











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCTION
The spider genus MelloinaBrignoli, 1985, currently included in the family Theraphosidae, is represented by three species: M. gracilis (Schenkel, 1953) and M. santuarioBertani, 2013 from Venezuela and M. rickwestiRaven, 1999 from Panamá (World Spider Catalog, 2022). Melloina gracilis is only known from males and M. rickwesti from females, and M. santuario is the only species of the genus with both sexes des cribed.
The genus is characterized by distinct maxi llary lobes (longer than in Theraphosidae, shor ter than in Paratropidinae); presence of thin claw tufts on the legs and female palp and eye tubercle normally elevated (Raven, 1985, 1999); tarsal scopulae reduced or absent and tarsi of legs with spines (Mori & Bertani, 2020). Studies on this genus have been scarce, and their phylo genetic position appears to be an important piece in determining the relationship of the families Theraphosidae, Barychelidae and Paratropididae. The morphological and molecular studies di ffer in their position (Raven, 1985; Goloboff, 1993; Goloboff, 1995; Raven, 1999; Bertani, 2013; Opatova et al., 2020; Mori & Bertani, 2020). The genus was originally described wi thin Theraphosidae by Schenkel (1953), under Melloa, and transferred to Paratropididae by Raven (1985). Brignoli (1985) proposed the name Melloina as a replacement for Melloa, preoccu pied by a genus of Opiliones. Recently, Mori & Bertani (2020) reinstated the genus in the family Theraphosidae. Their morphological analysis fo cused on the subfamily Trichopelmatinae, but also included representatives of Theraphosidae, Paratropidinae and Melloina. Although in Mori and Bertani’s analysis Melloina was recovered as closer to Holothele (Theraphosidae) than to the rest of the Paratropididae, the support for those relationships was weak, and the evidence tenuous. The familial position of Melloina thus remains far from settled.
During the course of a separate study on Paratropididae, examination of specimens de posited in the Museo Argentino de Ciencias Naturales “Bernardino Rivadavia” revealed a fe male specimen belonging to the genus Melloina. Several differences (fovea, cuspules, and sper mathecae) suggest that the specimen belongs to neither M. santuario nor M. rickwesti. Given the short geographical distance to the type locality of M. gracilis, the most conservative decision is to consider them as conspecific, and the specimen is therefore described here as the female of M. gracilis.
MATERIAL AND METHODS
The general format of the description follows Bertani (2013) with modifications. All measure ments are given in millimeters and were taken on the left side of the specimen. Total lengths were taken with chelicerae, and carapace lengths without chelicerae. Reference points for measu rements followed Coyle (1975). The specimens were analyzed in a MOTIC trinocular/SMZ-168 stereoscope and were measured and photogra phed with a Leica M205A stereomicroscope. The notation for leg spines follows Raven (1999); variation in the sides of a specimen (in number of spines, cuspules, or teeth) is indicated as two numbers or formulae separated by a slash (“/”).
The studied material is deposited in the Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”. The following abbreviations were used in the text: AME = anterior median eyes, ALE = anterior lateral eyes, PME = posterior median eyes, PLE = posterior lateral eyes, D = dorsal, P = prolateral, R = retrolateral, V =ventral, AP = apical, STC = superior tarsal claw, ITC = inferior tarsal claw. NHMB: Natural History Museum, Basel, Switzerland.
Our phylogenetic considerations are made taking into account the current reclassification made by Mori & Bertani (2020). In their phylo genetic analysis, they included representatives of the families Theraphosidae, Paratropididae and Barychelidae, in a matrix with 94 morphological characters scored across 66 species. Phylogenetic data matrix available in Morphobank (http://morphobank.org/permalink/?P4499).
We included the female described here in the matrix presented by in Mori and Bertani (2020) and explored changes in coding for some charac ters that seem problematic. Characters 0 and 1 (number of Maxillary and Labial cuspules) were treated as continuous in their analysis, but they represent some differences between very small and other differences between very large values. Thus, a change from (say) 5 to 10 maxillary cus pules is much less costly than a change from 300 to 315 (maximum possible value for this charac ter is 367), even when the first change amounts to a duplication of the number of maxillary cus pules, and the second only an increase in 5%. The mere rescaling of the characters to a 0-1 range (used by Mori & Bertani 2020) does not solve this problem; we instead transformed the values into logarithms and then scaled to 1, making diffe rences in low numbers more important (applying logarithms and rescaling so that the maximum of 367 cuspules correspond to a value of 1.0, the differences in the 5-10 versus 300-315 exam ple are respectively 0.117 and 0.008, so that the change 5-10 is much more “important” than the 300-315). Character 56 (Palp, legs I and II of females, scopula: (0) absent; (1) sparse; (2) dense; (3) spatulate), was treated as additive and Character 35 (soil encrusted cuticle: (0) lac king encrusted soil particles; (1) encrusted with soil) was recoded as 1 for all species of Melloina. Character 22 (labium shape: (0) quadrate; (1) tra pezoid; (2) ellipsoid) was coded as 2 for Melloina (Fig. 1C); Character 26 (book lung combs: (0) ab sent; (1) present) was coded to have a gain cost of 2 and lost 1. One may consider that many re peated identical originations of such an unusual structure, known in only some barychelids and nowhere else, is less likely than one or a few acquisitions and secondary losses. Characters 70 and 72 (tarsus III and tarsus IV: integral, crac ked or pseudosegmented) were merged into one (as they are not fully independent in both sexes). Note that none of these changes represent major modifications to the matrix presented by Mori & Bertani (2020), which highlights that any resolu tion of the relationships between theraphosids, barychelids and paratropidids must be conside red as very weakly supported.
A cladistic analysis was carried out with TNT version 1.5 (Goloboff, Farris & Nixon, 2008; Goloboff & Catalano, 2016), using parsimony as the optimality criterion. The tree searches were carried out under equal and implied weight (k = 12). The minimum scores were found 20 times, using the New Technology options. The values of combined Bremer support were estimated in the implied weight trees, by means of TBR swapping (which compared 1,877, 910 trees using subopti mal trees of value 0.967), displayed on the pruned strict consensus tree (Fig. 2). This measure ment combines values from the absolute Bremer (Bremer, 1994) and relative Bremer (Goloboff & Farris, 2001) in a single value, producing in simple cases evaluations that approximate the results of jackknifing (Farris et al., 1996).
SYSTEMATICS
Family Theraphosidae
Genus Melloina Brignoli, 1985.
Type species: Melloa gracilisSchenkel, 1953 (NHMB).
MelloaSchenkel 1953: 3 = GlabropelmaRaven 1985: 122 = MelloinaBrignoli 1985: 380.
Melloina gracilis (Schenkel, 1953)
(Fig. 1A-F)
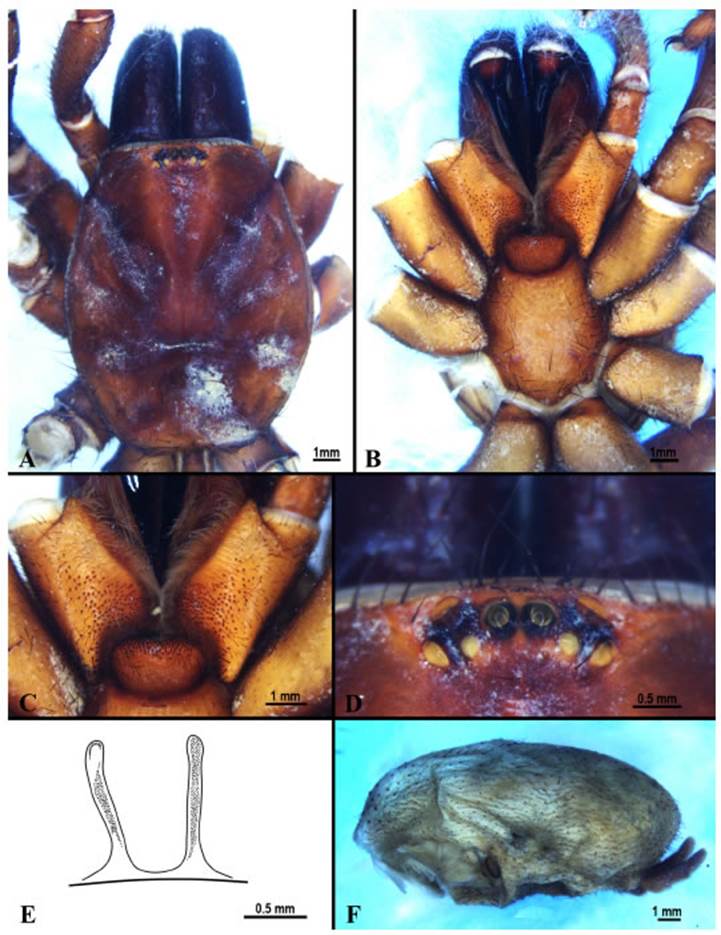
Fig. 1 A-F. Female of Melloina gracilis (Schenkel, 1953). A carapace, dorsal view; B sternum, ventral view; C labium and maxillae, ventral view; D ocular pattern, dorsal view; E spermathecae, F abdomen, lateral view.253 Goloboff-Szumik & Ríos-Tamayo: female of Melloina gracilis
Melloa gracilisSchenkel, 1953: male, by origi nal description (No examined). Melloina graci lisBrignoli, 1985 (replacement name for Melloa Schenkel, 1953); Glabropelma gracilisRaven, 1985; Melloina gracilis Raven, 1999; Bertani, 2013.
Key for identification of female Melloina
1. Fovea procurved ................. Melloina rickwesti
1’. Fovea straight or recurved ........................... 2
2. Maxillae with >120 cuspules............................ ...............................................Melloina gracilis
2’. Maxillae with <100 cuspules........................... ............................................Melloina santuario
Diagnosis. Females of M. gracilis can be dis tinguished from M. santuario and M. rickwesti by their higher number of maxillary cuspules (129/141 versus 82/90 and 60 respectively) and by their larger size (30 mm versus ~20 mm and 12 mm respectively). They can be differentiated from M. santuario by their higher number of labial cuspules (91 versus 77) similar to M. ric kwesti. Also, it can be differentiated from M. ric kwesti by the shape of their spermathecae (Fig. 1E) with the apical lobes less rounded; and their slightly recurved fovea (Fig. 1A) and from M. santuario by their longer spermathecae.
Material examined. Venezuela. Estado Falcón. San Luis. 1973. Philip Chapman. 1 ♀ (MACN-Ar 38247).
Description: Female (MACN-Ar 38247): to tal length 30.32, carapace length 11.24, width 9.0. Carapace (Fig. 1A). Glabrous; lines of thick bristles along the edges. Mid-length of cara pace posteriorly with ~35 bristles. Fovea short, slightly recurved, 1.94 length. Clypeus absent. Eyes (Fig. 1D): tubercle low, well defined; ante rior edge is carapace edge. Anterior row slightly procurved, posterior row recurved. Ocular group, length 0.73, width 2.02, with 3 bristles anterior of AME and 6 between posterior eyes. Ocular measurements: AME 0.23, ALE 0.36, PME 0.25, PLE 0.35; AME-ALE 0.32, PME-PLE 0.19, AME-PME 0.22, ALE-PLE 0.21, AME-AME 0.22, PME-PME 0.81. Labium (Fig. 1C): width 2.23, length 1.61; broad, rounded, with an in vagination in the anterior part, ca. 91 cuspules. Maxillae (Fig. 1D): anterior length 3.21, poste rior length 4.21, width 2.03, with ca. 129/141 uniformly distributed cuspules. Heel slightly produced; anterior lobe distinct. Chelicerae: dor sally with a wide row of dark bristles; promargin with long row of 17 large teeth with ca. 35 denti cles. Sternum (Fig. 1B): 5.43 length, 4.27 width; longer than wide; strong setae around margins shorter. Cuticle: clean without soil encrusted. Sigilla small, oval, about a diameter of distance from the edge. Abdomen (Fig. 1F): 14.30 length. Spinnerets: posterior median: length 0.78; poste rior lateral with basal: medial: apical articles of lengths 1.41:0.85:1.16. Booklung oval with scle rotized openings.
Legs: Glabrous; covered only with bristles and spines. Cuticle lightly soil encrusted. Only tarsi IV cracked. Lengths of legs and pedipalp (femora, patellae, tibiae, metatarsi, tarsi, total): I: 9.35, 5.75, 8.62, 7.60, 3.94, 35.24. II: 8.28, 4.73, 6.99, 6.57, 3.73, 30.30. III: 7.75, 4.00, 6.00, 7.78, 3.52, 29.26. IV: 10.51, 4.50, 9.35, 11.85, 4.56, 40.77. pedipalp: 5.91, 3.90, 5.04, -, 3.99, 18.8. Chaetotaxy: leg I, fe 0; pa 0; ti v3 ap; me v10; ta v6+6 in two straight rows; leg II, fe 0; pa 0; ti p2, v5, r2; me p5, v7, r2, d2; ta v6+8 in two straight rows; leg III, fe 0; pa 0; ti p3, v3 r3; me p6, r3, v10; ta v10+8 in two straight rows; leg IV, fe 0; pa 0; ti p2, r2, v7; me p3, d8, v7, ta v8+8 in two straight rows; palp fe 0; pa 0; ti v3 ap; ta v12. Claws: ITC absent from all legs; STC with 3 small teeth on medial keel of leg I, II, III, 2 on IV. Palp with single claw without tooth. Claw tufts weak but distinct on all legs. Tarsi IV cracked. All legs with translucent spatulate hairs interspersed with the trichobothria. Trichobothria: palp with a wide band of thin bristles interspersed among trichobothria. Tibiae (in two rows): palp 8/8, I 10/9, II 10/10, III 9/8, IV 10/9. Metatarsi: I 10, II 14, III-IV 20. Tarsi: Filiform: palp 15, I 5/5, II 9/10, III 10/10, IV 12/14. Clavate: palp 18, I 7/6, II 8/9, III 7/8, IV 10/12. Spermathecae (Fig. 1E): two long undivided lobes. Color (in alcohol): car apace, and legs reddish brown, chelicerae dark brown, abdomen light brown.
Other material examined. None
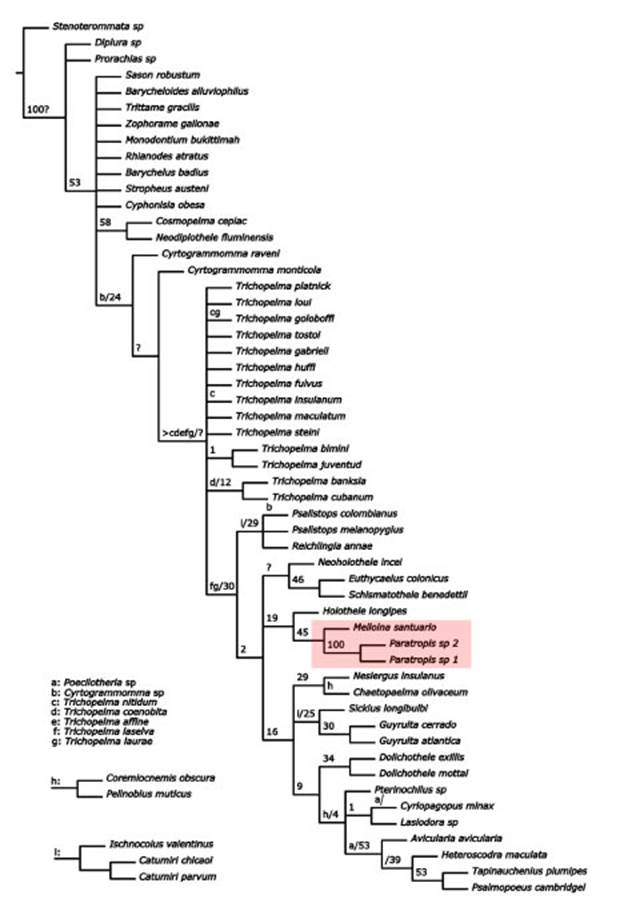
Fig. 2 Strict consensus between both analyses when the most unstable taxa were excluded. Pink box shows the recovery of the genera Melloina and Paratropis together. Branch letters represent the position of each taxon or group of taxa. Numbers above branches represent support values of combined Bremer support.
DISCUSSION
The genus Melloina comprises three species, described exclusively from the Neotropical re gion, and distributed in Panama and Venezuela (World Spider Catalog, 2022). In the pres ent work, a female of Melloina is recorded for Venezuela, geographically close to the male of M. gracilis (around 100 km). Both specimens (male and female) are present in similar biogeograph ic sites with the presence of evergreen forests (Jonathan Liria, per. comm). This new female can be placed in the genus Melloina by the shape of their spermathecae (simple and long), a very styl ized body with the cuticle lightly soil-encrusted, elongated glabrous legs covered only with thin erect bristles and booklung apertures oval and sclerotised. This female differs from other spe cies by the shape of their fovea and morphology of their spermathecae. The number of maxillary cuspules was used as a diagnostic character by Bertani (2013); the examination of this structure in the newly described female shows a higher number than that of the other species.
The phylogenetic position of the genus Melloina has long been critical for understand ing the relationships of the family Paratropididae (Raven, 1985; Hedin & Bond, 2006). Raven (1985) proposed Melloina to be more closely related to Paratropidinae than to the Theraphosidae, and if this is correct, supports the view that paratropi dines are very autapomorphic Theraphosoidina, with Melloina acting as an intermediate link. If Melloina is not more closely related to Paratropidinae, then paratropidines may well be only distantly related to Theraphosoidina, even sharing some potential synapomorphies with Raven’s (1985) Rastelloidina; this would explain the instability of Paratropidinae in the analyses of Hedin & Bond (2006) or Opatova et al. (2020), which included no species of Melloina.
The present description of the female of M. gracilis allowed us to include this new specimen in the morphological matrix published by Mori & Bertani (2020), and explore changes in coding for some characters. Two analyses were made, un der equal (retaining 10 trees, best score 526.730) and implied weights (retaining 9 trees, best score 19.10519). Figure 2 shows the strict consensus between both analyses when the most unstable taxa are excluded. All phylogenetic analyses made (under equal and implied weight), recovered the Theraphosoidina clade, but -in contrast to the results of Mori & Bertani (2020)-the genus Melloina grouped with Paratropis (and inside of “Theraphosidae”; see Fig. 2). We do not intend to propose any taxonomic changes, but aimed only to comment that only minor changes to the matrix presented by Mori & Bertani (2020) suffice to tilt the results toward the more tradi tional hypothesis of grouping Melloina and the Paratropidinae, as proposed by Raven (1985) and accepted without dispute (e.g., Goloboff, 1993) until Hedin & Bond’s (2006) analysis. Clearly, future studies must consider more species of the family Paratropididae, as well as Melloina and potentially related genera of Theraphosidae.

