











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Las costras biológicas del suelo (CBs) son comunidades de cianobacterias, hongos, musgos y líquenes agregados con partículas de suelo que habitan los primeros 5 cm del perfil. Se encuentran ampliamente distribuidas en todas las zonas áridas y semiáridas del mundo, donde pueden cubrir más del 70% de la superficie del suelo entre plantas (Na vas et al., 2020; Belnap et al., 2016). Su presencia en los ambientes áridos puede contribuir en gran medida al manteni miento y redistribución del agua, influyendo en el funcionamiento y en la diná mica ecosistémica (Garibotti et al., 2018; Chen et al., 2019). Los efectos de las CBs sobre las condiciones hidrológicas puede ser el aspecto más importante, porque el agua es el principal factor limitante en las tierras secas y las CBs influyen tan to en su propia dinámica como en la de las plantas vasculares en dimensiones verticales y horizontales (Chamizo et al., 2016a; Ludwig et al., 2005; Chen et al., 2018). Las CBs modifican la respues ta hidrológica de los suelos donde apa recen, controlando la transferencia de agua, nutrientes y sedimentos de áreas desnudas a áreas con vegetación (Belnap et al., 2016). A pesar de su reconocida importancia, el papel de las CBs en los procesos hidrológicos, en comparación con los suelos desprovistos de ellas, no se comprende aún completamente (Whit ney et al., 2017; Li et al., 2014).
Entre las funciones hídricas afecta das por la presencia de CBs se pueden mencionar la infiltración y la retención de humedad en el suelo (Belnap et al., 2016). A pesar de los numerosos estu dios en infiltración, existen pruebas con tradictorias sobre el papel que juegan las CBs en la regulación del flujo de agua en los suelos (Zhao & Xu., 2013; Belnap et al., 2013). Maestre et al. (2002) y Belnap et al. (2013) observaron una mayor in filtración en áreas con CBs en compa ración con las áreas sin CBs; contraria mente, Yair (2003) y Zhao & Xu (2013) demostraron reducciones en la infiltra ción en suelos cubiertos de CBs. La re tención de humedad ha recibido menos atención, concentrándose la mayoría de los trabajos en contenidos de humedad absolutos de agua en el suelo, más que en la capacidad de las CBs por conservar la humedad del mismo (Xiao & Hu, 2017). En general se observa que la presencia de CBs contribuye a la conservación de la humedad en el suelo (Belnap et al., 1996; Seghieri et al., 1997). En todos los casos, la discrepancia en los resultados podría estar relacionada con la diversidad de metodologías utilizadas, sumado a la condición de sitio, suelo y tipo de CBs.
Los diferentes tipos de CBs (domina das por cianobacterias, algas, líquenes o musgos) pueden mostrar propiedades distintas, como la capacidad de reten ción de agua, el espesor y la rugosidad, que generalmente son bajas en CBs de cianobacterias y relativamente altas para las CBs dominadas por musgos y líque nes, afectando así los procesos hidroló gicos del suelo (Chamizo et al., 2016a; Whitney et al., 2017; Belnap y Lange, 2003; Li, 2012). Estudios en las dunas del norte del Negev demostraron que la composición específica tuvo efectos drásticos en la infiltración y el conteni do de humedad del suelo (Kidron et al., 2003). Líquenes y musgos generalmente tienen una mayor capacidad para influir en el equilibrio hídrico del suelo que cia nobacterias y algas, y por lo general re distribuyen más el agua de lluvia en las capas poco profundas del suelo (Xiao et al., 2015). En consecuencia, las CBs do minadas por líquenes y musgos pueden ser desventajosas para los arbustos de raíces profundas mientras que las CBs de cianobacterias y algas pueden no limitar el crecimiento de los arbustos (Breckle et al., 2008; Veste et al., 2011; Li et al., 2014; Whitney et al., 2014).
La influencia de las CBs en la infil tración puede quedar enmascarada por propiedades del suelo como la estruc tura o la textura. Williams et al. (1999) informaron un efecto significativo de las CBs dominadas por cianobacterias sobre las propiedades hidrológicas de un sue lo franco-arenoso (Yair et al., 2011). Por otra parte, el aumento de la porosidad, la estabilidad mejorada de los agregados y una estructura física mejorada causan una mayor infiltración (Menon et al., 2011; Rossi et al., 2012), mientras que la repelencia al agua y la obstrucción de los poros la reducen (Malam Issa et al., 2009).
A pesar de los avances recientes en la comprensión de cómo la CB influye en los procesos hidrológicos, todavía se tie nen que desarrollar predicciones defini tivas de cómo el tipo de suelo y las ca racterísticas de la CBs interactúan para influir en estos procesos. Nuestro objeti vo fue evaluar la influencia de diferentes grupos funcionales de CBs en la infiltra ción y la retención de humedad en tres texturas diferentes de suelo. Hipotetiza mos que las CBs modifican la tasa de in filtración y la retención de humedad del suelo y que esta modificación varía con la textura del suelo y el tipo funcional de CBs.
Material y Método
Se recogieron tres tipos de suelo con distribuciones de tamaño de partículas contrastantes, de tres zonas semiáridas de la Argentina que pertenecen a la Pro vincia Fitogeográfica del Monte. Desde los más finos hasta los más gruesos, estos suelos se clasificaron en: (1) Limoso (L), (2) franco-arenoso (FA), (3) arenoso (A). La distribución del tamaño de las partí culas para cada tipo de suelo se muestra en la Tabla 1.
Tabla 1: Comparaciones por pares y diferencias significativas dentro del factor tipo de CBs. Los asteriscos (*) indican diferencias significativas. Table 1: Pairwise comparisons and significant differences in the type of CBs factor. Asterisks (*) indicate significant differences
Los suelos muestreados se secaron al aire y se tamizaron a 2 mm, y luego se esterilizaron en autoclave dos veces durante 20 min a 120 °C para suprimir la actividad biológica. El suelo limoso se recolectó en la parte suroeste de la provincia de San Juan (32°00′8.43″ S; 68°45′10.18″ W) a 1139 m s.n.m. La precipitación media anual es de 370 mm y la temperatura media de 18 °C con un mínimo de 6 °C y un máximo de 20,7 °C. Las comunidades de Zuccagnia punctata Cav., Baccharis salicifolia Ruiz y Pav. y Baccharis pingraea D.C. son dominantes en esta área. En este sitio las comunida des de CBs están dominadas por mus gos como Pseudocrossidium arenicola (Dusen) M.J. Cano, Pseudocrossidium chilense R. S. Williams y líquenes como Enchylium coccophorum (Tuck.) Otálora, P.M. Jørg. y Wedin, Placidium lachneum (Ach.) Breuss, siendo el valor medio de cobertura para CBs de 21,1% (Navas Ro mero, 2019). Los suelos franco-arenosos se recolectaron en el distrito de Capde vile (32°43′24.3″ S, 68 ° 50′29.69″ W), Mendoza. a 741 m s.n.m., en el área mor foestructural de la precordillera. La pre cipitación media anual es de 220 mm y el 38% de la precipitación ocurre duran te la temporada de verano (diciembre-febrero). La temperatura media anual es de 17,5 °C con un valor máximo de 30 °C y un valor mínimo de 3 °C. Larrea cuneifolia es la especie dominante. En este sitio las comunidades de CBs son abundantes y están dominadas por mus gos como Pseudocrossidium arenicola, Pterygoneurum ovatum (Hedw.) Dixon, Bryum argenteum Hedw., líquenes como Psora decipiens (Hedwig) Hoffm., Enchylium coccophorum y cianobacterias, en particular el género Microcoleus sp., Oscillatoria sp. y Nostoc sp., siendo el va lor medio de cobertura de CBs de 42,3% (Navas Romero, 2019). El suelo arenoso se recogió en Los Médanos Grandes, en la provincia de San Juan (31°47′10.13″ S, 67°58′55.75″ W), en el lado este de la sierra Pie de Palo a 729 m s.n.m. La precipitación media anual es de 103 mm y se produce principalmente a partir de diciembre (principios de verano) a mayo (otoño). La temperatura promedio anual es 18 °C; con un máximo de 40 °C y un mínimo de 10 °C. La vegetación do minante está formada por arbustos de Bulnesia retama y Larrea divaricata. En este sitio, las comunidades de CBs están dominadas por musgos como Pseudo crossidium arenicola, Bryum argenteum y cianobacterias, en particular el género Scytonema sp. y Oscillatoria sp., siendo el valor medio de cobertura de CBs del 25% (Navas Romero, 2019).
En cada sitio se colectaron tres tipos funcionales de CBs: dominadas por mus gos, por cianobacterias y por líquenes. La forma de colecta dependió de la metodo logía empleada para la evaluación de la retención de humedad y de la infiltración.
Para determinar la función de las CBs como retentora de humedad se colecta ron 360 muestras de tipos funcionales distintos de CBs: 90 musgos (domina dos por Pseudocrossidium arenícola), 90 líquenes (dominados por Enchylium coccophorum), 90 cianobacterias (do minados por Scytonema sp.) y 90 con evidencia visual de suelo desnudo (SD). Las muestras se recogieron con un sa cabocado de 6 cm de diámetro después de humedecer la superficie y se coloca ron en recipientes plásticos de 5,5 cm de diámetro x 4 cm de profundidad. En la boratorio se pesaron con balanza digital en seco (precisión 0,002 g) y se les incor poró agua destilada hasta capacidad de campo; se mantuvieron a 25 °C y expues tas a 12 h luz/12 h oscuridad y se pesaron cada 24 h hasta peso constante.
Para determinar el efecto sobre la tasa de infiltración se colectaron los mismos tipos funcionales de CBs, así como de SD en igual cantidad de réplicas que para la determinación de la retención de hume dad. Para evaluar la tasa de infiltración se empleó el método descripto por Zaady et al. (1999). Se extrajo una muestra cir cular de cada tipo funcional de CB con una caja de Petri (9 cm diámetro, 2 cm profundidad) con la ayuda de un cincel y una espátula intentando minimizar la alteración de la superficie del suelo re cogido. Previo a la recolección de mues tras, se humedeció la superficie con agua destilada con un pulverizador. Las CBs se trasladaron a laboratorio y se mantu vieron en una incubadora durante 2 días en condiciones constantes (irradiancia de 3 μSm- 2s-1 y temperatura de 20 °C). En laboratorio, se realizaron 5 perforaciones de 1 mm en la cara inferior de cada caja de Petri. Las muestras colectadas fueron saturadas hasta capacidad de campo. Luego de 24 horas, las muestras se in sertaron en el centro de contenedores de 20 cm de altura y se incorporaron 100 ml de agua destilada en cada caja. La experiencia se extendió durante 5 minutos. La cantidad de agua colectada después de 5 minutos se empleó para calcular la tasa de infiltración en cada muestra.
Previo a los análisis estadísticos se aseguró la independencia de los datos, la normalidad se comprobó mediante la prueba de Kolmogorov-Smirnov. La homocedasticidad, mediante la prueba de Levenne. Para todos los análisis el nivel de significancia fue de α= 0,05.
Para determinar si existían diferencias significativas en la tasa de infiltración entre las tres texturas de suelos y entre los grupos funcionales de CBs (musgos, líquenes, cianobacterias) se realizó un ANOVA de dos factores, considerando textura, tipo de CBs y su interacción.
Para determinar si existían diferencias significativas en la retención de hume dad entre las tres texturas de suelos y en tre los grupos funcionales de CBs (mus gos, líquenes, cianobacterias) se realizó un ANOVA de medidas repetidas.
Todos los gráficos se construyeron con el software “SigmaPlot v.11” (SigmaPlot, 2008). Los análisis se efectuaron con el software “Infostat v.16” (Di Rienzo et al., 2016) y SPSS Statistics 17.0.
Resultados
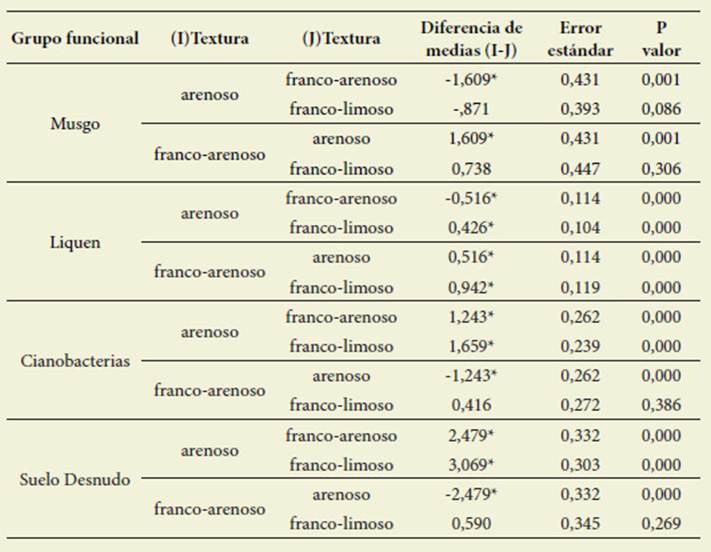
La tasa de infiltración mostró un com portamiento diferente según la textura del suelo y el grupo dominante en la CB. El ANOVA realizado con los datos de los factores “textura” y “tipo de CBs” tuvie ron efectos significativos sobre la tasa de infiltración, al igual que la interacción de los dos factores. Para la textura FA todos los tipos de CBs produjeron un aumento en la tasa de infiltración con respecto al SD y la mayor tasa de infil tración la presentaron las CBs domina das por musgos (p<0,0001) (Figura 1, Tabla 2).
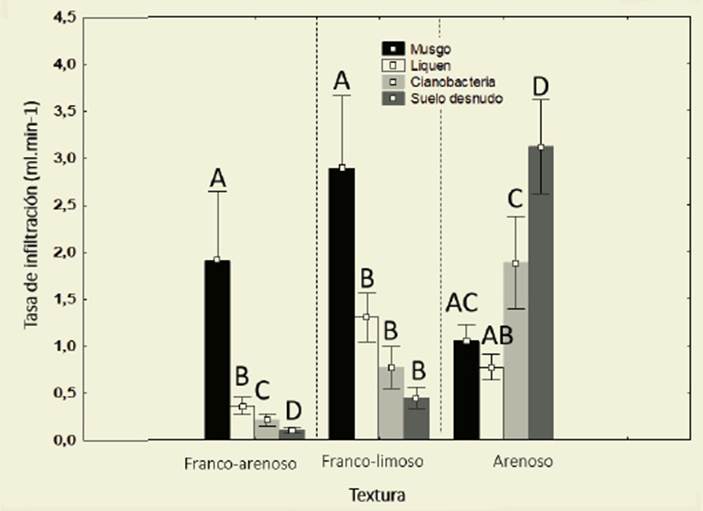
Figura 1: Valores medios de tasa de infiltración (ml.min -1 ) para los tres grupos funcionales do minantes y para el suelo desnudo en las tres texturas de suelo evaluadas. Las cajas representan los valores medios y las barras el desvío estándar. Letras distintas indican diferencias significati vas entre grupos funcionales dominantes (α=0,05) Figure 1: Mean values of infiltration rate (ml.min-1) for the three dominant functional groups and for bare soil in three soil textures evaluated. Box indicate mean values and bars standard deviation. Different words indicate significant differences between dominant functional groups (α=0-05)
Tabla 2: Comparaciones por pares y diferencias significativas dentro del factor textura. Los asteriscos (*) indican diferencias significativas. Table 2: Pairwise comparisons and significant differences in the texture factor. Asterisks (*) indicate significant differences

Para la textura L solo las CBs dominadas por musgos aumentaron la tasa de infiltración con respecto al SD (p<0,0001) (Figura 1, Tabla 1). Para la textura A la presencia de CB generó una reducción de la tasa de infiltración y la mayor tasa de infiltración fue para SD ( =3,23±2,34; p<0,0001) (Figura 1, Tabla 1). La comparación por pares para tipos de CBs mostró diferencias signifi cativas al comparar las texturas. Los mus gos tuvieron la mayor tasa de infiltración en la textura FA ( =2,189±2,41), líquenes en el L ( =0,36±0,29) y cianobacterias ( =1,89±1,69) y SD ( =3,12±2,34) en la A (Tabla 2).
En los tres tipos de textura evaluados la pérdida porcentual de agua fue más abrupta en el SD. Las tendencias en el tiempo fueron significativamente dife rentes entre los distintos tipos de CBs y los suelos con distinta textura. Las CBs mostraron un comportamiento diferente de acuerdo al grupo que dominó en cada textura. En todos los casos los musgos retuvieron el agua por más tiempo. En el sitio L las CBs dominadas por musgos y líquenes y el suelo desnudo perdieron el 50% del agua a las 96 h de iniciado el ex perimento, mientras que las dominadas por cianobacterias a las 72 h (Figura 2).
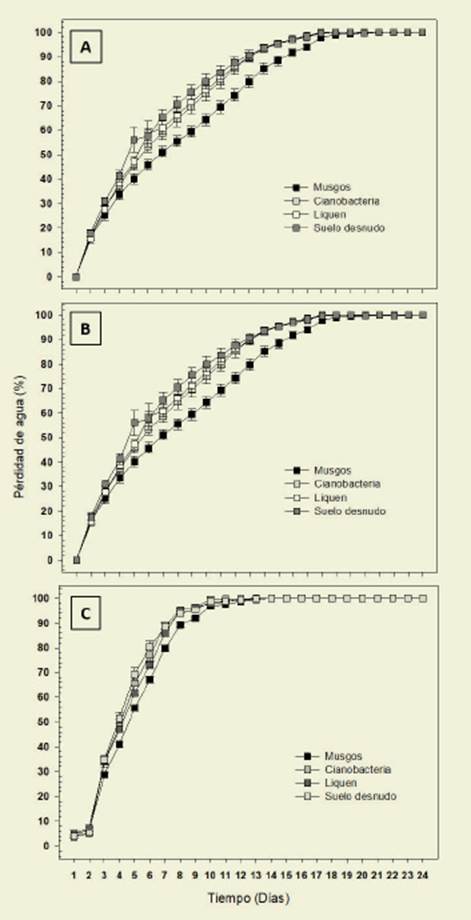
Figura 2: Valores medios de pérdida de agua (%) a lo largo del tiempo (días) para los tres grupos funcionales dominantes y para el suelo desnudo, en las tres texturas de suelo evaluadas. Los símbolos representan los valores medios y las barras el desvío estándar. A-Franco-arenoso, B-Limoso, C-Arenoso. Figure 2: Mean values of water loss (%) during time (days) for the three dominant functional groups and for bare soil in the three soil textures evaluated. Symbols represent mean values and bars standard deviation. A- Franc-sandy, B- Silty, C- Sandy
En el sitio FA el SD perdió el 50% del agua a las 72 h de iniciado el experimen to, los líquenes a las 96 h y los musgos a los 144 h. Los musgos perdieron el 100% del agua a las 480 h de iniciado el expe rimento, los líquenes y el SD a las 408 h (Figura 2). En el sitio A el SD y las cia nobacterias perdieron el 50% del agua a las 96 h de iniciado el experimento y los musgos a los 120 h. Las cianobacte rias y el SD perdieron el 100% del agua a las 240 h de iniciado el experimento, mientras que los musgos a las 312 h (Fi gura 2).
Discusión
Con excepción del suelo con textura A, la presencia de las CBs generó un aumen to en la tasa de infiltración (Figura 1). Coincidente con estos resultados, Kakeh et al. (2018) en zonas con Cbs en Irán encontraron que las tasas de infiltración fueron aproximadamente dos veces más altas en las superficies con CBs que en las superficies desnudas. El comportamiento inverso en la tasa de infiltración obteni do para el sitio A puede estar relacio nado con la textura del suelo. En suelos arenosos con grandes poros la presencia de CB actuaría impidiendo la infiltración rápida y mantendría más tiempo la hu medad del suelo al menos en la capa superficial, evitando la perdida rápida del agua característica de suelo arenosos, lo que permitiría aprovechar inclusos pul sos cortos de precipitación, propios de las zonas áridas evaluadas (Rodríguez- Caballero et al., 2015). En las CBs, al hu medecerse, las vainas de algunas ciano bacterias absorben hasta ocho veces su peso en agua y aumentan varias veces su volumen original, bloqueando aún más el espacio disponible de los poros cerca de la superficie (Belnap & Gardner, 1993; Kidron et al., 1999). Por lo tanto, nues tros resultados confirman que la presen cia de CBs en suelos arenosos reduce la tasa de infiltración, en relación con los suelos sin CBs. Por el contrario, los sue los arcillosos tienen abundantes poros, pero forman microcanales donde la ten sión superficial, el efecto adhesión, las fuerzas de tensión-gravedad, además de la estructura de la partícula, contribuyen a disminuir la infiltración, por lo que en este tipo de textura la presencia de CBs favorecería la infiltración. Nuestros re sultados concuerdan con Warren (2003) y Schulten (1985) para quienes las CBs mejoran la agregación de partículas finas del suelo y generan un efecto positivo sobre la infiltración. Graetz & Tongway (1986) encontraron en suelos francos arenosos, en matorrales de quenopodiá ceas en el sur de Australia, que la tasa de infiltración final en zonas con CBs fue menor a 10 mm·h-1 en comparación con 45 mm·h-1 en una superficie adyacente sin CBs. Se obtuvieron resultados simi lares en la reserva de Koonamore en el sur de Australia, donde la presencia de CBs en un suelo arenoso redujo la tasa de infiltración a 70 mm·h-1 en comparación con 140 mm·h-1 en un área cercana de suelo similar sin CBs (Rogers, 1977). En nuestro estudio, a pesar de que en el si tio A las CBs redujeron la infiltración, la ausencia de la CBs permite mayor expo sición de la capa superficial del suelo al agua, produciéndose una redistribución de las partículas finas en el perfil del sue lo generando así una obstrucción de los poros y el desarrollo de un sello super ficial (Danin, 1983; Bresson & Valentin, 1993). Por otro lado, el incremento en la rugosidad que supone la presencia de las CBs disminuye el escurrimiento superfi cial, reteniendo el agua por más tiempo y facilitando una mayor infiltración neta (Navas et al., 2019).
Los musgos presentaron las tasas de infiltración más elevadas (Figura 1). Coincidiendo con Mücher et al. (1988) y Chamizo et al. (2012) que encontra ron que las tasas de infiltración se incrementaban con el nivel de sucesión de las CBs, siendo las CBs dominadas por musgos (los de mayor nivel sucesio nal) las que presentaron mayor nivel de infiltración. El incremento en las con centraciones de sustancias poliméricas extracelulares (SPE) de origen microbia no en los musgos podría ser una de las causas de su mayor infiltración (Belnap et al., 2001). Las SPE pueden desempe ñar un papel importante en la mejora de la infiltración, probablemente debido a su capacidad para impartir una textura “esponjosa” a los agregados constituidos por partículas de suelo y polímeros. Por su parte en el sitio A, la mayor rugosi dad local dada por las CBs dominadas por cianobacterias (Navas et al., 2019) aumentaría la cantidad relativa de agua en ese lugar incrementando así las tasas de infiltración en relación a las domi nadas por musgos (Miralles et al., 2011; Chamizo et al., 2012). Las tasas de infil tración más bajas en algunas ocasiones dadas por el liquen pueden estar relacio nadas con la naturaleza de las especies presentes en los diferentes sistemas y en la capacidad de algunas especies de sellar completamente la superficie, bloquear el acceso a los poros del suelo (Warren, 2003) e impedir el paso del agua. Según Chamizo et al., (2012) las CBs de líque nes poseen una alta porosidad por el dominio de poros alargados; sin embar go, al estar separada la CB del sustrato subyacente no hay continuidad entre los poros de la CB y del sustrato afectando la infiltración. Por otra parte, las bajas tasas de infiltración también pueden atribuir se a las propiedades hidrofóbicas de los líquenes (Souza-Egipsy et al., 2002) y a su capacidad para bloquear los poros del suelo cuando están húmedos (Warren, 2003). Estas bajas tasas de infiltración en líquenes son consistentes con los resul tados encontrados por diversos autores (Alexander & Calvo, 1990; Chamizo et al., 2012).
Las CBs lograron retener por más tiempo el contenido de agua (Figura 2). Esta capacidad estaría relacionada con el incremento en el contenido de materia orgánica propia de los organismos cons tituyentes de las CBs y con los atributos morfológicos específicos de los organis mos poiquilohídricos que la componen. Así, la capacidad de las vainas de las cia nobacterias de incorporar hasta 8 veces su volumen en agua, de los musgos de expandir sus hojuelas e hidratarse rápi damente frente a un pulso de agua, o de disminuir la pérdida de agua a través de la presencia de costas excrecentes, cera en las hojuelas, o reducción instantánea de sus actividades, parece ser una de las explicaciones más plausibles para la re tención. Nuestros resultados coinciden con los obtenidos por Sun et al. (2004), quienes al evaluar los efectos de las CBs sobre el contenido de agua del suelo encontraron que este fue de 10,3 a 16,8 veces superior en las zonas con CBs. Ro driguez-Caballero et al. (2012), hallaron que la retención de agua de la capa su perior del suelo y el almacenamiento de agua en la superficie del suelo aumenta ron con la presencia de CBs.
Los musgos fueron los que retuvieron el agua por más tiempo (Figura 2). Re sultados similares fueron obtenidos por Chamizo et al. (2012) quienes hallaron un incremento en los niveles de agua en CBs dominadas por musgos. La capaci dad de los musgos de absorber agua di rectamente a través de los pelos en sus hojas y expandir su cobertura y biomasa hasta 13 veces, a diferencia de los otros grupos, es una de las explicaciones más plausibles (Galun et al., 1982). Las me nores diferencias detectadas en el suelo L podrían estar relacionada con la alta capacidad de estos suelos finos de rete ner agua, lo que podría llevar a una re tención de agua por momentos superior en la zona sin CBs. Por el contrario, en suelos de texturas más gruesas, que tie nen menor capacidad de retención de agua, la presencia de las CB podría tener un efecto positivo (Rodriguez-Caballero et al., 2012). Coincidente con estos resul tados Chamizo et al. (2012), en el Cauti vo-España, encontraron que en suelos de textura fina la presencia de CBs tenía un impacto muy insignificante hasta inclu so negativo en el contenido de agua del suelo.
Conclusiones
Las CBs juegan un rol importante influ yendo en la tasa de infiltración y en el tiempo de persistencia del agua en los primeros centímetros del suelo. Sin em bargo, la influencia está determinada por el grupo funcional que domina en la CB y por la textura del suelo. En los ecosis temas secos donde el agua es el recurso que más limita la productividad, la con servación y optimización de los cortos y escasos pulsos de agua favorecidos por la presencia de CB se transforma en una ventaja fundamental para asegurar la ac tivación de procesos ecológicos que de otra forma permanecerían en latencia.

