algunos datos de interés para discutir los cambios en la alimentación de las poblaciones nativas, durante la conformación de las reducciones indígenas, pueden obtenerse de la documentación disponible en el Archivo General de Indias (AGI), que corresponde a la Audiencia de Charcas (ACH) y Audiencia de Buenos Aires (ABA). De allí se desprende que, durante la primera etapa de las instalaciones españolas en la región, la mano de obra para trabajar los cultivos que se pretendían instaurar era escasa “...no ay casi personal domestico y a esta causa está la gente muy pobre porque las haciendas de aquí son Sementeras de trigo y faltando quien las haga no tienen nada…” (AGI-ACH, 1599, f. 3). Al respecto, los funcionarios españoles esbozan algunas causas relacionadas principalmente con la economía de caza, recolección y pesca de estos grupos, por ejemplo, indican que estos individuos “no tienen casa ni asiento” (AGI-ACH, 1599, f. 1) o como destaca el gobernador Negrón, la mayoría de los nativos no necesitan sustentarse de los alimentos propuestos/impuestos por los españoles (AGI-ACH, 1610, f. 10).
Al respecto, los registros documentales y arqueológicos dan cuenta de la presencia de una macro-etnia en el Paraná medio e inferior (Cerutti, 2009) a la cual Politis y Bonomo (2018) llaman de manera genérica chaná-timbú y que habría estado compuesta por distintas parcialidades con distintos grados de afiliación (chaná, mbeguá, chaná-timbú, chaná-mbeguá, timbú, entre otras). Sus economías se habrían basado principalmente en la caza-recolección-pesca (CRP), aunque los registros etnohistóricos también dan cuenta del consumo de maíz por algunos de estos grupos, como los chanás (Fernández de Oviedo y Valdés, 1851-55; García de Moguer, extraído de Medina, 1908, pp. 244-245; Ramírez, 2007 [1528]). Por su parte los grupos guaraníes desarrollaron prácticas hortícolas que incluían el cultivo de maíz y la mandioca, entre otros vegetales, y complementaron su alimentación con recursos obtenidos mediante la caza y la pesca (Lothrop, 1932; Politis y Bonomo, 2012; Politis, 2014; Auge y Tapia, 2021; Cardozo et al., 2021). A pesar de la resistencia presentada, los guaraní y aquellos grupos que tenían algún grado de prácticas hortícolas fueron más susceptibles a ser reducidos debido que las mismas implicaban cierto grado de sedentarismo (González Lebrero, 2002).
No obstante, la forma de vida reduccional incidió en el estado nutricional de los nativos. Entre otras circunstancias se destacan: las limitaciones de sus prácticas habituales de sustento debido al trabajo en la reducción o en las tierras del encomendero, el tiempo que debían dedicar a las prácticas religiosas, la poca abundancia de ganado dentro de la reducción, y una economía previa que no se basaba en la acumulación intensiva de alimentos (Tapia, 2002). Si bien, la introducción del ganado vacuno en los campos pampeanos significó un recurso nuevo y de fácil obtención para los nativos, por momentos el acceso fue restringido y castigado con severidad. Por ejemplo, en el año 1616 ante la matanza desmedida de hembras y terneros se prohibió su consumo durante algunos años (AGI-ACH, 1616 f. 15; 1617, f. 19). Otros ejemplos de las prohibiciones fue el consumo de la yerba mate, pues consideraban que “es muy general este vicio, que además de ser sin provecho y que consumen y gastan sus haciendas en comprarla, hace a los hombres viciosos haraganes y abominables” (AGI-ACH, 1617, f. 18).
Como los documentos fueron escritos por distintos funcionarios del gobierno colonial, es posible encontrar incongruencias en los relatos. Por ejemplo, en el año 1619 el gobernador Diego de Góngora realizó una visita a la reducción Santiago del Baradero, y al respecto comentó que los nativos: “siembran maíz, frijoles y habillas y otras semillas con lo cual y carne de yeguas cimarronas y pescado que pescan en el río se sustentan” (AGI-ACH, 1619 f. 21). No obstante, el 7 de diciembre de 1619 el Fray Luis Bolaños elaboró un informe pidiendo apoyo económico para la orden franciscana de Santiago del Baradero, donde comenta que se sustentaban “con solo maíz cocido, y raíces” (AGI-ACH, 1619, f. 25). En una visita posterior, Góngora consideraba que el lugar donde había sido instalada la misión Santiago del Baradero no era el adecuado: “el sitio (es) pantanoso de mala agua y poca leña, y el pescadero esta desviado a tres leguas” (AGI-ACH, 1620, f. 28) razón que podría haber afectado la obtención de alimentos cuando escaseaba lo cosechado, especialmente el consumo de pescado. A pesar de las diferencias que pueden encontrarse en el relato de la documentación colonial, ésta fue narrada desde el punto de vista hegemónico de los funcionarios españoles, por ello las perspectivas arqueológicas y bioantropológicas resultan relevantes para generar un análisis crítico de las fuentes, establecer un mayor control de los sesgos etnocéntricos de los distintos autores y comprender los patrones de cambio económico y procesos de desestructuración étnica, de etnogénesis y de resistencia cultural que se llevaban adelante en estos establecimientos (Buscaglia, 2011; Oliva y Lisboa, 2009; Tapia, 2014).
El resultado de las reducciones en el Río de la Plata no fue el que esperaban los españoles. Los documentos refieren que muchos nativos no se adaptaron a las condiciones de vida planteadas por la dominación ejercida por españoles y criollos, tal es el caso de aquellos que huían espantados por las enfermedades, por los malos tratos y los trabajos forzados. Además, la resistencia cultural indígena y los desafíos a la colonización fueron favorecidos por la disponibilidad tanto de recursos autóctonos como introducidos (principalmente animales cimarrones), que en algunos casos les permitía sustentarse de manera autónoma. Aquellos pocos nativos que continuaron viviendo en el pueblo de indios quedaron sujetos a un tipo de dieta diferente a la tradicional.
Los estudios previos realizados de los materiales encontrados en el sitio Cementerio Indígena de Baradero (CIB), vinculado con el funcionamiento de la reducción Santiago del Baradero durante el siglo XVII, tanto arqueológicos como bioantropológicos (Tapia, 2002; Tapia y Pineau, 2011; Tapia, 2014; Kozameh et al., 2015; Cardozo et al., 2018; Auge et al., 2021; Cardozo et al., 2021; entre otros), han aportado abundante información acerca de la vida y el funcionamiento de esta misión, controlada por la orden franciscana, desde su fundación hasta su constitución administrativa como pueblo de indios (Nasif, 2021). Por ejemplo, se logró determinar que los individuos recuperados en el sitio arqueológico CIB presentan ancestría genética amerindia (Cardozo et al., 2021) y en una primera aproximación a la dieta de dos de ellos, mediante el análisis de isótopos estables, se observó que los peces y el maíz podrían haber contribuido a la alimentación de manera sustancial (Cardozo et al., 2018, Rizzo et al., 2018).
Paleodieta. Fundamentos teóricos y herramientas conceptuales
En la reconstrucción de la dieta, la composición isotópica del carbono brinda indicios acerca del patrón fotosintético dominante de las plantas y frutos consumidos directa o indirectamente a través de los animales que formaban parte de la alimentación. Por ello, es importante tener en cuenta el patrón fotosintético al que pueden pertenecer las plantas: 1) Calvin o C3, que representa a la mayoría de las especies de la flora mundial y poseen valores de δ 13C más bajos (por ejemplo, el trigo, el poroto, el arroz y la palmera pindó son algunas de las especies típicas de interés nutricional); 2) Hatch-Slack o C4, con el menor número de representantes y con valores de δ 13C más elevados (por ejemplo, el maíz, el sorgo y la caña de azúcar son exponentes de este patrón); y 3) Crasuláceas de metabolismo ácido o CAM que poseen valores de δ 13C intermedios (tunas y suculentas) (Tieszen, 1991). También se debe tener en cuenta que la composición isotópica de la fracción orgánica del hueso (colágeno) refleja, en mayor medida, la composición proteica de la dieta mientras que en la fracción inorgánica de huesos (bioapatita) los valores se correlacionan mejor con todos los componentes de la dieta que aportan al metabolismo energético (proteínas, carbohidratos y lípidos) (Ambrose, 1993; Frohele et al., 2012).
Por otra parte, a medida que los consumidores se encuentran más arriba de la cadena trófica, presentan una discriminación isotópica de 3-4‰ en los valores de 15N, en relación a sus dietas (Ambrose, 1991). Como los carbohidratos y grasas no poseen nitrógeno, estos valores se relacionan principalmente con la proteína dietaria más el efecto de enriquecimiento trófico (Ambrose et al., 1997). En tal sentido, nos da un indicio del consumo de carne del individuo analizado y por ello, los carnívoros son quienes presentan valores más altos. Aquí se debe tener presente que las cadenas tróficas de los cuerpos de agua son más extensas y que el nitrato tomado por los productores primarios se encuentra más enriquecido en 15N (Montoya, 2007). Por ello, los recursos de origen marino y ribereño suelen exhibir valores de δ 15N más altos que aquellos de origen terrestre.
Esto último es importante en lo que respecta al análisis isotópico de Santiago de Baradero ya que el sector correspondiente al HPI y el Delta del Paraná, que se extiende desde la ciudad de Diamante (Entre Ríos) hasta la desembocadura del Río de La Plata, constituye una “cuña” subtropical formada por bosques en galería y atravesada por diversos arroyos, riachuelos y ríos, que penetran en un medio templado característico del paisaje pampeano (Bó y Quintana, 2013). Tales características ambientales propiciaron, como ya fuera mencionado, que durante el periodo prehispánico se asentaran e interactuaran diferentes grupos humanos con variadas formas de vida. En lo que respecta a las investigaciones isotópicas sobre este tema, se han observado diferencias claras en el análisis isotópico de restos humanos recolectados en sitios arqueológicos datados aproximadamente entre 1.800 y 460 años AP. Aquellos individuos que poseían una economía basada en la CRP o con horticultura incipiente han presentado valores de δ 13C más bajos, indicando un mayor consumo de plantas con patrón fotosintético C3; mientras que para individuos provenientes de grupos horticultores, como los guaraníes, se observaron valores más elevados, lo cual indicaría que el maíz habría sido una parte importante de su economía (Loponte, 2020). Resulta fundamental aclarar que investigaciones isotópicas disponibles para esta región muestran que la fauna poseía una dieta basada principalmente en alimentos C3, a excepción del carpincho (Hydrochoerus hydrochaeris) que habría consumido mayor proporción de alimentos C4 (Loponte et al., 2011; Loponte et al., 2016b). Por otro lado, si bien los valores de δ 15N son más elevados en los grupos cazadores recolectores, los grupos horticultores también presentaron valores altos, lo cual señalaría que en su alimentación habrían aprovechado otros recursos regionales, tal como los peces u otros animales disponibles (Acosta y Loponte, 2002-2004, Loponte y Acosta, 2007, Loponte et al., 2011, Ottalagano y Loponte, 2016; Loponte et al., 2016a; Loponte et al., 2016b).
Materiales y Métodos
Selección de muestras a partir de restos óseos y dentales
Para el análisis de la composición isotópica de carbono (13C:12C, δ 13C) y nitrógeno (15N:14N, δ 15N) se seleccionaron elementos óseos correspondientes a nueve individuos adultos y molares de dos de ellos provenientes del sitio CIB (33º 48’ 04’’ LS, 59º 30’ 06’’ LO), vinculado a la misión franciscana Santiago del Baradero, provincia de Buenos Aires (Kozameh et al., 2015). Este sitio fue excavado por Salvador Debenedetti en 1910, oportunidad en la que realizó el hallazgo de catorce enterratorios asociados con materiales tanto de procedencia europea como indígena (Debenedetti, 1911). Los restos óseos recuperados en el sitio por otros investigadores integran una colección museográfica donde se han separado los huesos del cráneo y poscráneo, un procedimiento típico de la arqueología a comienzos del siglo XX (Kozameh et al., 2015). Por las características de los materiales asociados a las inhumaciones, Debenedetti consideró que los hallazgos correspondían a momentos coloniales tempranos. Esta presunción pudo ser corroborada con cuatro fechados que se dieron a conocer en anteriores publicaciones y que arrojan una temporalidad entre 450 ± 25 a 230 ± 5 AP (Tapia, 2014; Cardozo et al., 2016; Rizzo et al., 2015). Adicionalmente, fue datada la muestra CIB 21, arrojando una temporalidad de 360 ± 20 años AP (UGAMS-27011). Es importante destacar que para conducir este estudio se han respetado las normativas establecidas por el Código Deontológico establecido por la Asociación de Antropología Biológica Argentina (Aranda et al., 2014) y que no existen comunidades originarias que hayan manifestado vínculos con los individuos recuperados.
Muestras seleccionadas para Ecología isotópica (EI)
A partir de datos arqueológicos y etnohistóricos se seleccionaron un conjunto de alimentos que podrían haber contribuido a la dieta de los individuos que habitaron la misión. Al no haberse rescatado restos de fauna del sitio CIB, se procedió a analizar restos del mismo periodo, pero de otros emplazamientos coloniales de la región. De los pozos de descarte de residuos domésticos del sitio Bolívar N° 373, Ciudad Autónoma de Buenos Aires, Argentina (Zorzi y Tchilinguirián, 2013) se analizaron restos de sábalo (Prochilodus lineatus), bagre blanco (Pimelodus albicans), dorado (Salminus brasilensis), oveja (Ovis aries) y vaca (Bos taurus); y del sitio Siempre Verde, Benito Juárez, provincia de Buenos Aires (Lanza, 2006-2007) restos de caballo (Equus caballus). Ambos sitios han sido adjudicados temporalmente al siglo XVII. Adicionalmente, se analizó un resto óseo de vaca proveniente del sitio Casa Fernández Blanco, Ciudad Autónoma de Buenos Aires, adjudicado al siglo XIX (Ramos et al., 2014).
Debido a la falta de conservación de la flora, se seleccionaron vegetales contemporáneos teniendo en cuenta los recursos mencionados en las fuentes escritas (AGI-ACH, 1619 f. 21; AGI-ACH, 1619, f. 25). Estos corresponden a semillas de maíz (Zea mays) y poroto negro (Phasaeolus vulgaris) que provienen del partido de General Rodríguez y de trigo (Triticum spelta) del partido de Coronel Pringles, ambas localidades en la provincia de Buenos Aires. Estas semillas no poseen modificaciones genéticas y fueron cultivadas sin el uso de pesticidas o fertilizantes químicos, condición relevante para disminuir sesgos comparativos y obtener resultados isotópicos cercanos a los valores que podrían tener los alimentos consumidos durante la época en que funcionó la misión (Szpack, 2014). Los análisis se realizaron por duplicado utilizando 1,5-2 gr de muestras del año 2014 o 2018 en cada determinación.
Procedimientos aplicados para los análisis isotópicos
En el laboratorio de Antropología Biológica del Centro de Ciencias Naturales, Ambientales y Antropológicas (CCNAA), Universidad Maimónides, se pretrataron los elementos óseos correspondientes a la arqueología isotópica (salvo EI2, EI3 y EI4) y la muestra CIB3, para lograr la extracción del colágeno y bioapatita según el protocolo propuesto por Tykot (2004, 2006) y Garvie-Lok et al. (2004), respectivamente. En el primer caso, el procedimiento consta de dos fases, la desmineralización y la eliminación de partículas posdepositacionales. El primer paso requiere un ataque con ácido clorhídrico (HCl 2 %) por 72 horas, cambiando el reactivo cada día. En etapas anteriores y posteriores a este paso, el material se sumerge por 24 horas en hidróxido de sodio con el fin de eliminar ácidos húmicos. Luego, la muestra es enjuagada y secada en una estufa a < 60 °C. El pretratamiento de la fracción inorgánica consiste en sumergir a las muestras en hipoclorito de sodio al 2% durante 72 horas para la eliminación de la materia orgánica. Luego son enjuagadas con agua desionizada en una máquina centrífuga, para luego ser secadas en la estufa a la misma temperatura que el caso anterior. A este procedimiento le continúa la eliminación de carbonatos posdepositacionales, mediante el uso de un buffer de ácido acético 0.1M durante cuatro horas.
El resto de las muestras que conformaron la ecología isotópica y los elementos óseos CIB10, CIB11, CIB16 (H y M), CIB 20 (H y M) y CIB21 fueron pre-tratadas en el Center for Applied Isotope Studies (CAIS, Universidad de Georgia, EEUU). Este proceso consistió en la limpieza con cepillo y lavado en un baño ultrasónico, luego se secó y trituró suavemente para tratar los pequeños fragmentos del hueso con ácido acético 1N con el fin de eliminar carbonatos secundarios. Posteriormente la muestra se trató, en vacío, con ácido clorhídrico 1N para disolver el hueso mineral y liberar el dióxido de carbono de la bioapatita. Los residuos se enjuagaron con agua desionizada y calentados a 80°C durante 6 horas bajo condiciones ácidas (pH=3) para disolver el colágeno y aislar las sustancias húmicas en el precipitado.
El pretratamiento de las muestras CIB15, CIB17, CIB18 y CIB22 para obtener bioapatita se realizó en el Laboratorio de Isótopos Estables en Ciencias Ambientales (LIECA), San Rafael, Mendoza. Para ello se limpió la superficie del hueso con una herramienta Dremmel®, luego se pulverizaron 30-40 mg de la muestra que se colocaron en un tubo eppendorf de 1,5 ml junto con H2O2 3% y se mezcló por agitación. Posteriormente se llevó adelante el proceso de aclarado donde se separó la bioapatita mediante centrifugación a 5.000 rpm durante 5 minutos, se descartó el líquido y se llenó el tubo con agua destilada. Luego se agitó y centrifugó a 5.000 rpm durante 5 minutos. Nuevamente se descarta el líquido y se repite el proceso dos veces más. Para remover los carbonatos contaminantes, se llenó el tubo con ácido acético 1M, se mezcló por agitación, luego se abre la tapa y se deja reposar por 15 minutos. Posteriormente, se repite tres veces el proceso de aclarado. Finalmente, la muestra se seca en un horno de secado a menos de 70°C. El pretratamiento de estas muestras para obtener colágeno se llevó adelante en el CAIS según el protocolo detallado en el párrafo anterior.
El análisis de la composición isotópica para todas las muestras se realizó en el CAIS. El equipo utilizado para la medición isotópica de colágenos y vegetales fue un analizador elemental acoplado a un espectrómetro de masa de relaciones isotópicas Thermo Scientific Delta V. La composición isotópica del carbono fue calibrada en relación con el estándar VPDB. La composición isotópica del nitrógeno fue calibrada en relación al nitrógeno atmosférico. La incertidumbre de la medición se controló utilizando tres estándares internos: Spinach (δ 13C: −27,6 ‰, δ 15N: −0,3 ‰), Bovine (δ 13C: −17,6 ‰, δ 15N: +8,1‰) y Blue harbor tuna (δ 13C: −16,7 ‰, δ 15N: 14,8 ‰). Se informó que la incertidumbre analítica total era ± 0,1 y 0,2 ‰ para δ13C y δ15N, respectivamente. El equipo utilizado en la medición de bioapatita fue un Gas Bench acoplado a un espectrómetro de masa de relaciones isotópicas Thermo Scientific Delta V. El estándar internacional utilizado fue NBS 18 (δ13C: −5,01 ‰), mientras que los estándares internos del laboratorio fueron Fisher (δ 13C: −0,7 ‰), A1296 (δ 13C: 2,4 ‰) y TSF-1 (δ 13C: 1,95 ‰). La precision analítica es de ± 0,1 ‰.
Se realizaron un conjunto de controles para corroborar la calidad de los datos isotópicos obtenidos. El primero implicó la recuperación de colágeno mayor al 1% con respecto al peso inicial de la muestra; a éste sólo fue sometida la muestra CIB3 ya que se le extrajo el colágeno en el CCNAA. El colágeno del resto de las muestras fue extraído en aquellos laboratorios donde se le realizaron los análisis (CAIS o LIECA). Además, se analizaron los porcentajes de carbono (entre 22,6 y 47%), de nitrógeno (entre 5,5 y 17,3%) y las relaciones C:N (2,9-3,6) para la totalidad de las muestras consideradas (Ambrose, 1990; De Niro, 1985).
Metodología utilizada en el análisis de datos
Para el análisis entre las muestras provenientes de CIB y sus diferencias con las muestras de procedencia pre-hispánica se utilizó una análisis de varianza y Test de Tukey (valor p <0,01), mediante el programa R (https://www.R-project.org/). Luego los datos de isótopos estables obtenidos de los individuos del CIB fueron analizados por dos métodos distintos que nos brindan información complementaria. En primera instancia se realizó un análisis comparativo con las muestras de la bibliografía utilizando el modelo estadístico multivariado propuesto por Froehle et al. (2012). Esta primera aproximación es útil para diferenciar en distintas poblaciones la relevancia de los distintos conjuntos de recursos en la dieta mediante el uso simultáneo de tres sistemas isotópicos, δ 13C apatita, δ 13Ccolágeno y δ 15N. Para ello, el modelo se basa en dos funciones discriminantes generadas a partir de un gran número de datos arqueológicos publicados que permiten definir cinco grupos dietarios (Froehle et al., 2012). El uso de nuestros datos en estas funciones permite ubicar las muestras en la gráfica, comparándolas con los grupos propuestos por los autores e identificando el aporte de los distintos componentes a la dieta:
Carbono: F1 = (0,322 x δ 13 C apatita ) + (0,727 x δ 13 C colágeno ) + (0,219 x δ 15 N) + 9,354
Nitrógeno: F2 = (-0,393 x δ 13 C apatita ) + (0,133 x δ 13 C colágeno ) + (0,622 x δ 15 N) - 8,703
Para la reconstrucción cuantitativa paleodietaria optamos por el modelo de mezcla bayesiano FRUITS (Food Reconstruction Using Isotopic Transferred Signals), en su versión 3.1 (https://sourceforge.net/projects/fruits/). Este modelo ha mostrado ser eficiente a la hora de estimar la probabilidad de las contribuciones de diferentes fuentes, sobre todo cuando los diferentes macronutrientes de los alimentos contribuyen en proporción variable a la señal isotópica de los tejidos del consumidor (Fernandes, 2016). Asimismo, permite manejar diferentes incertidumbres, tales como aquellas asociadas a las señales isotópicas de los grupos de alimentos potenciales, los fraccionamientos isotópicos entre la dieta y los tejidos y la contribución relativa de los macronutrientes (Fernandes et al., 2014; Fernandes, 2016). Además, este modelo permite introducir información preexistente, como es el caso de aquella procedente de estudios fisiológicos o metabólicos (Fernandes et al., 2014).
Para la reconstrucción de la ecología isotópica y la conformación de grupos de recursos alimenticios se utilizó información de vegetales y fauna de consumo humano generada en el marco de esta investigación (Tabla 1) y aquella disponible para el área del Paraná medio e inferior (Tabla Suplementaria 2). Al observarse un solapamiento entre los recursos de origen animal, terrestre y ribereños, para el modelo se decidió generar un macrogrupo integrando la variación de animales en general. A su vez, se contempló el grupo de plantas C3 y el maíz, como único representante de plantas C4. Es importante destacar que este modelo también fue aplicado para estimar la dieta de los grupos prehispánicos, por lo tanto, en ese caso, no se incluyeron los recursos introducidos por Europa en pos de una representación realista de la dieta anterior a la conquista. Los valores isotópicos procedentes de especímenes actuales fueron corregidos considerando el efecto Suess (Schwarcz y Schoeninger, 2012). La información relativa a la composición a nivel macronutricional de los alimentos, factores de discriminación e información previa se consideró de acuerdo con Killian Galván (2018) y Killian Galván et al. (2021).
Tabla 1: Valores isotópicos de plantas de interés nutricional. Procesadas y analizadas en el Center for Applied Isotope Studies (CAIS) de la Universidad de Georgia, EEUU. *1 procesadas y analizadas en el CAIS de la Universidad de Georgia, EEUU
| n | Especie | Muestra | Procedencia de las Muestras | Código de Laboratorio | δ 13C (V-PDV, ‰) | δ 15N (AIR, ‰) | C/N |
|---|---|---|---|---|---|---|---|
| 1 | Zea mays | EI11*1 | General Rodríguez, Buenos Aires | UGAMS-#24559 | -12,05 | 4,89 | 33,8 |
| 2 | EI12*1 | UGAMS-#39460 | -11,59 | 2,89 | 36,2 | ||
| 3 | Phasaeolus vulgaris | EI13*1 | UGAMS-#24560 | -28,64 | -0,34 | 17,3 | |
| 4 | EI14*1 | UGAMS-#39461 | -22,88 | 2,94 | 12,4 | ||
| 5 | Triticum spelta | EI15*1 | Coronel Pringles, Buenos Aires | UGAMS-#39462 | -25,52 | 3,6 | 26,1 |
| 6 | EI16*1 | General Rodríguez, Buenos Aires | UGAMS-#24561 | -26,69 | 9,35 | 22,3 |
También se utilizaron estos modelos para evaluar la información isotópica disponible para series esqueletales humanas prehispánicas del Paraná medio e inferior. De los trabajos publicados por otros investigadores se seleccionaron las muestras que disponían de datos de δ 13Ccol, δ 13Cap y δ 15N (Bonomo et al., 2017; Gascue et al., 2016; Loponte y Acosta, 2007; Loponte y Kozameh, 2009; Loponte et al., 2016a; Loponte et al., 2016b; Ottalagano y Loponte, 2016; Rizzo et al., 2016). En la Tabla Suplementaria 1 se encuentra una descripción detallada de las muestras utilizadas.
Resultados y Discusión
En la Tabla 1 se detallan los resultados obtenidos sobre semillas. Con respecto a los valores de carbono en vegetales debemos mencionar que estos son los esperables para las plantas medidas, siendo importante la amplitud presente al interior de una misma especie para el caso de P. vulgaris. Respecto a los valores isotópicos del nitrógeno, esta especie también muestra variación, pero es mayor la hallada en Triticum spelta. Debemos tener en cuenta que los valores son bajos si consideramos los utilizados por otras aproximaciones paleodietarias (e.g. Bonomo et al., 2017) donde se recurrió a ejemplares procedentes de zonas más áridas, como es el caso del área del centro oeste de la Argentina (Gil et al., 2010).
Los valores de carbono en los ejemplares de fauna (Tabla 2, Figura 2) muestran que en ninguno de ellos la dieta se encuentra dominada enteramente por pasturas C3 y que existe un aporte de plantas C4 a la dieta marginal en la mayoría de los casos.
Tabla 2: Valores isotópicos de animales analizados. *1 procesadas y analizadas en el CAIS de la Universidad de Georgia, EEUU; *2 procesadas en el laboratorio de Antropología Biológica del CCNAA (Universidad Maimónides, Buenos Aires) y analizadas en CAIS.
| n | Sitio | Cronología | Especie | Nombre común | Muestra | Código de Laboratorio | %C | %N | C/N | δ 13C (V-PDB, ‰) | δ 15N (AIR, ‰) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Bolivar 373 | Siglo XVII | Prochilodus lineatus | Sábalo | EI1*2 | UGAMS-#24551 | 41 | 14 | 3,3 | -21,9 | 5,1 |
| 2 | EI2*1 | UGAMS-#27013 | 39 | 14 | 3,2 | -25 | 5,9 | ||||
| 3 | Pimelodus albicans | Bagre blanco | EI3*1 | UGAMS-#27012 | 30 | 9,7 | 3,6 | -18,5 | 9,5 | ||
| 4 | Salminus brasilensis | Dorado | EI4*1 | UGAMS-#27014 | 32 | 10 | 3,7 | -22 | 11,1 | ||
| 5 | Ovis aries | Oveja | EI5*2 | UGAMS-#24552 | 42 | 15 | 3,3 | -18,9 | 7 | ||
| 6 | EI6*2 | UGAMS-#24553 | 42 | 15 | 3,4 | -17,5 | 6,9 | ||||
| 7 | Bos taurus | Vaca | EI7*2 | UGAMS-#24554 | 42 | 15 | 3,3 | -10,9 | 5,8 | ||
| 8 | Siempre Verde | Equus caballus | Caballo | EI8*2 | UGAMS-#24555 | 44 | 15 | 3,5 | -20,1 | 6,4 | |
| 9 | Siempre Verde | EI9*2 | UGAMS-#24556 | 45 | 16 | 3,3 | -21,5 | 7,5 | |||
| 10 | Casa Fernández Blanco | Siglo XIX | Bos taurus | Vaca | EI10*2 | UGAMS-#24557 | 45 | 16 | 3,3 | -18,3 | 7,5 |
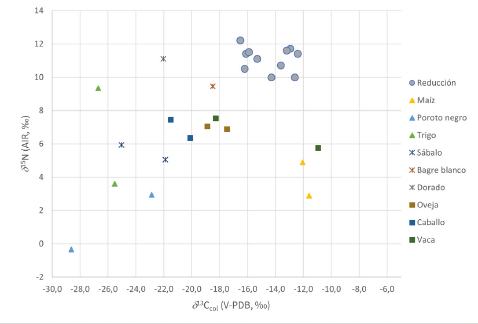
Figure 2: Carbon and nitrogen isotopic values measured on skeletal remains found in CIB site and food resource specimens from the province of Buenos Aires obtained in the framework of this research.
En lo que respecta a los elementos de Cementerio Indígena, de la muestra CIB3 se logró recuperar casi el 9,9% del colágeno. Con respecto al resto de los parámetros utilizados para considerar los valores como primarios, tanto los porcentajes de carbono como de nitrógeno obtenidos son aceptables, como también las relaciones C:N para la totalidad de las muestras consideradas (Tabla 3). Luego, una primera aproximación muestra que los valores medidos en colágeno presentan mayor variación en el caso del carbono respecto al nitrógeno, presentándose individuos que se enmarcan mayormente dentro del consumo de recursos C4 y otros C3. En cambio, todos parecen compartir un mismo nivel trófico, en tanto la diferencia en los valores de nitrógeno no llegan a ser mayores al 2‰. La variación es aún mayor cuando observamos los valores de carbono sobre bioapatita, donde el aporte del componente energético C4 parece ser relevante en algunos individuos y marginal en otros (Tabla 3).
Tabla 3: Restos humanos del sitio Cementerio Indígena de Baradero analizados. *1 procesadas y analizadas en el CAIS de la Universidad de Georgia, EEUU; *2 procesadas en el laboratorio de Antropología Biológica del CCNAA (Universidad Maimónides, Buenos Aires) y analizadas en CAIS y *3 procesadas en el Laboratorio de Isótopos Estables en Ciencias Ambientales (LIECA), San Rafael, Mendoza y analizadas en CAIS.
| n | Rótulo | Elemento esqueletal | Edad del tejido (años) | Código de laboratorio | δ 13Ccol (V-PDB, ‰) | δ 15N (AIR, ‰) | Código de laboratorio | δ 13Cap (V-PDB, ‰) | %C | %N | C:N | Funciones discriminantes Froehle et al. (2012) | Referencia | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F1 (C) | F2 (N) | ||||||||||||||
| 1 | CIB3*2 | Tibia der. | A | UGAMS-#24558 | -12,6 | 10 | UGAMS-#24562 | -7,5 | 45,1 | 16 | 3,2 | 0 | -1,2 | Este trabajo | |
| 2 | CIB10*1 | Radio izq. | A | UGAMS-#27009 | -13 | 11,7 | - | -9,3 | 41,9 | 15,3 | 3,2 | -0,5 | 0,5 | ||
| 3 | CIB11*1 | Radio izq. | A | UGAMS-#27010 | -14 | 10 | - | -9,7 | 42,6 | 15,5 | 3,2 | -1,9 | -0,5 | ||
| 4 | CIB15*3 | 2M sup. der. | 4-8 | MSR-805 | -12,4 | 11,4 | MSR-521 | -7,6 | 40,4 | 14,6 | 3,2 | 0,4 | -0,3 | ||
| 5 | CIB17*3 | 2M sup. der. | 4-8 | MSR-803 | -16,2 | 10,5 | MSR-525 | -11,2 | 38,7 | 13,9 | 3,2 | -3,7 | 0,1 | ||
| 6 | CIB21*1 | 2M | 4-8 | UGAMS-#27011 | -13 | 11,6 | - | -9,5 | 42,2 | 15,2 | 3,2 | -0,8 | 0,5 | ||
| 7 | CIB22*3 | Mandíbula | A | MSR-804 | -16,1 | 11,4 | MSR-524 | -9,7 | 40,4 | 14,6 | 3,4 | -3 | 0,1 | ||
| 8 | CIB16H*1 | Mandíbula | A | UGAMS-#17419 | -13,6 | 10,7 | - | -8,4 | 46,5 | 15,9 | 3,3 | -0,9 | -0,6 | Cardozo et al (2016) | |
| 9 | CIB16M*1 | 2M | 4-8 | UGAMS-#17420 | -15,3 | 11,1 | - | -9,6 | 46,1 | 16,1 | 3,4 | -2,4 | -0,1 | ||
| 10 | CIB20H*1 | Mandíbula | A | UGAMS-#17421 | -15,9 | 11,5 | - | -9,5 | 45,9 | 15,9 | 3,4 | -2,7 | 0,1 | ||
| 11 | CIB20M*1 | 3M | 10-15 | UGAMS-#17422 | -16,5 | 12,2 | - | -11,8 | 47,3 | 16,2 | 3,4 | -3,8 | 1,3 | ||
| Media | -13,2 | 10 | -9,4 | 43 | 15 | 3,3 | |||||||||
| d.s. | 0,7 | 0,1 | 1,3 | 2,9 | 0,7 | 0,1 | |||||||||
Cuando comparamos los valores isotópicos de los individuos de CIB respecto a aquellos asociados a contextos prehispánicos (Tabla Suplementaria 1), encontramos coincidencias con los de economías hortícolas, al menos en los valores obtenidos sobre el colágeno (test ANOVA y test de Tukey p < 0,01). Esta asociación es menos clara cuando observamos los valores de carbono en bioapatita (Figura 3b), no obstante, no se encuentran diferencias significativas entre estos dos grupos.
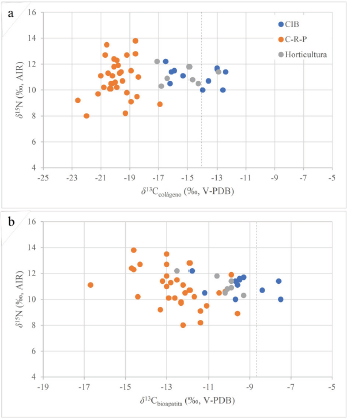
Figure 3: Isotopic composition of the organic (top) and inorganic (bottom) fractions of bone and dental remains from the CIB site and other pre-Hispanic sites of the middle and lower Paraná, discriminated by their predominant economy (hunting-gathering-fishing vs. horticulture).
Interpretación de valores según modelo multivariado
El modelo (Figura 4) muestra a los individuos adultos de la misión con una dispersión importante en relación a la Función 1 (carbono). En uno de estos extremos se encuentran CIB3, CIB10 y CIB15 con un aporte importante de alimentos C4, particularmente a la dieta total pero también en la fracción proteica, lo cual indicaría la ingesta de animales que consumían este tipo de alimentos. En el otro extremo se encuentran CIB17 y CIB22 cuya dieta puede explicarse por un mayor aporte de alimentos C3, tanto a la dieta total como a nivel proteico. Estos casos no solo abarcan los extremos de las dietas de los individuos de la misión, sino que también engloba a los individuos prehispánicos caracterizados por practicar la horticultura.
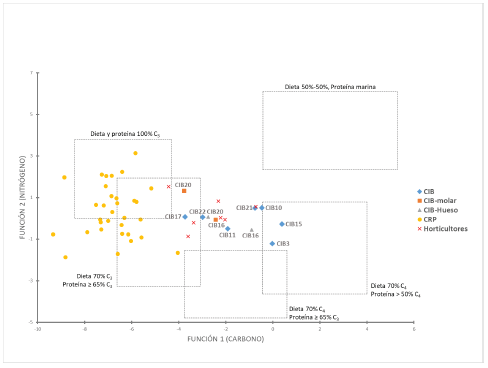
Figure 4: Pre- and post-Hispanic samples (according to the values obtained from the discriminant functions) plotted in comparison with the dietary groups proposed byFroehle et al. (2012).
Si bien, no se observa una gran dispersión en el eje de la Función 2 (nitrógeno) al analizar los dos casos en que se poseen datos isotópicos a partir de un molar y el resto óseo del mismo individuo (CIB16 y CIB20) se destaca que durante los primeros años de vida el componente proteico C3 y el consumo de proteínas fue más relevante que durante los últimos años de vida. Si bien se analizaron un segundo y un tercer molar, por lo que el efecto de amamantamiento no estaría influenciando estos resultados, sí debe tenerse en cuenta que al no poder determinar la edad de los adultos, es posible que los datos obtenidos entre el tercer molar y el resto óseo de CIB20 estén representando un periodo similar. No obstante, existen diferencias claras entre estos resultados, particularmente en los valores de δ 13Cap.
Los individuos caracterizados por poseer una economía de CRP se ubican dentro de los grupos dietarios de mayor consumo de alimentos C3, en concordancia con otros trabajos publicados para la región (Ottalagano y Loponte, 2016; Bonomo et al., 2017; Loponte, 2020). Estos grupos no habrían explotado ciertos alimentos como el carpincho, una especie que posee los valores de carbono menos negativos pero que se encuentra poco representada en los sitios de grupos cazadores-recolectores de la región y su caza podría haber tenido un significado simbólico (Sartori et al., 2014; Otalagano y Loponte, 2016). A diferencia de los otros grupos, aquí se observa una dispersión importante en relación a la Función 2 (nitrógeno), con varios individuos ubicados en el grupo de mayor consumo de C3, tanto para dieta como para proteína. Considerando que los peces suelen tener valores de nitrógeno más elevados, esto podría ser un indicador de la importancia de la pesca en la alimentación de muchos de estos individuos. No obstante, son necesarias mayores investigaciones sobre peces ribereños puesto que los valores disponibles hasta el momento no muestran una diferencia isotópica respecto a la fauna terrestre.
Interpretación de los valores a partir del modelo FRUITS.
Para la reconstrucción de la ecología isotópica y la conformación de grupos de recursos alimenticios, se utilizó la información generada en el marco de esta investigación, correspondiente a los ejemplares de flora y fauna; y a su vez, los valores disponibles en la literatura de fauna (Tabla Suplementaria 2), tales como son el ciervo de los pantanos (Blastocerus dichotomus), el venado de las pampas (Ozotoceros bezoarticus), el carpincho (Hydrochoerus hydrochaeris), el coipo (Myocastor coypus), el surubí (Pimelodidae) y el bagre sapo (Rhamdia) correspondiente al Paraná inferior y asociados a los sitios arqueológicos del Holoceno Tardío (Sartori et al., 2014; Loponte et al., 2016; Bonomo et al., 2017). Considerando la distribución en la composición isotópica de los diferentes recursos, entendemos que, para garantizar los requisitos de un modelo de mezcla bayesiano, resulta deseable generar grupos de alimentos con escaso o nulo solapamiento entre sí y que sean coherentes con nuestras preguntas de investigación. En este caso, buscamos saber si los individuos que habitaron la reducción poseían hábitos alimenticios radicalmente diferentes a aquellos que caracterizaron a las poblaciones prehispánicas de la región. En particular, si el maíz, generalmente asociado a aquellas dietas pobres en proteínas de buena calidad, se encuentra mayormente representado en la alimentación post hispánica.
Considerando los resultados del modelo de mezcla bayesiano FRUITS, hallamos una importancia relativa del maíz en al menos tres individuos, con un 35% de aporte de este cereal a la dieta. Incluso llega a ser de casi el 50% en el individuo CIB3 (Figura 5 Modelo 1, Tabla Suplementaria 3), coincidiendo con los resultados arrojados por el modelo multivariado (Froehle et al., 2012). Por su parte, el aporte del componente animal es menor al 35% en todos los casos, con la excepción del individuo CIB10 y CIB21. El aporte de las plantas C3 es mayor al 30% en todos los casos, siendo la excepción el individuo CIB15, donde este llega al 26%. Resulta llamativo que en el caso del estudio longitudinal de los individuos CIB16 y CIB20 el aporte del componente animal en la dieta cae en los últimos años de vida, mientras que el maíz cobra mayor relevancia, incluso hasta de manera dramática en CIB16.
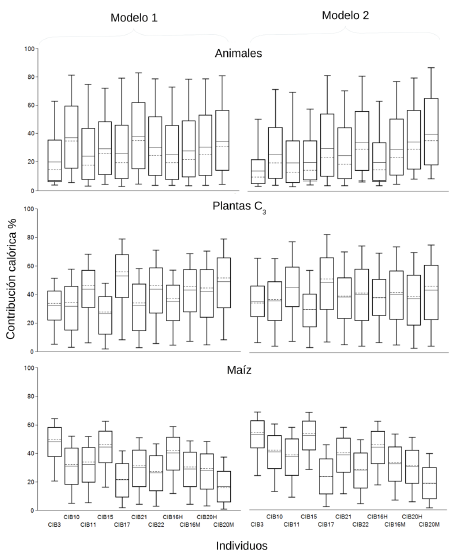
Figure 5: Box-plots corresponding to the caloric contribution (expressed in percentages) of the three groups of resources considered for the individuals of the CIB, including animals and vegetables introduced during the colony (Model 1) and excluding them (Model 2).
Este escenario cambiaría si sólo fueran considerados los recursos locales, no europeos en el modelo de estimación (Tabla Suplementaria 3). En el modelo 2 de la Figura 5, el maíz cobra mayor protagonismo en los individuos analizados, al no considerarse los especímenes europeos que mostraron tener una composición de la dieta mixta en cuanto la predominancia de plantas con patrón fotosintético C3 o C4. Por su parte, bajo esta premisa también se incrementa la relevancia de las plantas C3, al explicar siempre más del 35% de la dieta estimada de los individuos de la Reducción. A su vez, el componente animal explica siempre menos del 33% en la mayoría de los casos, siendo la excepción la dieta estimada durante la adolescencia del individuo CIB20M, quien presenta un aporte del 38% para ese conjunto de recursos.
En cuanto a la aplicación del modelo en los casos de individuos asociados a contextos prehispánicos (Figura 6), hallamos que la mitad de ellos poseen dietas que incluyen al menos un 30% o más del componente animal (Tabla Suplementaria 4). Esto incluso en uno de los individuos que poseía una economía hortícola. A su vez se observa una predominancia de plantas C3 o C4 según se trate de individuos con economías de CRP o de horticultura respectivamente, aunque es importante destacar que ninguna de las dietas se centró en un solo tipo de recurso vegetal. Análisis bioarqueológicos previos indican la presencia de fitolitos de maíz en el individuo proveniente del sitio Rancho José, el cual tiene una datación de 1900 años AP, por lo cual su consumo se habría encontrado en esta área mucho antes de la llegada más tardía de los grupos guaraníes (Auge y Tapia, 2023).
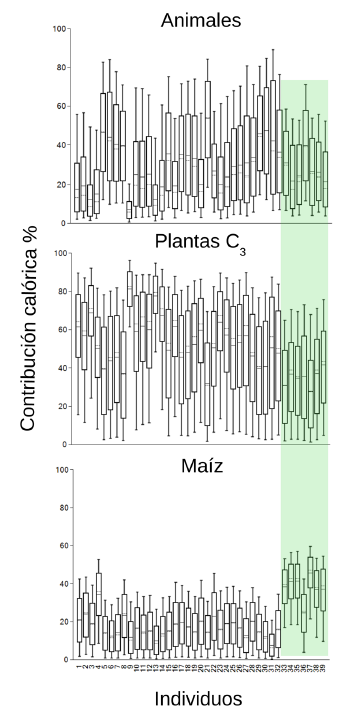
Figure 6: Box-plots corresponding to the caloric contribution (expressed in percentages) of the three resource groups considered for individuals associated with prehispanic occupations of the middle and lower Paraná (the numerical labels match the individuals in SupplementaryTable 1) including local animals and vegetables. Individuals associated with horticultural economies are indicated in the green box.
En conjunto, los resultados obtenidos de las distintas aproximaciones analíticas son coherentes entre sí. Las dietas de los individuos CIB presentan valores similares a los grupos horticultores prehispánicos de la zona, caracterizándose por un mayor consumo de alimentos asociados al patrón fotosintético C4, principalmente a nivel proteico, es decir, por consumo directo de vegetales como el maíz, pero también indirecto mediante animales que consumían plantas bajo ese patrón fotosintético, como el carpincho. No obstante, la alimentación de los individuos CIB no se habría basado únicamente en este tipo de alimentos ya que las plantas C3 explican solo un tercio de la dieta y en la mayoría de los casos el componente animal es menor al 35%. Respecto a este último punto es importante destacar que en aquellos individuos de los que se disponen datos para distintos momentos, en los últimos años de sus vidas, se observa un menor consumo de proteínas animales y un mayor consumo de alimentos C4. Finalmente, cuando los alimentos introducidos por los europeos no se consideran en la dieta de individuos del CIB, los aportes de plantas C3 y C4 incrementan mientras que disminuyen los del componente animal, en comparación con los individuos prehispánicos.
Discusión general
La información generada para la reconstrucción paleodietaria mediante el AIE de las poblaciones prehispánicas de la cuenca media e inferior del río Paraná da cuenta de la diversidad de patrones alimenticios, existiendo una predominancia variada de cada conjunto de recursos. La variación de los valores isotópicos en sí no solo es el resultado de los sistemas económicos predominantes, es decir, la relevancia de la caza-pesca y recolección o de la horticultura entre los diferentes grupos, sino también por la diversidad de recursos animales y vegetales que caracterizan a este sector de la región Mesopotámica.
El establecimiento de las reducciones en ésta área fue uno de los principales mecanismos de ruptura de los modos de vida de las poblaciones nativas. A pesar de la dificultad para la sedentarización, adoctrinamiento y explotación de la fuerza de trabajo que impusieron a las poblaciones nativas, se reestructuró la relación de estos grupos respecto a los aspectos más primarios y esenciales de la subsistencia. Los hábitos alimenticios no quedaron ajenos a ello, en tanto se modificaron las formas de ocupar el espacio y las modalidades de explotación de los recursos. En esta investigación, la información isotópica y las estrategias analíticas propuestas permitieron poner a prueba las hipótesis derivadas de otras fuentes de información.
El análisis bayesiano nos permitió indagar en la contribución de los distintos recursos a la dieta de los individuos prehispánicos. Al respecto, resulta interesante que en los individuos CRP se observó que el maíz contribuía a sus dietas en promedio el 21%, por lo que el consumo de esta planta se habría producido incluso mucho antes que la llegada de los grupos guaraníes al área. Este dato concuerda con los análisis de tártaro dental realizado sobre el individuo de Rancho José (Auge y Tapia, 2023). En tal sentido los circuitos amplios de interacción e intercambio con grupos nativos de otras regiones propuestos por Bonomo et al. (2009) y Bonomo et al. (2011) para materias primas líticas, también podrían haber sido utilizados para los alimentos desde momentos tempranos. Respecto a la relevancia del maíz, en las dietas de los grupos guaraníes se observa que su consumo habría rondado el 40% de los alimentos analizados, lo cual indica que, si bien fue un componente importante, la dieta no se encontraba dominada por este recurso. A su vez, si nos detenemos a analizar el aporte proteico de la dieta, el componente animal pareciera estar definiendo las diferencias isotópicas observadas entre las economías extractivas y productivas, por ejemplo, el consumo de animales con preferencia por plantas C4 y también, de alto retorno. Estas dos variables coinciden en los carpinchos, si nos remitimos a los valores obtenidos en otras investigaciones que indican una media de δ 13Ccol -15,2 ±2,5‰ (Loponte et al., 2016b; Ottalagano, 2021). Debemos destacar que esto concuerda con las expectativas generadas a partir de estudios de arqueofauna realizados a lo largo del Paraná, donde se señala que este recurso no habría sido explotado asiduamente por las poblaciones cazadoras-recolectoras-pescadoras (Sartori et al., 2014; Ottalagano y Loponte 2016). A su vez la dilución de las diferencias al analizar los valores de bioapatita sería producto de un alto consumo de plantas C3 por parte de todos los grupos analizados.
Otro dato relevante obtenido a partir de FRUITS, está asociado al consumo de recursos animales por parte de las poblaciones prehispánicas siendo en todos los casos no menor a un 30% o más del componente animal. Todos estos datos concuerdan con los relatos de los primeros cronistas que se adentraron en el río Paraná en el siglo XVI, como Diego García de Moguer (extraído de Medina 1908, pp. 244-245), Fernández de Oviedo y Valdés (1852, pp.191) y Ulrico Schmidel (1903 [1567]), quienes destacan el alto consumo de carne y pescado por parte de todos los grupos que habitaban esta área, así como también maíz por parte de los grupos guaraníes y chaná-timbués. Como plantean Bonomo et al. (2017), el escenario fue mucho más complejo con la presencia de economías heterogéneas entre los distintos grupos.
De acuerdo con lo que hasta aquí se ha planteado, la composición isotópica de los restos esqueletales de los individuos hallados en el sitio CIB presenta mayor afinidad con aquella de los grupos hortícolas. Dicha similitud se infiere a partir de la coincidencia observada en el patrón fotosintético dominante, tanto del componente proteico como energético de los alimentos consumidos.
Aunque solo tres individuos asociados a la Reducción Santiago de Baradero muestran una relevancia sustancial del maíz en su dieta, recurso que evaluamos como de menor calidad en términos alimenticios, resulta evidente que su importancia es mayor en términos generales respecto a los individuos prehispánicos. Esto concuerda con otras líneas de investigación, por ejemplo, un trabajo reciente de Auge y Tapia (2023) se identificaron almidones atribuibles al maíz en el tártaro dental del incisivo de un individuo de CIB. En definitiva, lo que se observa en las dietas inferidas tanto para el conjunto de CIB como para los horticultores es, por un lado, una pérdida en la variación respecto a los CRP. En el caso de la reducción, esto podría ser consecuencia del menor acceso periódico a zonas de pesca y recolección producto del mayor tiempo que le tendrían que haber dedicado a otras actividades, o bien debido a los límites espaciales y de movilidad impuestos por la colonización española. Una agenda de trabajo posible para el futuro sería cuantificar la pérdida de nutrientes que pudo ocurrir al reducirse la ingesta de recursos, tales como los peces y frutos recolectados y consumidos por las poblaciones del Paraná inferior en tiempos prehispánicos.
Por otro lado, se observa en los individuos de CIB una caída en la relevancia del consumo de recursos animales, al menos en relación a lo observado en las dietas prehispánicas de la región. Esto es especialmente evidente en aquellos casos donde se pudieron analizar molares y restos óseos de un mismo individuo. Si bien no es posible saber con precisión en qué momento ingresaron a la reducción, sí se observa una disminución en el consumo de proteína hacía sus últimos años de vida. Esto podría ser consecuencia de la escasa presencia de ganado que habría en la reducción, así como de la prohibición sobre su consumo, tal como se ha registrado en las fuentes escritas. No obstante, es probable que los individuos de CIB hayan incorporado recursos herbívoros consumidores de plantas C4 con mayor frecuencia en su dieta que lo que indican las fuentes documentales, como lo hicieran también los individuos horticultores prehispánicos. De todas maneras, con la información disponible resulta dificultoso dilucidar si, en el caso del CIB, se trata del consumo de este tipo de animal o si es el resultado de una práctica forrajera sobre las especies europeas introducidas. Pues, uno de los ejemplares de vaca muestra un aporte sustancial de plantas C4, siendo probable que se les haya dado algún tipo de suplemento a base de maíz durante el invierno. En este sentido, cabe mencionar que durante los tiempos de la colonia en Norte América el maíz no fue relevante para los europeos en términos de nutrición directa, sino que en buena parte quedó relegado a su uso como suplemento dietario animal, mientras que eran introducidos los cereales europeos para el consumo humano (Larsen, 1997). En la Argentina, esta estrategia también podría haberse dado desde tiempos coloniales tempranos como lo muestra el ejemplar de Bos taurus (EI7).
Debemos destacar que la aparente continuidad en el patrón alimentario que se observa entre horticultores prehispánicos y nativos residentes en Santiago del Baradero podría ser el resultado de una estrategia de cooptación de individuos pertenecientes a grupos indígenas que mantenían aquella forma de vida. Bajo esta premisa, los individuos con prácticas culturales que requerían un mayor grado de sedentarismo pudieron ofrecer menos resistencia para ser reducidos y amoldarse a la vida de la misión franciscana.
Consideraciones finales
La región del Paraná medio e inferior prácticamente no presenta especies endémicas debido a la juventud de su ecosistema y a la ausencia de barreras geográficas. Especies de linajes subtropicales, chaqueñas y paranaenses se adentran a través de los ríos Paraná y Uruguay y se distribuyen por los cursos de agua. También ocupan este espacio especies de la llanura pampeana, templada y mesopotámica. Esto provoca que exista gran variedad de especies de flora y fauna, una yuxtaposición de sus nichos y por ende una elevada interacción entre ellas (Malvárez, 1999; Quintana y Bó, 2010). Estas características del ambiente facilitaron la instalación de numerosos grupos humanos con distintas economías, principalmente la caza-recolección y pesca (con diferentes grados de importancia de cada actividad), y distintos grados de horticultura. El arribo de los grupos europeos, la instalación de las reducciones, la necesidad de mano de obra indígena y la generación de límites políticos, provocó la disrupción de los estilos de vida de las poblaciones nativas, entre los cuales se destacan el consumo de nuevos recursos y la readecuación de sus dietas.
En este trabajo nos propusimos analizar la dieta de los individuos de la reducción Santiago del Baradero a través del análisis de isótopos estables de carbono y nitrógeno, utilizando la información proveniente de las fuentes documentales disponible como una aproximación a la dieta y comparar estos resultados con aquellos de las poblaciones prehispánicas de la zona. Esta estrategia metodológica ya fue utilizada anteriormente para analizar la transición alimentaria de las poblaciones de Tierra de Fuego, como consecuencia de la ocupación europea; por ejemplo, en Tafuri et al. (2017) y Valenzuela et al. (2019). Las aproximaciones en ambos casos dieron resultados diferentes, exhibiendo continuidades en la alimentación en el primer caso y diferencias en el segundo. Más allá de las posibles discrepancias, que pueden deberse a las respectivas estrategias de análisis, ambas investigaciones muestran la relevancia de la introducción europea en la alimentación local, ya sea por la resistencia a los cambios (Tafuri et al., 2017) o a la adecuación a los mismos para la supervivencia (Valenzuela et al., 2019). Investigaciones similares se han realizado en Cuba (Lafoon et al., 2020) donde la transición de la dieta indígena a la del tipo europeo habría ocurrido en unos 50 años, lo cual sugiere que durante el sistema de encomienda se obtuvo un acceso rápido a los alimentos recientemente introducidos; particularmente se observó un mayor consumo de plantas C4 y la incorporación de cerdos a la dieta. Por otro lado, en un estudio realizado por Price et al. (2012), a partir de muestras recuperadas en un cementerio colonial en Campeche (México), los datos isotópicos indicaron una dieta rica en maíz similar a la dieta de momentos prehispánicos pero con un bajo consumo de alimentos marinos, si se considera que se trataba de un pueblo costero. Como estos datos son similares a los que presentan los grupos que vivían tierra adentro, se interpretó que los pobladores rurales estarían migrando a Campeche, impulsados por las medidas de reclutamiento europeo u obligados por el hambre.
A partir de estos casos se observa que los cambios nutricionales que ocurrieron en las poblaciones locales producto de la conquista hispánica varió de acuerdo a múltiples factores, como la economía y hábitos alimenticios previos, así como también a las necesidades generadas en los grupos nativos producto de las nuevas limitantes impuestas por los conquistadores.
Nuestros datos y las distintas aproximaciones analíticas indican que la dieta de los individuos CIB para los momentos coloniales tempranos se asemeja a la de los grupos horticultores prehispánicos, tales como los guaraníes y chaná, observándose a lo largo de los años una disminución en el consumo de proteínas y un incremento de alimentos C4. El maíz habría sido un componente importante en la dieta de la reducción, aunque en la mayoría de los casos no se basó únicamente en este alimento, apoyando la información hallada en las crónicas españolas, donde se detalla que en la reducción también se alimentaban de frijoles, trigo, yegua y pescado.
Aún quedan algunos interrogantes abiertos, como a qué se debe la similitud entre las dietas, en particular en lo que refiere al componente proteico. Si bien en las poblaciones prehispánicas se debería al consumo de recursos con valores de carbono elevados, en los individuos de la reducción habría distintas posibilidades. Por un lado, podría ser la consecuencia de alimentar a los animales de granja, como vacas o yeguas con maíz. Por otra parte, podría deberse a que en la reducción se introdujeron y consumieron carpinchos, al igual que en el caso de los individuos horticultores prehispánicos. También podría considerarse que el tiempo transcurrido desde el ingreso a la misión no haya sido el suficiente para generar un cambio en la señal isotópica de los individuos analizados.
Ciudad Autónoma de Buenos Aires, 8 de septiembre de 2023

