











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkSOBRE EL CULTIVO DEL TOMATE
Origen y domesticación
El tomate cultivado (Solanum lycopersicum L.) pertenece a la familia de las solanáceas que incluye a más de 3.000 especies, muchas con importancia económica como la papa, berenjena, petunia, tabaco y pimiento, entre otras. Es originario de América y las 12 especies silvestres relacionadas se encuentran distribuidas desde el centro de Ecuador, pasando por Perú hasta el norte de Chile como así también en las Islas Galápagos (Spooner et al., 2005). Estas especies crecen en una gran variedad de hábitats desde el nivel del mar a lo largo de la costa del Pacífico a más de 3.300 metros en los valles del lado occidental de los Andes, desde climas áridos a climas lluviosos. La gran diversidad encontrada en las especies silvestres se expresa tanto en características morfológicas, como fisiológicas y sexuales, y las convierte en recursos genéticos muy valiosos. La primera fase de domesticación habría ocurrido en manos de los primeros agricultores en Ecuador o el norte de Perú y luego selecciones adicionales habrían tenido lugar en México a partir de poblaciones predomesticadas y nuevamente en Sudamérica luego de una reintroducción (Razifard et al., 2020; Blanca et al., 2021). Desde Centroamérica, el tomate fue llevado a Europa por los españoles y luego se extendió por todo el mundo. Se considera que S. pimpinellifolium L. sería el antecesor silvestre más cercano del tomate cultivado y que S. lycopersicum var. cerasiforme es un intermediario evolutivo entre S. pimpinellifolium y el cultivado. Desde una perspectiva evolutiva, la domesticación y la selección artificial a manos del hombre indujo importantes cambios fisiológicos y redujo notablemente su diversidad genética. Las consecuencias más importantes de este proceso han sido la modificación del sistema reproductivo, el incremento del tamaño del fruto y la reducción de la base genética (Warnock, 1991).
Historia del mejoramiento del cultivo
Los primeros registros que describen los intentos de seleccionar los mejores tipos varietales fueron realizados en Europa en la segunda mitad del siglo XIX. Durante los primeros treinta años del siglo XX, se fundaron varias empresas de mejoramiento y producción de semillas de tomate que se establecieron por todo el mundo. El mejoramiento de las variedades adaptadas a la cosecha mecánica se inició en 1943, pero la liberación al mercado recién ocurrió en la década de 1960 (Rasmussen, 1968). Esta especialización del mercado ha llevado a la diferenciación de las variedades de tomate que hoy se producen (Sim et al., 2009). En la década de 1940 se liberaron al mercado los cultivares híbridos de tomate que garantizaban uniformidad de plantas y frutos, resistencia y altos rendimientos. Estudios recientes han demostrado la recuperación de la riqueza alélica a partir de la década de 1960 con la introgresión de genes para resistencia a enfermedades e insectos desde S. peruvianum L., S. pennellii Correll, S. chilense Dunal Reiche y S. habrochaites Knaap & Spooner. En la década de 1980 también se incorporaron genes alternativos para tamaño de fruto, color y sabor (Sim et al., 2012; Schouten et al., 2019).
En Argentina, recién en 1946 se comenzó un plan de mejora en tomate y según Gallardo (2012), es muy probable que las selecciones obtenidas de tomate platense (Cattáneo et al., 2020) y las realizadas en la EEA Tucumán, hayan sido las primeras resultantes de la aplicación de un método científico. En la actualidad, el país posee un perfil netamente importador de semillas de tomate. En tomate se comercializan dos tipos de estructuras genéticas: híbridos F1 o líneas puras o variedades. El desarrollo de nuevos genotipos ha quedado relegado a las universidades y las estructuras de I+D públicas, tal es así que de los 351 genotipos registrados en los últimos 10 años ante el Instituto Nacional de Semillas (INASE), sólo hubo 12 variedades desarrolladas en nuestro país. De estas, diez son variedades o cultivares mejorados cuyo objetivo de producción es la industria y la Institución que tiene los derechos de obtentor es el INTA y las dos restantes son cultivares mejorados para destinar la producción al consumo en fresco cuyo derecho de obtentor pertenece a la Facultad de Ciencias Agrarias de la Universidad Nacional de Rosario. Las semillas importadas satisfacen los requerimientos buscados por los productores locales pero, además del alto costo, son cultivares que fueron desarrollados para otros ambientes y condiciones de cultivo y, sobre todo, carecen de la calidad demandada por los consumidores.
Producción mundial y nacional de tomate
La producción mundial de tomates frescos ha sido liderada desde 1960 por China en Asia, por Italia y España en Europa y por los Estados Unidos, México y Brasil en el continente americano. La misma ha aumentado sostenidamente desde mediados de los años 1990 hasta superar en 2019 los 180 millones de toneladas a nivel mundial y los 6,5 millones de toneladas en Sudamérica. En nuestro país la producción se ha mantenido estable desde 1996 a la fecha en valores cercanos a 1,1 millones de toneladas anuales, cosechadas en unas 17 mil ha productivas (FAO STAT, 2021). Aproximadamente el 60-70% de la producción nacional se destina a consumo en fresco mientras que el 30-40% restante, a la elaboración de productos industrializados. Esta actividad está concentrada principalmente en la región de Cuyo y algunos años resulta deficitaria requiriendo generalmente de la importación de productos desde Chile e Italia. En cambio, el mercado interno nacional se mantiene provisto de tomates para consumo en fresco durante todo el año a pesar de la estacionalidad del cultivo. Esto se logra gracias a la amplia distribución de las zonas productivas que van desde Salta y Jujuy, al Sur de la provincia de Buenos Aires y a las diferentes estrategias de manejo aplicadas (Ministerio de Agricultura, Ganadería y Pesca, 2020).
El Cinturón Hortícola de Rosario (CHR) se destaca no sólo por su nivel de producción, sino también por la comercialización, ya que concentra y abastece a más de 2 millones de habitantes. Por convicción comunitaria o imposición legislativa, la zona productiva lindante o integrada a la ciudad de Rosario, así como también a otras ciudades del país en las que existen producciones hortícolas se han convertido recientemente a la agroecología. Este sistema de producción amerita la generación de nuevos conocimientos y desarrollos tecnológicos.
PROGRAMA DE MEJORAMIENTO PARA CALIDAD DE FRUTO EN LA UNIVERSIDAD NACIONAL DE ROSARIO
Este programa se inició en la década de 1990 para mejorar la calidad del fruto en tomate, con especial énfasis en incrementar la vida poscosecha y ampliar la variabilidad genética con la incorporación de genes silvestres al gran cultivo. La vida poscosecha (Vp), definida como los días que transcurren desde la cosecha del fruto cuando se visualiza el cambio de color en superficie hasta que el mismo es comercialmente inaceptable por presentar síntomas de deterioro tales como arrugamiento o exceso de ablandamiento, es un carácter de fundamental importancia cuando el fruto de tomate se destina al consumo en fresco. Las estrategias de mejoramiento para extender este carácter se basan en la utilización de parentales que portan mutaciones que alteran la madurez. Mutantes tales como el nor (non ripening), rin (ripening inhibitor) y cnr (colorless non-ripening) exhiben efectos pleiotrópicos indeseables sobre el color, la textura y la jugosidad del fruto incluso en condición heterocigota (Vrebalov et al., 2002; Manning et al., 2006; Wang et al., 2019). Otra estrategia que retrasa la maduración del fruto es el uso de la ingeniería genética que ha buscado modificar las proteínas de la pared celular (Brummell y Harpster, 2001; Meli et al., 2010; Uluisik et al., 2016; Yang et al., 2017) o incluso editar genéticamente estos genes mutantes por la metodología CRISPR/Cas9 (Yu et al., 2017). Aunque estas estrategias han tenido éxito científico, carecen de la aceptación del consumidor y no son aceptadas por leyes existentes en diferentes países (Ishii y Araki, 2016; Schmidt et al., 2020).
Hemos demostrado que los frutos de las formas silvestres de S. pimpinellifolium y S. lycopersicum var. cerasiforme tienen mayor vida poscosecha que los cultivares comerciales de tomate (aunque menor que los genotipos homocigotos para los mutantes de madurez del fruto tales como los ya mencionados nor y rin). Además, presentan para este carácter una acción génica de tipo aditivo lo que permite una respuesta a la selección (Zorzoli et al., 1998; Pratta et al., 2000; Rodríguez et al., 2006a; Rodríguez et al., 2010; Pereira da Costa et al., 2013; Cambiaso et al., 2019a). En este contexto se han desarrollado diferentes poblaciones a partir del cruzamiento interespecífico entre el cultivar argentino Caimanta de S. lycopersicum y la accesión LA0722 de S. pimpinellifolium: poblaciones F2, líneas endocriadas recombinantes (RIL, Recombinant Inbred Lines), Híbridos de Segundo Ciclo (HSC) y líneas casi isogénicas (NIL, Near Isogenic Lines) (Rodríguez et al., 2006b; Cabodevila et al., 2017; Di Giacomo et al., 2020).
Con la generación de cruzamientos entre estos padres y el avance de la selección generacional se han realizado distintas experiencias para dilucidar las bases genéticas involucradas en los caracteres que definen la calidad del fruto contando con la generación y utilización de datos genómicos, posgenómicos y bioinformáticos.
MÉTODOS DE MEJORAMIENTO UTILIZADOS
Obtención de Líneas Endocriadas Recombinantes (RIL) por el método genealógico.
La población base fue la F2 del cruzamiento interespecífico entre el cultivar Caimanta de S. lycoperscicum y la línea LA0722 de S. pimpinellifolium. El cultivar fue provisto por la Estación Experimental Agropecuaria de INTA Cerrillos, Salta, Argentina y la línea silvestre por el TGRC (Tomato Genetic Resources Center, Department of Vegetable Crops, University of California, Davis, USA). Para el proceso se utilizó el método genealógico (Hallauer, 1981). Al final de dicho proceso, se obtuvieron 18 líneas recombinantes (RIL) (autofecundadas F2:6) generadas a través de los distintos ciclos por selección antagónica - divergente de la vida poscosecha (Vp) y el peso (P) de los frutos (Zorzoli et al., 2000). El criterio de selección se basó en obtener genotipos con las combinaciones de ambos caracteres: 1) larga Vp y alto P, 2) corta Vp y bajo P, 3) larga Vp y bajo P, y 4) corta Vp y alto P. Así se obtuvieron estos 18 cultivares o RIL discrepantes para el peso, la vida poscosecha y otros atributos que hacen a la calidad de los frutos (Rodríguez et al., 2006 a, b) (Figura 1). Estos cultivares fueron caracterizados por RMN- 1H demostrando que S. pimpinellifolium amplió la variabilidad para la composición química de los frutos y para algunos metabolitos relacionados con el sabor y componentes nutricionales (López et al., 2015). Estas RIL son líneas homocigotas que tienen en su constitución genética fragmentos recombinados al azar del genoma de ambos progenitores a lo largo de los 12 pares de cromosomas. Estos 18 nuevos cultivares presentan mejores características de calidad de fruto que el progenitor cultivado del cual derivan y su tamaño y peso se ajustan al de los llamados tomates tipo cherry.
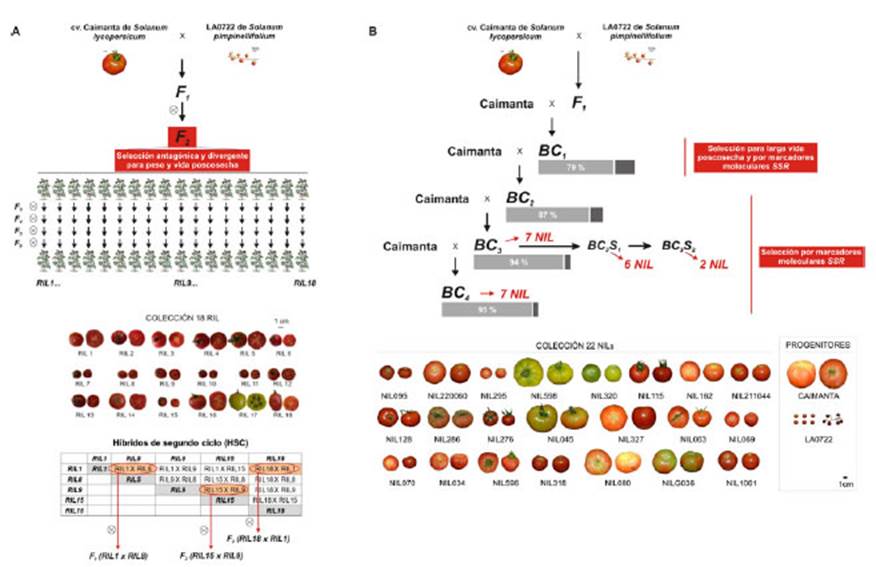
Figura 1 Esquemas de las metodologías de mejoramiento utilizadas en el Programa de mejoramiento para calidad de fruto de la Universidad Nacional de Rosario. A) Obtención de RIL (líneas endocriadas recombinantes) por el método genealógico aplicando selección antagonistadivergente para el peso y la vida poscosecha de los frutos. B) Obtención de NIL (líneas casi isogénicas) por el método de las retrocruzas aplicando selección para larga vida poscosecha y por marcadores moleculares del tipo SSR.
Esta población de RIL junto a sus progenitores fueron caracterizados por marcadores de ADN tales como AFLP (Amplified Fragment Length), SRAP (Sequence Related Amplified Polymorphism), microsatélites o SSR (Simple Sequence Repeats), InDel (inserción/ deleción), SNP (Single Nucleotide Polymorphism) y marcadores funcionales para la forma y el peso de los frutos (Pratta et al., 2011; Mahuad et al., 2013; Cambiaso et al., 2019a; Cabodevila et al., 2021). También fueron caracterizados por marcadores proteicos (Gallo et al., 2010). Todos estos estudios han permitido identificar polimorfismos de ADN y proteínas asociados a los cambios fenotípicos para la calidad de los frutos.
Obtención de Líneas casi Isogénicas (NIL) por retrocruzas
Inicialmente, se evaluó la BC1 obtenida de cruzar la F1 (Caimanta x LA0722) por el progenitor Caimanta. Todos los caracteres evaluados (Vp, P, forma, acidez, contenido en sólidos solubles, entre otros) mostraron una amplia variación, confirmando la diversidad genética entre los progenitores y la segregación posterior (Pereira da Costa et al., 2013). A partir de estos resultados se comenzó un proceso de detección, validación e introgresión, en el contexto genético del cultivar Caimanta, de QTL (Quantitative Trait Loci) asociados a caracteres de calidad de fruto de tomate aportados por regiones genómicas de la accesión LA0722 de S. pimpinellifolium. Se continuó con el proceso de introgresión obteniendo las generaciones BC3 y BC4, seleccionando en cada generación aquellas plantas en las que al menos un marcador SSR estuviera segregando. Actualmente se dispone de 22 NIL (Near Isogenic Lines), cada una de las cuales difiere con el progenitor Caimanta en un segmento cromosómico señalado por 22 marcadores SSR distintos (Figura 1). Las diferencias observables entre las NIL y el cultivar Caimanta (Di Giacomo et al., 2020) se deben exclusivamente a la región introgresada desde la especie donante o silvestre.
Cruzamientos dialélicos para el estudio de la variabilidad genética, el efecto recíproco y la heterosis en caracteres de fruto.
Cinco de las RIL seleccionadas por sus valores contrastantes fueron evaluadas en un diseño de cruzamiento dialélico (Híbridos de Segundo Ciclo, HSC) para el P, la Vp, el contenido en sólidos solubles y la acidez titulable, entre otros. La existencia de alta variabilidad entre las RIL y la aparición de distintos tipos de efectos génicos en los HSC, que incluyó el fenómeno de heterosis, demostró que la variación fenotípica observada responde en gran medida a componentes genéticos (Marchionni Basté et al., 2010). Estudios recientes que incluyeron cruzamientos entre tres de nuestras RIL y dos cultivares de origen estadounidense siguiendo un dialélico completo demostraron que la calidad y el contenido de metabolitos en los frutos de híbridos de tomate son afectados por la dirección del cruzamiento y la presencia de heterosis (Fortuny et al., 2021). En poblaciones segregantes derivadas de HSC se detectó variabilidad fenotípica y genética para varios caracteres de calidad de fruto demostrando que es posible continuar seleccionando genotipos para obtener nuevos cultivares con características fenotípicas superiores (Cabodevila et al., 2021).
Conservación y divulgación del germoplasma de Tomate (CodiGo TomATe).
La biodiversidad presente en las especies silvestres de tomate está aún subexplotada en los programas de mejoramiento del cultivo especialmente para enriquecer las bases genéticas con alelos nuevos que mejoren la productividad, calidad de los frutos y la adaptación del cultivo a distintas condiciones ambientales. Estudios recientes han detectado que en las especies silvestres de tomate existen 10 millones más de polimorfismos de una única base (SNP) comparado con la diversidad presente en la especie cultivada, es decir que la diversidad en el germoplasma silvestre es 20 veces superior (Aflitos et al., 2014). Para conservar y estudiar esta diversidad nuestro grupo está creando una colección de germoplasma que en la actualidad cuenta con 162 genotipos de tomate de diferentes especies y orígenes y con variabilidad para características de fruto. Entre estas tenemos 92 genotipos cultivados de S. lycopersicum y 70 especies silvestres (7 S. arcanum Peralta, 3 S. chilense, 5 S. corneliomulleri Macbr, 4 S. galapagense Darwin & peralta, 25 S. habrochaites, 2 S. pennelli, 5 S. peruvianum, 2 S. chmielewskii Rick, Kesicki, Fobes & Holle, 1 S. neorickii Rick, Kesicki, Fobes & Holle, 3 S. huaylense Peralta y 13 S. pimpinellifolium). Se multiplican, se cultivan y se recolectan datos fenotípicos e imágenes con los que se desarrolla el catálogo de variedades del Banco de Germoplasma de la Cátedra de Genética FCA-UNR. Las actividades de divulgación del grupo se ilustran en la Figura 2. Además, esta información junto a otras actividades académicas, de divulgación y experimentales se difunden a través de la página web www.codigotomate.com.ar.

Figura 2 Actividades para la conservación y divulgación del germoplasma de tomate. A) Frutos de especies silvestres y cultivadas de tomate presentes en el catálogo de variedades del Banco de Germoplasma de la Cátedra de Genética FCA-UNR. La línea blanca horizontal representa la escala de 1 cm. B) Entrega y trasplante de plantines durante jornada de extensión en huertas agroecológicas del periurbano de Rosario. C) Presentación del proyecto CodiGo TomATe en jornadas académicas.
CULTIVARES DESARROLLADOS
Cultivares inscriptos de tomate tipo cherry.
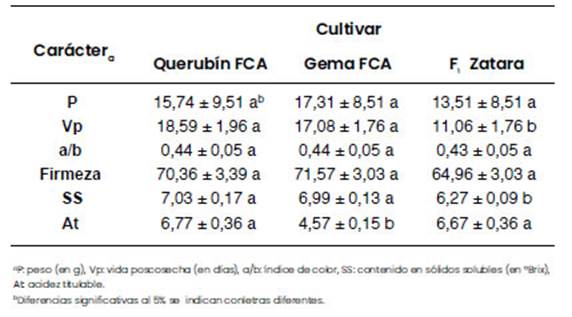
De las 18 RIL, dos líneas resultaron las más promisorias por sus características organolépticas, por lo que se han inscripto en el Registro Nacional de Propiedad de Cultivares (RNPC) del INASE. Ellas son: GEMA FCA y Querubín FCA (expedientes Nº 79929/12 y Nº 79928/12 publicados en el Boletín Oficial de la República Argentina Año CXX Número 32.449). En la Figura 3 se muestran frutos representativos de ambas líneas. Gema FCA y Querubín FCA presentan mayor vida poscosecha y contenido en azúcares en los frutos que el híbrido comercial Zatara (Florensa Argentina) de amplia difusión en nuestro país y utilizado en este ensayo como testigo. Gema FCA, además, produce frutos más ácidos (Tabla 1).

Figura 3 Frutos representativos de los cultivares tipo cherry Gema FCA y Querubín FCA y tipo redondo Dulcinea FCA y Matusalén FCA obtenidos en el Programa de Mejoramiento para calidad de fruto de tomate de la Universidad Nacional de Rosario.. La línea horizontal representa la escala de 1 cm.
Cultivares experimentales de tomate tipo redondo.
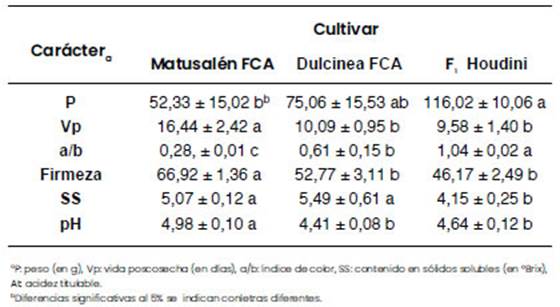
De las 22 líneas casi Isogénicas (NIL) de tomate, dos líneas resultaron las más promisorias por lo que se encuentran en evaluación para ser inscriptas en el RNPC del INASE. Ellas son: Matusalén FCA y Dulcinea FCA. En la Figura 3 se muestran frutos representativos de ambas líneas: Matusalén FCA y Dulcinea FCA producen frutos grandes, pero más chicos que el híbrido comercial Houdini utilizado como testigo (Florensa Argentina). Matusalén FCA presenta mayor vida poscosecha y Dulcinea FCA mayor contenido de azúcares en los frutos. Además, ambas poseen mayor contenido en azúcares y un color más anaranjado que el testigo (Tabla 2).
INCORPORACIÓN DE NUEVAS TECNOLOGÍAS AL PROGRAMA DE MEJORA
Las innovaciones tecnológicas en el área de la genética y el mejoramiento tales como los secuenciadores de última generación de genomas y transcriptomas completos, la edición génica, la adquisición de datos fenotípicos a gran escala (fenómica), entre otros, generan en la actualidad un crecimiento exponencial del volumen de los datos que necesitan nuevas metodologías para su análisis impactando en los programas de mejora. El análisis integral de estos datos no sólo permite ampliar los conocimientos básicos de los determinantes genéticos que hacen a la calidad del fruto de tomate, sino que directamente pueden ser la base para el desarrollo de nuevos cultivares con características mejoradas y adaptados a los sistemas productivos de nuestro país.
Hemos recientemente secuenciado el genoma completo del cv. argentino Caimanta y la entrada LA0722 de S. pimpinellifolium, los que fueron alineados al genoma de referencia de tomate cv. Heinz 1706 (The Tomato Genome Consortium, 2012). En base a las listas de polimorfismos se desarrollaron marcadores moleculares para construir mapas de ligamiento genético y detección de QTL en diferentes poblaciones del programa (Cambiaso et al., 2019 a,b; Cabodevila et al., 2021). En cuanto a estudios a nivel transcriptómico, se analizaron los transcriptos expresados en diferentes estados de madurez en los genotipos Caimanta y LA0722 por la técnica de cDNA-AFLP (Pereira da Costa et al., 2018) y por la técnica de RNA-seq (Cacchiarelli et al., 2021). A nivel proteómico hemos caracterizado genotipos uniformes y poblaciones segregantes por polipéptidos resueltos en 1-DE y 2-DE, para la detección de QTL para calidad de fruto y la identificación de un polipéptido asociado con cambios en el pH que mostró homología con una ATPasa cloroplastídica en los pericarpios de frutos (Rodríguez et al., 2008; Gallo et al., 2010, 2011; Pereira da Costa et al., 2014, 2017). En cuanto a fenómica, se ha avanzado en el uso del análisis a tres modos (Análisis Factorial Múltiple y Análisis Procrustes Generalizado) para caracterizar generaciones segregantes de híbridos de segundo ciclo y estimar en ellas una heredabilidad multivariada para caracteres de interés agronómico (Del Medico et al., 2019, 2020). Estas aproximaciones ómicas permiten identificar genes o familias génicas con roles fundamentales en la determinación de características organolépticas del fruto de tomate, conocer su nivel de expresión ante cambios ambientales o de contextos genéticos y retroalimentan el programa de mejoramiento para obtener genotipos superiores que satisfagan las demandas de productores y consumidores.
TRANSFERENCIA DE DESARROLLOS TECNOLÓGICOS AL MEDIO SOCIOPRODUCTIVO
La horticultura urbana y periurbana es una actividad que aporta múltiples ventajas territoriales ya que se asocia al mercado de cercanía, ideal para alimentos perecederos como lo es el tomate, genera puestos de trabajo, y los productores se convierten en potenciales custodios de los recursos naturales que utilizan contribuyendo además a preservar los espacios verdes frente al avance de la urbanización (Mitidieri y Corbino, 2012). Las huertas urbanas y periurbanas están demandando nuevas metodologías de producción; entre estas, nuevos cultivares, especialmente de polinización abierta y adaptados a sistemas de producción agroecológicos. Por esta razón, hemos iniciado un proyecto que busca una transferencia directa de tecnología de semilla de tomate hacia el medio socio-productivo y de conocimientos a partir de actividades de extensión y vinculación con los huerteros urbanos y periurbanos de ciudades importantes de nuestro país (Figura 2). De esta manera se contribuye a la soberanía alimentaria favoreciendo el acceso a semillas de cultivares de tomates desarrollados en instituciones públicas y permitiendo a los productores multiplicar las semillas para su propio uso. Los genotipos que se transfieren han sido mejorados para características organolépticas, resistencia a plagas y enfermedades y algunos de ellos tienen características nutricionales diferenciales en cuanto al tipo de azúcares, ácidos orgánicos y aminoácidos. Estas características nutricionales diferenciales, evaluadas en condiciones experimentales, se confirmarán mediante ensayos en sistemas reales de producción agroecológica.

