











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCTION
The available information on the human genome gives a starting point for searching in different populations and among individuals for genome variability related to pain sensitivity and to the effectiveness of different drugs in pain relief (Owusu Obeng et al., 2017; Crews et al. 2021). The human prodynorphin gene (PDYN), located on chromosome 20, encodes α-neoendorphin, β-neoendorphin, dynorphin A and dynorphin B. These molecules selectively activate kappa opioid receptors (KOR), encoded by OPRK1 gene, which is located on chromosome 8 (Schwarzer, 2009; Hashemi et al., 2018). Genetic association studies gave evidence of a link among certain DNA sequence variants of both genes and various pathologies, including cognitive disorders and drug abuse, as well as variations in pain sensitivity (Clarke et al., 2012; Hashemi et al., 2018; Nosova et al., 2021).
It has been reported that an efficient management of postoperative acute pain is essential not only for improving the wellness of the patient, but also for reducing the risk of chronicity of pain, morbidity and mortality (Carr and Goudas, 1999). Genetic polymorphisms can explain some of the variation in response to analgesics, while other important variables also involved are the sex of the patient, the intensity and kind of pain, the environmental influences, and several psychological aspects including among others, anxiety and somatization (Stamer and Stüber, 2007; Schreiber et al., 2014).
The challenge is to decipher the biological basis of such a complex phenotype, considering pain perception and response to analgesic drugs, both showing clear differences among populations of distinct origins. In Argentina, the current population is the result of several generations of intermixing among various groups at different times, including indigenous (Amerindian) communities, Spanish conquerors (early 1500s), Africans (arriving as slaves since the late 1500s until the end of slavery), and a large European immigrant population (arriving between 1870 and 1950) (Avena et al., 2006). In this work we analyse four different populations from Argentina, namely from Ciudad Autónoma de Buenos Aires (CABA), La Plata, Resistencia, and Misión Nueva Pompeya (MNP). Historical events Ciudad Autónoma de Buenos Aires (CABA) is the capital city of Argentina and it has by far the largest population in the country. During the second half of the 20th century, a significant demographic increment occurred in Argentina, mainly due to migratory flows (Gallo and Cortés Conde, 1967). From 1940 onwards, the industrial development encouraged people to move to CABA from other provinces of Argentina and from bordering countries bringing their indigenous genetic component (Torrado, 1992). While the European migratory contribution declined after 1930, immigrants from bordering countries are currently increasing the foreigner’s contribution to the city (Avena et al., 2001). La Plata is the capital city of Buenos Aires province. Located 56 km southeast from CABA, it is the fourth most populous city in the country. As in CABA, an important European contribution in the past century is currently complemented by the arrival of populations from bordering countries looking for employment (Cerrutti, 2009). In the case of Chaco province, it was originally inhabited by native people until 1528, when the first Europeans arrived. In 1872, a group of people from the province of Corrientes and Italian immigrants settled in this region. The city of Resistencia was then founded, and in 1884 it was assigned as the capital of the province of Chaco (De Pompert de Valenzuela, 2008; Tissera, 2008). At the end of the 19th century, European immigration to Resistencia was in order to promote urbanization and agricultural development (Maeder, 2012).
In 1900, the Franciscan missionaries founded the location of Misión Nueva Pompeya (MNP) in the western region of this province known as the Impenetrable Chaqueño. Currently, an important number of Native American people of the Wichí community still live in this inner region (Franceschi, 2010). A previous report on the urban people living in MNP estimated a native contribution of 25% uniparental genetic markers (Sevini et al., 2013). In fact, within the province of Chaco, Native American people from different communities live nearby several cities, and they still retain their traditional semi nomadic habits. The numerical importance of these native communities puts Chaco at present among the provinces with the highest number of living Native American people in Argentina (Instituto Nacional de Asuntos Indígenas, 2005). Previous studies have shown genetic differences between these four populations using non coding X chromosome markers (SNPs, INDELs and STRs) (Di Santo Meztler, 2018; Di Santo Meztler et al., 2019). Also, differences between a native Wichí community of Chaco and the population of Resistencia were found in the OPRK1 gene (Raggio et al., 2018).
In this work, our aim was to analyse whether interpopulation differences observed for non-coding genetic markers are also noticeable in coding regions, particularly for two genes of the opioid system and, therefore, to understand whether those differences have influence on the perception of pain. In particular, we focused on the genetic variability of PDYN and OPRK1 in four Argentinian populations. Genetic association with clinical variants influencing pain sensitivity was analysed for one of the populations, particularly after a surgical intervention.
MATERIALS AND METHODS
Populations
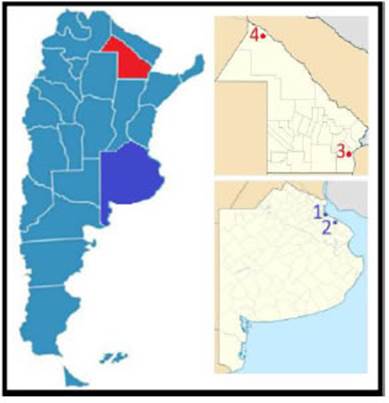
Between 2009-2012 we collected a total of 286 samples from adult, unrelated persons from four different locations in Argentina: the capital city of Argentina, CABA (n=106), the capital city of Buenos Aires province, La Plata (n=33), the capital city of Chaco province, Resistencia (n=96), and a small city of Chaco province, MNP (n=54). Figure 1 shows a map indicating these locations. Samples from La Plata, Resistencia and MNP consisted of both male and female donors, and were collected during three field trips. Samples from CABA were only female donors, which were collected at Gynecology Service´s Breast Pathology Section of the Hospital General de Agudos Bernardino Rivadavia. The intensity of perceived pain and the requirement of analgesia after gynecological surgery were recorded for 50 out of 106 females from CABA. After discarding individuals with one or more missing data, the genetic association study was performed for a sample size of 35 females. DNA was isolated from buccal and blood cells following protocols described in Gemmel and Akiyama (1996). OPRK1 data for the population of Resistencia were previously reported in Raggio et al. (2018) and were included in this work for comparison. All biological samples were genotyped by author G. P. Di Santo Meztler at the Instituto Multidisciplinario de Biología Celular (IMBICE). This study was part of a project previously approved by the Ethics Committee of the IMBICE, and all donors gave written consent for participation in the study.
Genetic determinations
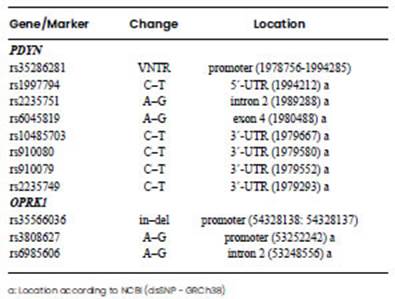
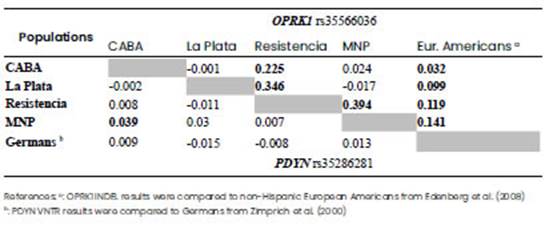
Eight polymorphisms (rs35286281, rs1997794, rs2235751, rs6045819, rs10485703, rs910080, rs910079, and rs2235749) were genotyped for PDYN gene, and three polymorphisms (rs35566036, rs3808627 and rs6985606) for OPRK1 gene (Table 1). Genotyping was performed by PCR and separation of amplified fragments in 1.8% agarose gels, except for rs10485703, rs910080, rs910079 and rs2235749, which were amplified together in a fragment by PCR and then sequenced. For rs35286281 VNTR polymorphism, alleles were designated as 1(271pb), 2(339pb), 3(407pb) or 4(475pb) based on the number of repeated elements that were identified. Such elements contain a transcription factor binding site that is associated with transcriptional efficiency of the human PDYN gene, and higher gene expression is associated with repeated alleles 3 or 4 (Zimprich et al., 2000). Therefore, the alleles 1 and 2 were categorised as low (L), and the alleles 3 and 4 as high (H) gene expression.
Primers for PDYN VNTR were obtained from Nikoshkov et al. (2008), primers for the SNPs located in the 3´ untranslated region (3´-UTR) of the gene were obtained from Yuferov et al. (2009), and primers for rs1997794, rs2235751 and rs6045819 were designed for this work (Supplementary Table 1). Primers for OPRK1 INDEL (rs35566036) were obtained from Edenberg et al. (2008) and allele specific primers for OPRK1 SNPs were designed in our lab and reported in Raggio et al. (2018).
Statistical analysis
Population study
For the genetic polymorphisms, allele frequencies were calculated using R v. 3.6.3 program (R Core Team, 2021). Heterozygosity, Hardy-Weinberg equilibrium (HWE) and genetic distance (as pairwise Fst values) were calculated through the program ARLEQUIN v.3.5 (Excoffier and Lischer, 2010), and linkage disequilibrium (LD) was calculated using the webtool SNPStats (Solé et al., 2006) for obtaining both D´ and r values. In the case of repetitive polymorphism rs35286281, the alleles were pooled into short (271 and 339) and long (407 and 475) given that extreme alleles (271 and 475) are very infrequent, so that pooling them would produce no bias. The adaptation of the tables for R program and ARLEQUIN was made using GA-TA program (https://github.com/santimda/GA-TA) (Gamboa Lerena et al., 2020). For population comparisons, data of four populations from 1000 Genomes database were included: Japanese in Tokyo (JPT); Mexican ancestry from Los Angeles (MXL), California, USA; residents of Utah with North and Western European ancestry (CEU) and Yoruba in Ibadan, Nigeria (YRI). Allele frequencies of these reference populations are detailed in Supplementary Table 2. Association study For the association study we considered only females from the sample of CABA without missing data (n=35). We analysed the variation of pain informed by the physicians in a follow-up of 1, 2, 12 and 24 h after surgery; although the observations were informed by different surgeons, all pain reports were registered under supervision of author J. Schiaffi. For this analysis two models were evaluated: in one case the dependent variable was the pain scale reported by the physician, who considered wound palpation, analgesia requirement, possibility of walking, and pain escalation according to medical impression (model M); while in the other model the scale was reported by the patient according to selfperception (model P). The independent variables used in both models were the above mentioned polymorphisms of PDYN and OPRK1 genes (Table 1), and the clinical variables time after surgery, age of the patient, dose of intravenous Klosidol (Bagó Laboratory, Argentina) -which consists of a combination of dextropropoxyphene hydrochloride and dipyrone, supplied every 8 h according to pain intensity-, associated pathologies, and type of incision (Table 2). The type of incision depended on the type of surgery, which was either gynecological surgery (Median Infraumbilical laparotomy, and Pfannenstiel laparotomy incisions) or breast surgery (Radian, Arcuate, and Stewart mastectomy -Orr type- incisions). The analyses were made using R v.3.6.3, and GEE (Generalised Estimating Equation) model fitting was performed with the geepack library (Halekoh et al., 2006).
RESULTS
Population genetic analysis
Allele frequencies for the PDYN and OPRK1 polymorphisms are detailed in Table 3, and genotype frequencies in Supplementary Table 3. In the case of MNP, for rs35286281 the frequency of genotype 339/407 was higher than in the other populations, and for rs6045819 genotype G/G was absent. In none of the populations the genotype C/C was observed for rs10485703. In the case of Resistencia, for rs35566036 the frequency of genotype del/del was higher and that of the genotype in/del was lower than in the other populations. For MNP, all the polymorphisms fitted HWE (p-value>0.05), but for the other three populations some of the markers did not fit HWE (Table 3). As we expected, linkage disequilibrium for PDYN gene was considerably high in all the populations. CABA and Resistencia showed higher LD than La Plata and MNP. For OPRK1 the LD was lower in Resistencia and La Plata, while in CABA and MNP there were significant values of LD for all the markers (Supplementary Table 4).
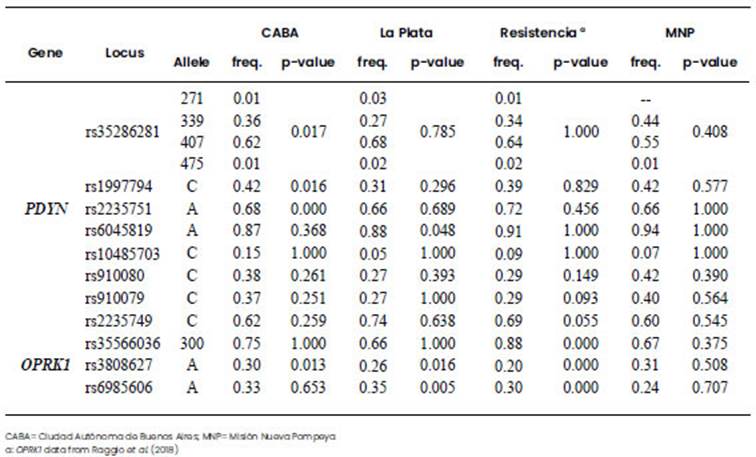
Table 3 Allele frequencies and Hardy-Weinberg Equilibrium p-values for the polymorphisms analysed. For biallelic polymorphisms, one of the allele frequencies is showed.
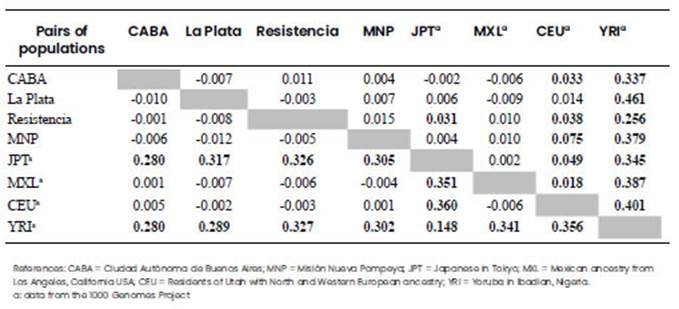
Table 4 Pairwise genetic distances (Fst values) for SNP markers among the analysed populations and data from the 1000 Genomes Project. Above the diagonal: Fst values for OPRK markers; and below the diagonal: Fst values for PDYN markers. Significant Fst values (p-value <0.05) are highlighted in bold.
The genetic distances were calculated for all the populations, and Fst with p-values lower than 0.05 were considered as significant (Tables 4 and 5). All four Argentinian populations showed significant differences with Africans from the Yoruba tribe in both genes, and also with Asians from Japan in PDYN, but differences in OPRK1 resulted only significant for Resistencia. On the contrary, OPRK1 resulted in significant values in the comparison with Europeans, while no differences were found when comparing with Mexicans for these two genes. Within Argentina, no differences were found for the SNPs comparisons (Table 4), whereas rs35566036 INDEL of OPRK1 showed a differentiation among Resistencia and the other three Argentine populations, and rs5286281 VNTR of PDYN resulted significant only for CABA-MNP (Table 5).
Association Study
GEE analysis
Two GEE models were used to evaluate the association of the reported pain scale with the following variables: age of the patient (Age), dose of intravenous dextropropoxyphene hydrochloride + dipyrone every 8 hrs (Ke8), associated pathologies (AP), type of incision (I), time after surgery (Time) and the genotypes for each polymorphism. Model M was based on the pain scale reported by the physician (MScale) and Model P on the pain reported by the patient (PScale). Thus,
• Model M-->MScale(Time, Age, AP, Ke8, I, genetic polymorphisms)
• Model P-->PScale(Time, Age, AP, Ke8, I, genetic polymorphisms)
For this study we used 7 out of 11 genetic polymorphisms that fitted HWE (p-value>0.05). We only focused on these polymorphisms for the association study in order to avoid statistical artifacts of markers out of HWE, probably given by the sample size, so that we can reach an accurate association between genetic markers and pain. The ANOVA p-values obtained with the GEE models are shown in Table 6, where p-values<0.01 were considered as significant values.
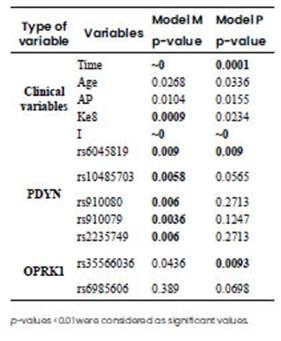
Table 6 A ANOVA p-values obtained for two models of Generalised Estimating Equation. Model M uses the pain scale reported by the physician as a dependent variable, while Model P uses the pain scale reported by the patient. Significant p-values (p<0.01) indicate the clinical variables and/or genetic variants that influence pain susceptibility.
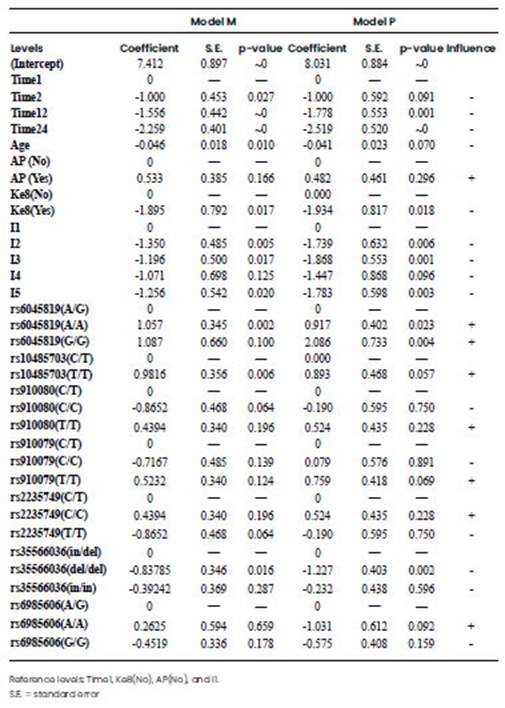
Table 6 B Influence of clinical variables (Time, Ke8, I, AP and Age) on the pain scale reported by the physician or the patient. Coefficients, deviation standard and p-values of the generalised estimating equation (GEE). For each variable, the influence of the levels in the sensitivity to pain is shown. Negative coefficients for a level indicates lower pain respect to the reference level (coefficient for reference equals to zero).
After analysing the results of the previous GEE models, three clinical variables presented a strong association with pain (p-value<0.01) for the model M: Time, Ke8, and I, while for Model P only Time and I were significant. Regarding genetic polymorphisms, 5 out of 7 were significant for model M while 2 out of 7 resulted significant for Model P (Table 6A). Once the variables influencing pain sensation were identified, the analysis was focused within each level of the clinical variables and polymorphisms, taking into account one level per variable as a reference. The reference level were 1 h after surgery (Time1), no analgesia rescue (Ke8(No)), Median Infraumbilical Laparotomy (I1), and no associated pathology (AP(No)), which were set at a coefficient value of zero (Table 6B). Thus, negative coefficients indicate pain decrease in comparison to the reference case. Postsurgical acute pain intensity was found to decrease as time increased in hours in both models, showing negative coefficients: Time 2(-1), Time 12(-1.556), Time 24(-2.259) for model M. The action of analgesia rescue (Ke8Yes) resulted in the same direction, showing a high negative coefficient. Additionally, an ANOVA analysis was performed considering Time separately for one surgical incision at a time. ANOVA results for the first and second hours after the intervention were non-significant, but differences emerged for the reports at 12 and 24 h after incision (data not shown). The Pfannenstiel, Radian, Arcuate, and Orr surgery incisions showed a higher decrease of pain scale, with respect to the reference type of incision (Median Infraumbilical Laparotomy), being Pfannenstiel, Radian and Orr, the types of incision that presented the most important decrease in pain scale in both models. Regarding genetic polymorphisms, in Model M the associated genotypes with lower pain sensitivity were rs6045819-A/G, rs10485703-C/T, rs910080-C/C, rs910079-C/C, and rs2235749-T/T, while for Model P the associated genotypes with lower pain sensitivity were rs6045819-A/G and rs35566036-del/del.
DISCUSSION
The genetic basis influencing postoperative pain through the screening for variations in the expression of genes coding for endogenous opioid system is a field of interest for improving pain therapies (Stamer and Stüber, 2007; Montes et al., 2015; Owusu Obeng et al., 2017; Crews et al., 2021). In this work we analysed the genetic diversity of PDYN and OPRK1 in Argentinian populations from different geographic locations, and the relationship between the genetic polymorphisms and the postsurgical pain, considering variables such as the use of analgesia and the different types of incisions. Our results show that the genetic background of the Argentinian population differs in some aspects from that of other countries and continents (Avena et al., 2006; Hohl et al., 2018). In a previous report, the variation of OPRM1 gene of the opioid system allowed to group Argentinians with other populations according to their ancestry, with 12.8% of differentiation among Africans, Asians, and European-Americans for this gene (López Soto and Catanesi, 2015). Genetic differences have also been found among different regions or provinces within Argentina, for several genetic polymorphisms, whether they were coding or non-coding markers (Corach et al., 2006; Avena et al., 2012; Di Santo Meztler et al., 2018; Muzzio et al., 2018; Sala et al., 2018; Caputo et al., 2021, among others). Interestingly, some differences emerged even when comparing Resistencia and MNP, which are located in the same province and only 425 km apart, as it was previously reported for non-coding markers (Di Santo Meztler et al., 2019). Our results showed less genetic differentiation among Argentinian populations, probably because the polymorphisms here analyzed are located in coding regions, having less chances for displaying variability. However, a significant Fst value was found between populations from CABA and MNP for the PDYN VNTR. This differentiation could be caused by the heterogeneous origin of immigrants from other continents, mainly Europeans from various countries who have settled in the past in particular locations along the territory of Argentina, as in CABA (Junta de Estudios Históricos del Municipio de Eldorado, 2015, 2016; Di Santo Meztler et al., 2018). As opposed, MNP is somehow isolated due to hard weather conditions and floods that discourage immigration.
Clinical variables
As several aspects of a surgical intervention can influence pain sensitivity, in this work we considered the interaction of both clinical variables and genetic variants. When considering the evaluation of pain scale, the physician integrated the knowledge of the patient and the type of surgery performed, the questioning, the medical examination, and the data of the medical record (such as use of analgesics, calls to the nurse, rescue medication, etc.). Therefore, we considered that an evaluation by the professional (pain scale reported by the physician) is more trustworthy, and, in fact, significant associations of three clinical variables were found when analysing the medical pain scale. Among the clinical variables to be taken into account, significant differences in reported pain scores were found according to the type of surgery. Although in the first two hours after the surgical intervention there were no differences between types of incision for pain scores, differences emerged as time increased, likely as a consequence of the severity of incision, being additionally influenced by biological factors as the genetic polymorphisms here analysed. Specifically, laparotomy incisions usually caused much higher pain scores on the first and second day after surgery than breast surgery incisions (either conservative or radical breast surgery). The results obtained in this work are in accordance with the grade of aggressivity of each incision. Pfannenstiel laparotomy involves the handling of the aponeurosis, thus giving a high pain sensation at the beginning but recovering faster in comparison to Radian, and even faster in comparison to Arcuate, and Orr incisions, which are less aggressive and usually cause a lower level of pain sensation from the beginning. On the contrary, Median laparotomy is the more aggressive, and the level of pain sensation likely persists more constantly along the first two days after surgery. Concerning the analgesic rescues, in general Klosidol was known to be well-tolerated as analgesia of choice for postsurgical pain in Latin American populations from Bolivia and Argentina, with a good balance cost/ benefit when it was prescribed for relief of postsurgical pain treatment (Daza Calderón et al., 2010). However, this combination of dextropropoxyphene and dipyrone was discontinued more recently because of some serious adverse effects that were reported for European populations (ANMAT, 2008). As expected, the analgesic rescues indicated in our study had a significant effect in decreasing the pain sensation during the first hours after surgery.
Genetic variables
Antinociception mediated by dynorphin and kappa receptors is known to be influenced by the sex of the patient, among other biological factors (Liu et al., 2013). Moreover, the effect of certain PDYN polymorphisms has been reported showing sexual dimorphism, with a higher impact on females (Clarke et al., 2012). Therefore, analysing genotype-phenotype association only in females avoids confounding results in this sense. Among the genetic variants that we analysed, an association with pain sensitivity in the physician model was observed for one SNP (rs6045819) in the exon 4 and four SNPs (rs10485703, rs910080, rs910079, and rs2235749) in the 3´-UTR of PDYN. Genotypes associated with higher pain sensitivity were GG for rs6045819 and TT, TT, TT and CC respectively for 3´- UTR polymorphisms. There is evidence that exon 4 could be involved in PDYN splicing. This is supported by the significant association of the risk allele G (SNP rs6045819) with alcohol and/or cocaine dependence (Xuei et al., 2006; Yuferov et al., 2009). This risk allele could form a non-canonical E-box, which is a target of binding transcription factors that could modulate PDYN transcription, thus increasing the expression levels (Taqui, 2011). Regarding SNPs in the 3´-UTR of the gene, they are located close to each other, resulting in a significant LD among them (Supplementary Table 4). This genetic linkage is stronger among rs910080, rs910079 and rs2235749, likely transmitted in a block. This result is consistent with the finding that rs910079 can be chosen as a reporter of the block (Yuferov et al., 2009). In addition, the haplotype rs910080-C / rs910079-C / rs2235749-T has been proposed to be associated with a lower level of gene expression (Yuferov et al., 2009). Several works show that dynorphins inhibit nociceptive transmission in the spinal cord via interaction with the kappa opioid receptor (Werz and Macdonald, 1985; Randic et al., 1995; Rusin et al., 1997; Wiley et al., 1997; Zachariou and Goldstein, 1997; Ogura and Kita, 2000). However, other authors have found evidence of dynorphin A having pronociceptive functions (Draisci et al., 1991; Dubner and Ruda, 1992; Riley et al., 1996; Vanderah et al., 1996et al., 1996; Wagner and Deleo, 1996; Laughlin et al., 1997; Malan et al., 2000; Laughlin et al., 2001). The switch between anti or pronociceptive effects of dynorphin A may depend on peptide concentrations, and kinetics of peptide interactions with either opioid or NMDA (N-methyl-D-aspartate) receptors. Dynorphins at physiological concentrations may be antinociceptive through the opioid receptors, typically playing an inhibitory role in acute pain conditions, whereas elevated pathophysiological levels may be pronociceptive and can interact with the NMDA receptors (Hauser et al., 1999; Tan-No et al., 2009). During peripheral inflammation, dynorphin induces its own synthesis through interaction with NMDA receptors, generating a regenerative, feedforward process (Laughlin et al., 2001). In our work, we found genotypes that are associated with a high pain sensitivity and, according to bibliography, induce the expression of PDYN. We suggest that an overexpression of PDYN after surgery, in particular in patients with these genotypes, is giving rise to an activation of NMDA receptors, causing increased sensitivity to pain.
Concerning OPRK1 gene, due to its wide presence in the central nervous system, its expression has been related to pain perception and behavioral traits as depression and drug abuse (Edenberg et al., 2008; Bruchas et al., 2010). The INDEL of OPRK1, rs35566036, was nearly significant for the model M. Different authors arrived to dissimilar conclusions on the importance of this polymorphism, either reporting a lack of association of this INDEL with the requirement of analgesia (Chatti et al., 2017), or finding a regulatory effect on gene expression in vitro for the longer allele (insertion), thus acting as a transcriptional promoter with effect on a complex phenotype of alcohol dependence (Edenberg et al., 2008). Our results are not conclusive for this matter, although the p-value near significance is suggestive of an influence of OPRK1 INDEL on pain sensitivity. Increasing the sample number is probably needed in order to obtain a more accurate result.
Several previous reports on PDYN and OPRK1 variation refer in general to dependence either on alcohol or drugs of abuse, given the abundance of dynorphin and kappa receptors on brain connections related to the formation of habits (Edenberg et al., 2008; Zhang et al., 2008; Dahl et al., 2018; Hashemi et al., 2018) and to emotional processing (Xu et al., 2013). Other reports consider the influence of PDYN and OPRK1 variants on pain modulation, mainly focused on chronic pain (Rosen et al., 2000; Wang et al., 2001; Podvin et al., 2016; Tian et al., 2018). Our results give an approach to the influence of the variation of both genes in pain, and suggest an association with levels of acute pain sensitivity and hyperalgesia after surgical intervention.
Concluding remarks
This is the first report on Argentinian population for PDYN variation, while information available on OPRK1 variation and pain sensitivity in the same population is scarce (Raggio et al., 2018), an unfavorable scene given the geographic extent and the heterogeneity of Argentinian people. The results presented in this work show differences between Argentinians and populations from other continents, even in the comparison to Europeans, suggesting that a component of admixture with Native American people probably reinforce the differences. This, however, cannot be confirmed due to the scarce available information for Native Americans on variation of the genes of the endogenous opioid system (Ehlers et al., 1998; Raggio et al., 2018) and other genes related to pain perception (Catanesi and Glesmann, 2015; López- Cortés et al., 2020). For this reason, an analysis on other populations of the region with known admixture with Native communities is needed. Although the number of individuals included in the analysis needs to be further increased, a genetic association with postsurgical acute pain phenotype has been found. These findings highlight the importance of a regional study of genetic variants influencing pain sensitivity and analgesic response, in tune with the current tendency of a personal therapy medicine.

