










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo
versión impresa ISSN 1853-8665versión On-line ISSN 1853-8665
Rev. Fac. Cienc. Agrar., Univ. Nac. Cuyo vol.50 no.2 Mendoza dic. 2018
ORIGINAL ARTICLE
Influence of interannual rainfall variability on the interception process in a continental mediterranean climate
Influencia de la variabilidad interanual de la lluvia sobre el proceso de interceptación en un clima mediterráneo continental
María Fátima Moreno-Pérez1, Rafael Pérez-Arellano1, José Roldán-Cañas1
1Hydraulic Engineering, Department of Agronomy. University of Córdoba, Campus Rabanales. 14071. Córdoba. Spain. mfatima@uco.es
Originales: Recepción: 17/02/2017 - Aceptación: 20/04/2017
ABSTRACT
The interception of rainfall by vegetation greatly affects the hydrology of a basin. The main objective of this paper is to evaluate the effect of annual rainfall variability on the interception in isolated individuals of two of the most common types of vegetation, Pinus pinea and Cistus ladanifer, in a Mediterranean watershed. For this, experimental data were taken for four years and the Rutter interception model was used to calculate the canopy drying time, while Rutter and Klaassen methodologies were used to determine the storage capacity of the canopy. On the whole, 29.6% of rainfall was intercepted for pine, and 17.7% for rockrose. Stemflow accounted for 0.3% for pine and 17% for rockrose, while 70.1% of the rainfall fell as throughfall for pine and 65.3% for rockrose. However, in dry years, the interception percentage almost doubled that of wet years. An average canopy drying time of 9.2 hours was established for pine, and 4.4 hours for rockrose. Approximately equal results of canopy storage capacity were obtained from both methods (1.8 mm for pine and 0.7 mm for rockrose). Both species registered higher values of canopy storage capacity in dry years than in wet years.
Keywords: Pinus pinea; Cistus ladanifer; Southern Spain; Canopy reservoir capacity; Canopy drying time
RESUMEN
La interceptación de la lluvia por la vegetación afecta sobremanera la hidrología de una cuenca. El objeto de este trabajo es determinar el efecto de la variabilidad anual de la lluvia sobre la interceptación en individuos aislados de Pinus pinea y Cistus ladanifer, en una cuenca mediterránea. Para ello, se tomaron datos experimentales durante cuatro años y se usó el modelo de interceptación de Rutter para calcular el tiempo de secado del dosel, en tanto que para determinar la capacidad de almacenamiento del dosel se consideraron las metodologías de Rutter y Klaassen. En conjunto, el 29,6% de la lluvia fue interceptada por pino y 17,7% por jara. El 0,3% en pino y el 17% en jara ocurrió como escorrentía cortical, y el 70,1% fue trascolación en pino y el 65,3% en jara. Sin embargo, en años secos, el porcentaje de interceptación casi duplicó el de años húmedos. El tiempo de secado medio en pino fue 9,2 horas y en jara 4,4. Aproximadamente, se obtuvieron iguales resultados de la capacidad de almacenamiento del dosel usando ambos métodos (1,8 mm en pino y 0,7 en jara), registrándose también valores más altos en años secos.
Palabras clave: Pinus pinea; Cistus ladanifer; Sur de España; Capacidad de almacenamiento del dosel; Tiempo de secado del dosel
INTRODUCTION
The partitioning of rainfall affects the water balance both locally and at basin scale, and accordingly the structure of the vegetation cover significantly affects the hydrology of a watershed. During and after a rainfall event, the vegetation temporarily stores rainwater on the canopy, which subsequently evaporates (interception), resulting in a reduction in the water available for infiltration (38). Rainfall that is not intercepted, falls as throughfall, affecting the surface layers of the soil and the rate of deep percolation below the root zone in a discontinuous forest cover, while stemflow can reach the deeper layers of soil and make water directly available to the roots. Some studies in the Mediterranean basins indicate that forest cover leads to a significant reduction in average annual measured flows, and so from a hydrological perspective, vegetation canopies exert significant control by modifying both evaporation and precipitation redistribution (4). It is demonstrated that interception losses represent a significant component of the water balance in forest areas (24). These losses can vary from 9% of incident rainfall in the Amazon rainforest (26) to almost 50% in coniferous forests in areas of Mediterranean mountainous climate (25).
The most common field method of determining interception is to measure incident precipitation, throughfall and stemflow. Interception is then the difference between precipitation and the sum of throughfall and stemflow. In general, trees have a higher relative throughfall (the ratio of throughfall to gross rainfall) and a lower relative stemflow than shrubs (26, 39).
Whatever the method used to obtain the canopy storage parameters for modeling interception, there is no standard protocol for measuring incident rainfall, throughfall and stemflow in interception studies (21). This is due, on the one hand, to the variety of study objectives and diversity in vegetation coverages studied (22) and, on the other hand, to the fact that sampling systems are not well adapted to the heterogeneity of the forests studied (40). The accuracy of measurement of these variables can give rise to negative interception values or, at least, errors in the interpretation of the measurements obtained (1, 8).
However, the process of interception is influenced by two main factors: the climate conditions and the structure of the species of trees or shrubs (35). In literature, no consensus exits on whether climate or vegetation is the dominant factor. For instance, (3) compared stemflow from Pinus sylvestris with five tree species at three sites in northern Britain and found that it was influenced by stand characteristics more than by weather conditions. On the other hand, (12) showed with a modeling study that the rainfall distribution is most dominant for the interception amount.
In Mediterranean forest where canopy cover is discontinuous, the wide variability in the relationships between rainfall and interception losses creates greater complexity (6, 15). In areas of dehesas pasture, such as our study area, where the forest is composed of isolated trees interspersed with shrubs, this complexity is even greater (5).
Canopy water storage capacity is a key factor determining the quantity of precipitation intercepted (13, 16). When rainwater that falls on the canopy exceeds the maximum holding capacity water begins to fall to the ground along with the water that manages to pass directly through the canopy without being intercepted (i.e., free throughfall). These parameters, the canopy water storage capacity (S) and the free throughfall coefficient (p), are related to climatic conditions during rainfall and to the canopy structure (26).
In coniferous forests, there is a close relationship between interception and the amount of water stored in the canopy (20). Moreover, in shrubs the water storage capacity can even at times be comparable to that of tree communities and, despite their short height, in arid and semi-arid environments water loss by evaporation in rainy periods may be similar to losses in more temperate forests (9).
Although several experiments have been carried out in areas generally referred to as Mediterranean climates, few have focused on the subtype and none to the best knowledge on a dehesa area, while dehesa's are a common ecosystem or land use type in southwestern Iberian Peninsula. The present study was conducted in isolated individuals of the most common types of vegetation in the Mediterranean "El Cabril" watershed, Pinus pinea and Cistus ladanifer, with objectives: (i) to estimate the impact of the annual rainfall variability on the process of interception; (ii) to determinate of the minimum number of hours required for all water stored in the canopy to evaporate and (iii) to calculate the amount of precipitation needed to reach saturation point in both species. While many studies in Mediterranean climates are of short duration this study encompasses a study period of four-year, which allows us to obtain more consistent results than those typically found in literature.
STUDY SITE AND MATERIALS
Site description
The experimental plots are located on a forest area known as "El Cabril" in the town of Hornachuelos, which is in the west of the province of Córdoba, Spain. "El Cabril" has an area of 11.26 km2 and its geographic coordinates are 38°4' N and 5°25' W (figure 1).
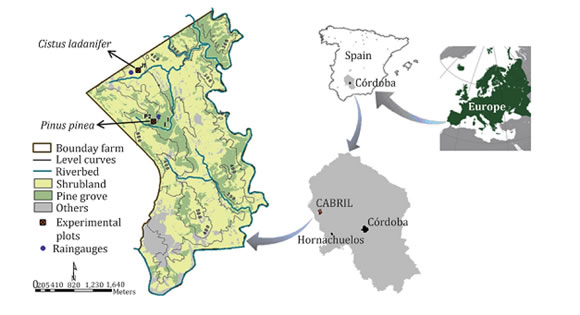
Figure 1. Location of the forest area of the "El Cabril".
Figura 1. Ubicación de la zona forestal de "El Cabril".
The climate of the study area is influenced by its two main geographical features: its elevation (500-700 m a. s. l.) and its relative continentality. It is characterized by a semi-arid continental Mediterranean climate, featuring hot, dry summers and moderately cold winters.
The average annual temperature is 16°C, with the maximum ranging between 37 and 41°C, and the minimum between -1 and 3°C. The average annual rainfall (1990-2013) is 506 mm/year with large variations (CV = 0.49), December being the wettest month (average 73 mm/month) and July and August the driest months (average 4 mm/month).
The rainfall regime is one of the most characteristic climatic aspects, with the majority of precipitation occurring from October to May.
The reason for studying isolated individuals is the frequent existence of sparse vegetation in the basin. The plant communities are dominated by Mediterranean sclerophyllous scrub (6.1 km2) with extensive thickets of rockrose (Cistus ladanifer) accompanied by retama (Retama sphaerocarpa), mastic (Pistacia lentiscus), flax-leaved daphne (Daphne gnidium), lavender (Lavandula stoechas) and rosemary (Rosmarinus officinalis). A large area at the basin rivers’ headwaters (2.6 km2) is covered by plantations of pine (Pinus pinea), planted in the early 1970s.
The relief of the study area is fairly steep, with an average slope of 30%. Other species such as eucalyptus and quercus are also present in the basin, but they cover a smaller area. The soils are sandy loam texture and relatively shallow with a depth not exceeding 30 cm in most of the basin (29).
Materials
Three experimental plots were established to determine throughfall (T f) and stemflow (S f), two of Pinus pinea and one of Cistus ladanifer. In each plot, there is only one individual of the corresponding species. The study was conducted over a four-year period from October 2010 to June 2014 in the case of the first pine plot and the rockrose plot. For the second pine plot, experimental data is available from September 2013 until June 2014. The characteristics of these plots are shown in table 1.
Table 1. Characteristics of experimental plots.
Tabla 1. Características de las parcelas experimentales.
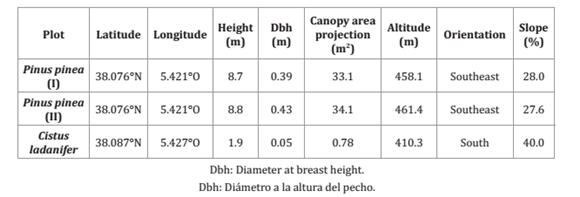
The second individual of pine has been chosen with similar dasometric characteristics to the first in order to eliminate the influence of geometry on the results obtained. To do this, it was measured the same variables that other authors chose to quantify the growth of species due to management strategies (i.e. Donoso et al., 2016) or to establish the value of individuals of commercial or ornamental interest (i.e. Ponce-Donoso and Vallejos-Barra, 2016). These variables, among others, are the height of the tree, the canopy area projection or the diameter at breast height (table 1).
Two tipping bucket raingauges were installed to record the incident precipitation with an accuracy of 0.2 mm (Eijkelkamp). Precipitation data are recorded every 10 minutes on a datalogger (e+® RAIN).
The raingauges are located in two open areas, approximately 20 meters away from the pine plots and the rockrose plot. Also, there is a complete meteorological station at El Cabril watershed.
In both pine plots, a permeable fence (two meters high) was installed to protect the instrumentation inside, delimiting an area of 28.27 m2, in both cases.
In the pine plots the volume of throughfall (T f) is measured by eight standard manual raingauges of 9.3 cm in diameter located under the pines according to cardinal directions and forming two concentric rings.
In addition, six troughs consisting of 20 cm wide and 2 m long metal guttering were installed radially just below the canopy within the research plots. Pérez Arellano et al. (2016), found that similar statistically results were obtained with any of these methods.
The guttering was installed at a slope of about 10% to encourage the draining of the collectors. Stemflow (S f) is measured by an open, flexible tube sealed with silicone around the trunk of the trees, and channeling the water collected to a 12-liter tank.
The guttering and the raingauges were cleaned at least once a month but on occasions they still got blocked.
Furthermore, it was also measured T f and S f on a rockrose plant that had representative characteristics for Cistus ladanifer, by means of an adaptation of the so-called "interception flow collection box" described by Belmonte and Romero (1998). This device consists of a 0.45 m2 impermeable square surface surrounded by an L-shaped perimeter that ensures that the throughfall does not overflow, and that it reaches the drainage point located in one of the corners (figure 2).
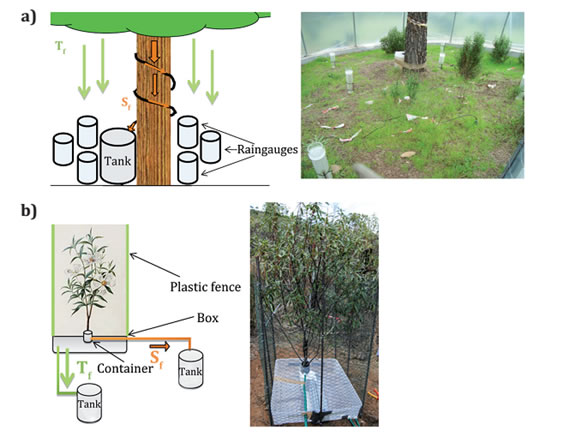
Figure 2. Experimental plots for Pinus pinea (a) and Cistus ladanifer (b).
Figura 2. Parcelas experimentales en Pinus pinea (a) y Cistus ladanifer (b)
All throughfall water is collected and channeled to a 25-liter tank.
The plot is also waterproofed laterally to prevent the entry of lateral rain. To measure stemflow, a container was placed around the base of the trunk to collect the water flowing through the branches to the trunk. This water is conveyed to a 12-liter collection tank.
The gaskets of the "interception box", the drain holes and cortical runoff collector were sealed with silicone.
Measurements were taken when the rainfall events had ended and after a sufficient period of time for the canopy storage to empty and for throughfall and stemflow to be fully collected (usually one day after the end of the rain).
Methods
Effect of annual rainfall variability on the process of interception
In this study, the amount of intercepted precipitation has been determined indirectly by taking the difference between the incident rainfall and the measure precipitation passing through the vegetation canopy, which is divided into throughfall and stemflow:
I = P − Tf − S f (1)
where:
I = interception (L)
P = incident precipitation (L)
Tf = throughfall (L)
Sf is stemflow (L).
During study period, it was calculated the water balance per precipitation event and we obtained information about T f and S f for Pinus pinea and Cistus ladanifer. The influence of precipitation annual variability on the interception process can be studied because it was disposed of four years of data and a very high number of rainfall events.
Canopy Drying time
A precipitation event is defined as one or multiple showers with at least enough time between the events to dry the canopy, where a shower is a single-rainfall occurrence. In this case, it was applied the Rutter model of interception (35) to estimate canopy drying time, using hourly climatic data from the El Cabril weather station.
The model is based on a calculation of the water balance in the canopy and trunks, and uses incident precipitation and evaporation in the canopy and the trunk to estimate throughfall, stemflow and interception. Wet canopy evaporation can be directly calculated from the Penman-Monteith potential evaporation equation (28).
Pereira et al. (2016), have proposed a wet bulb to estimate mean evaporation rate as an alternative to the Penman Monteith equation when the forest canopy is fully ventilated. The canopy drying time (CDT, h) was obtained by calculating the amount of time it takes for the canopy to dry completely after the end of the rainfall event ("rain shower"); given that we have hourly precipitation and potential evaporation data, we know when the rain stops and how many hours it takes until, according to the model, there is no longer any water stored in the canopy.
Canopy storage capacity
According to Leyton et al. (1967), canopy storage capacity (S, mm) can be defined as the amount of precipitation needed to reach saturation point and it can be determined using the graphic method whereby incident precipitation P is plotted against T f for individual events. Due to the fact that the field readings could comprise a number of rainfall events and that there was no continuous record of throughfall, it was selected only the events with one or two showers to calculate S.
In the case of an event with two showers, the total amount of intercepted water was divided in proportion to the amount of incident precipitation collected in each shower.
The scatter plot demonstrates a two-step relationship between incident precipitation and throughfall (34).
The first step was dictated by canopy storage and the second step occurs after saturation of the canopy.
The intersection point between the regression line fitted to the first-step data and the upper envelope line of the second-step data as proposed by Gash and Morton (1978) and Lloyd et al. (1988), represents the event size in which the amount of incident precipitation is sufficient to saturate the canopy (P s ). Finally, the value of S is obtained from the intercept of the upper envelope on the throughfall axis (in absolute value).
Specifically, when the amount of the incident precipitation in an event was less than P s , the incident precipitation was mostly intercepted by the canopy and the amount reaching the ground, does so as free throughfall; therefore, the regression line slope between Tf and P indicates the proportion of incident precipitation reaching the ground as free throughfall (18). So, for the first step, the coefficient of free throughfall (C), defined as the proportion of incident precipitation reaching the ground as free throughfall, can be calculated as the ratio between T f and P s , and the canopy storage capacity can be obtained as suggested (34), by applying equation 2.
S = (1 - C) Ps (2)
Following the methodology proposed by Huanhua et al. (2013), which defines the canopy storage capacity (S, mm) as (1 − ) s the maximum possible water storage after a quick drain has stopped, and calculates S using a scatter plot of measured interception (I) versus P, linear regression of the type I = α·P + β is fitted to the second-step data, taking α as the ratio of precipitation that evaporates from the canopy and β as the canopy storage capacity (S).
RESULTS
Effect of annual rainfall variability on the process of interception
It should be noted that in the first and third year precipitation was 60% and 25% higher, respectively, than the average of the previous 20 years, while in the second and fourth year precipitation was 44% and 20%, respectively, below the previous average (table 2, page 147).
Table 2. Precipitation and throughfall, stemflow and interception losses for period 2010-2014.
Tabla 2. Precipitación y pérdidas por trascolación, escorrentía cortical e interceptación durante el período 2010-2014.
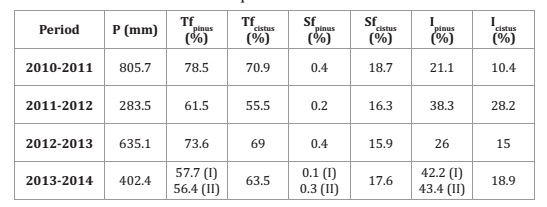
These data show the interannual variability of rainfall over the years.
It was recorded 64 rainfall events for Pinus pinea and 53 for Cistus ladanifer. The rain showers had an average intensity of 2.81 mm/hour.
The maximum intensity recorded at 10 and 60 minutes were 13.8 mm and 28.6 mm, respectively.
In Pinus pinea the cumulative amount of precipitation measured after 64 events was 2083.6 mm (CV = 0.96), with a high variability (ranging from 1.2 mm to 180.6 mm), being the volume of water intercepted 616.6 mm (29.6%).
In the Cistus ladanifer plot, only 53 events could be included in the study due to different problems when taking the samplers (such as filling up the water collection tanks and installation errors). In this case, the accumulated rainfall was 1289.4 mm, with a lower coefficient of variation (CV = 0.74), and an interception amount of 228.6 mm (17.7%).
For Cistus ladanifer the average throughfall was 65.3% and the average stemflow was 17%, whereas in Pinus pinea these values were 70.1% and 0.3%, respectively. Table 2, shows the results broken down by year. The year 2013-2014 displays results for both the first and second pine.
Canopy drying time
For the pine, the Rutter model identified a total of 278 rainfall events ("rain showers") with an average drying time of 9.2 hours. The canopy drying time for rockrose is 4.4 hours, and 374 rainfall events were identified in this case. Erring on the side of caution, it was established a minimum period of 10 hours to evaporate all the water stored in the canopy for Pinus pinea and 5 hours for Cistus ladanifer.
Based on the CDT values obtained, it can be assumed that the methodology followed for taking measurements of throughfall and stemflow is appropriate. All water from throughfall and stemflow had already been collected by the different tanks the day immediately after rainfall.
Canopy storage capacity
Figure 3 (page 148) shows the relationship between incident precipitation and throughfall of 38 showers at the first pine plot and of 35 showers at the rockrose plot (see section 3.3).
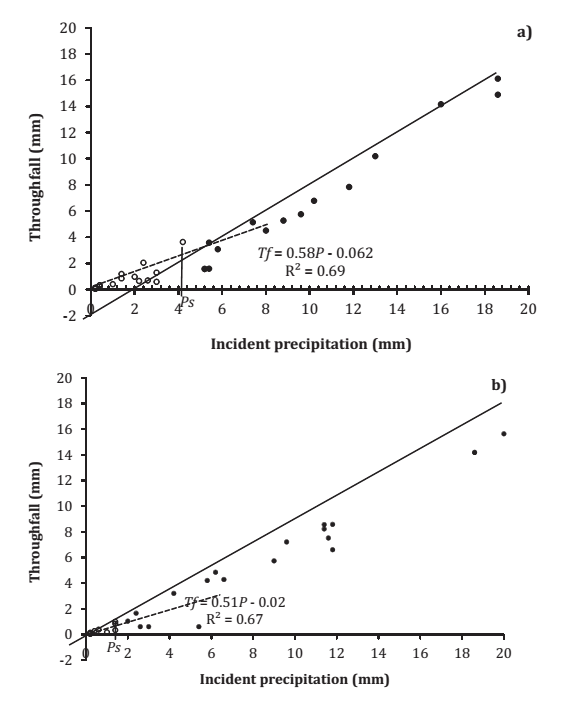
Figure 3. Scatter plot of throughfall against incident precipitation (Tf > Ps(○); Tf <Ps(•)) for Pinus pinea I (a) and Cistus ladanifer (b). The regression line fitted to the first-step data and the upper envelope line of the second-step data is depicted with a discontinuous and continuous line, respectively (Canopy storage capacity).
Figura 3. Representación de la trascolación frente a la precipitación incidente (Tf > Ps(○); Tf <Ps(•)) para Pinus pinea 1 (a) y Cistus ladanifer (b). La línea de regresión ajustada a los datos del primer escalón y la línea envolvente superior de los datos del segundo escalón se representan mediante una línea discontinua y continua, respectivamente (Capacidad de almacenamiento del dosel).
Measurements from the second pine plot are not shown as there are insufficient data to provide significant results. In these cases, P s and S obtained graphically are 4.2 mm and 1.8 mm for Pinus pinea, and 1.4 mm and 0.6 mm for Cistus ladanifer, respectively.
The value of C obtained as the values of slopes of the regression equations between throughfall and incident precipitation for the first step in Pinus pinea and Cistus ladanifer are 0.58 and 0.51, respectively, as indicated in section 3.3. Applying equation 2 (page 146), gives S values of 1.76 mm for Pinus pinea, and 0.69 mm for Cistus ladanifer. Both values are in close agreement with those obtained graphically (1.8 mm for Pinus pinea and 0.6 mm for Cistus ladanifer).
These results were compared following the methodology proposed by Klaassen et al. (1998), figure 4 (page 149), that yielded values of 1.81 mm for Pinus pinea, and 0.74 mm for Cistus ladanifer, similar to those obtained above.

Figure 4. Scatter plots of interception against incident precipitation for Pinus pinea I (a) and Cistus ladanifer (b).
Figura 4. Representación de la interceptación frente a la precipitación incidente para Pinus pinea (a) y Cistus ladanifer (b).
The value of S varies between the two types of years being greater in dry years (2.07 in pine and 1.09 in rockrose) than in wet years (1.71 in pine and 0.61 in rockrose). This would seem logical since in dry years the wet canopy evaporation increases and, consequently, the storage capacity of the plant also increases.
DISCUSSION
Effect of annual rainfall variability on the process of interception
The results show that the canopy interception differs per vegetation type and annual precipitation (table 2, page 147).
The interception rate is higher in dry years for both species, and is nearly double that of wet years. This can be explained with reference to the canopy storage capacity, which decreases in rainy years and is exceeded more easily, as will be made clear in section 4.3. Accordingly, throughfall is lower in dry years than in wet years.
The results from both the first and second pine are very similar, which might initially suggest that a single year is enough to record conclusive measurements. Nevertheless, table 2 (page 147), shows widely varying results between dry and wet years, thus indicating that study periods of more than a year are advisable.
There is a significant difference between species in terms of the redistribution of precipitation. Stemflow is negligible in Pinus pinea, but substantial in Cistus ladanifer. This is largely due to differences in plant structures.
In rockrose, the relative stemflow is very variable, which may be an indication of the effect of the resin produced by these shrubs, that acts as a repellent against the first drops of rain.
García-Estringana et al. (2010), showed that the impermeable wax, ladane, secreted by Cistus ladanifer acts as a water repellent and diminishes the retention of intercepted rainfall. Plant surfaces are rendered water repellent by three-dimensional microstructures of the epidermal cells and hydrophobic wax crystals (17).
The amount of throughfall for Cistus ladanifer is lower than that obtained by Simões et al. (2009), in Évora (southern Portugal) for the same specie (79% T f) during two years, but higher than Cistus salviifolius (61% T f). Such variation may be mostly a result of differences in canopy structure; Cistus ladanifer is characterized by sparse foliage clumps distributed vertically and with larger gaps in the canopies.
For Pinus pinea the results are consistent with those obtained in two studies carried out in a Mediterranean climate: Llorens et al. (1997), for Pinus sylvestris (24% I) in Barcelona, Spain; and Ibrahim et al. (1982), for Pinus pinea (27% I) in Petit Saint-Jean, France.
However, Ghimire et al. (2012), in their study of a forest of Pinus roxburghii located in Nepal recorded slightly different proportions of throughfall, stemflow and interception, at 83%, 0.5% and 16.5% of precipitation, respectively. This demonstrates the impact of climate on the interception process in similarly-structured species.
Levia Frost (2003), linked the large amounts of stemflow in rockrose, in all cases and even with low-intensity rains, to the plant having a cone-or funnel-shaped structure. Cistus ladanifer features just such a structure, with vertically-oriented branches at an insertion angle of more than 45° to the horizontal, thus favoring the flow of water from the branches to the trunk. However, in pine, cortical runoff is lower in drier years because the intensity and duration of rainfall decreases. Once again, the limited data available for the second pine means that results are of little relevance.
Canopy drying time
Canopy drying time generally varies depending on evaporation rate after the rain (10, 35). There is no standard criterion for the duration of the drying period, as it depends largely on the climate and natural processes affecting precipitation. Some authors have set this time at 4 hours (33), while others estimate 12 hours (3).
This results show that canopy drying time varies from specie to specie, being more than double for Pinus pinea that for Cistus ladanifer, which makes sense given the difference between canopy structure of both species, that is much greater for Pinus pinea.
Canopy storage capacity
The results are consistent with experimental values for interception measured in both plant species. It is indeed the case that a greater amount of incident precipitation is required to saturate the pine canopy compared to the rockrose. Accordingly, the canopy storage capacity is greater for pine.
For small quantities of incident precipitation-too small to saturate the canopy-the slope of the line-of-fit is practically the same in pine and in rockrose (figure 3, page 148), indicating equal throughfall increases in both species in response to equal increases in precipitation. Under these conditions, the only throughfall is free throughfall. Moreover, the slope in each case should show the relationship between throughfall and the incident precipitation, or the coefficient of free throughfall, with a zero intercept. Values different from zero obtained from said intercept in this study, although relatively small, are due to expected experimental measurement errors.
Thus it can be seen that either method can be used to determine such information, although it is advisable to use both in order to confirm the validity of the results, particularly with low values for the coefficient of determination R2, as was the case for Cistus ladanifer (R2=0.18) when the Klaasen methodology was used (figure 4, page 149).
These values are similar to those obtained by Saxena (1986), for a pine forest in Uppsala (S = 2.1 mm, C = 0.49 and Ps = 4.1 mm).
For Cistus ladanifer, we find a higher value than García-Estringana et al. (2010), who obtained an average S value of 0.54 mm using eight individuals in a laboratory setting. The different values obtained by different authors may be due, among other reasons, to the use of short data series that fail to reflect the annual rainfall variability.
The difference between dry and wet years in the value of seems logical since the maximum evaporation rate from wet/saturated forest canopies increases in dry years and, consequently, the storage capacity of the plant increases.
CONCLUSIONS
This research show that the canopy interception differs per vegetation type and annual precipitation. Notable differences in interception values were found when comparing dry and wet years. The interception rate is higher in dry years for both species, and is nearly double that of wet years. Accordingly, throughfall is lower in dry years than in wet years. Then, to reach significant conclusions in interception studies, the use of data from periods of greater than one year is therefore recommended.
Canopy drying time for each species following a rain event is quite different given the morphology and plant architecture differences between both species, being more than double for Pinus pinea that for Cistus ladanifer.
A greater amount of incident precipitation is required to saturate the pine canopy compared to the rockrose.
Accordingly, the canopy storage capacity is greater for pine.
The value of canopy storage capacity varies between the two types of years being greater in dry years than in wet years, in both species, because the canopy storage capacity decreases in rainy years and is exceeded more easily.
1. Allen, S. T.; Keim, R. F.; McDonnell, J. J. 2015. Spatial patterns of throughfall isotopic composition at the event and seasonal timescales. J. Hydrol. 522: 58-66. [ Links ]
2. Belmonte, F.; Romero, A. 1998. A simple technique for measuring rainfall interception by small shrub: Interception flow collection box. Hydrol. Process. 12: 471-481. [ Links ]
3. Cape, J. N.; Brown, A. H. F.; Robertson, S. M. C.; Howson, G.; Paterson, I. S. 1991. Interspecies comparison of throughfall and stemflow at three sites in northern Britain. Forest Ecol. Manag. 46: 165-177. [ Links ]
4. Cognard-Plancq, A. L.; Marc, V.; Didon-Lescot, J. F.; Normand, M. 2001. The role of forest cover on streamflow down sub- Mediterranean mountain watersheds: a modelling approach. J. Hydrol. 254: 229-243. [ Links ]
5. Cubera, E.; Moreno, G. 2007. Effect of single Quercus ilex trees upon spatial and seasonal changes in soil water content in dehesas of central western Spain. Ann. For. Sci. 64: 355-364. [ Links ]
6. David, T. S.; Gash, J. H. C.; Valente, F.; Pereira, J. S.; Ferreira, M. I.; David, J. S. 2006. Rainfall interception by an isolated evergreen oak tree in a Mediterranean savannah. Hydrol. Process. 20: 2713-2726. [ Links ]
7. Donoso, S.; Peña-Rojas, K.; Galdames, E.; Pacheco, C.; Espinoza, C.; Durán, S.; Gangas, R. 2016. Evaluación de la aplicación de biosólidos en plantaciones de Eucalyptus globulus, en Chile central. Revista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo. Mendoza. Argentina. 48(2): 107-119. [ Links ]
8. Frischbier, N., Wagner, S. 2015. Detection, quantification and modelling of small-scale lateral translocation of throughfall in tree crowns of European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.). J. Hydrol. 522: 228-238 [ Links ]
9. García-Estringana, P.; Alonso-Blázquez, N.; Marques, M. J.; Bienes, R.; Alegre, J.; 2010. Direct and indirect effects of Mediterranean vegetation on runoff and soil loss. Eur. J. Soil Sci. 61: 174-185. [ Links ]
10. Gash, J. H. C. 1979. An analytical model of rainfall interception by forests. Q. J. R. Meteorol. Soc. 105: 43-55. [ Links ]
11. Gash, J. H. C.; Morton, A. J. 1978. An application of the Rutter model to the estimation of the interception loss from the Thetford forest. J. Hydrol. 38: 89-105. [ Links ]
12. Gerrits, A. M. J.; Savenije, H. H. G.; Veling, E. J. M.; Pfister, L. 2009. Analytical derivation of the Budyko curve based on rainfall characteristics and a simple evaporation model. Water Resources Research. p. 45. [ Links ]
13. Gerrits, A. M. J.; Pfister, L.; Savenije, H. H. G. 2010. Spatial and temporal variability of canopy and forest floor interception in a beech forest. Hydrol. Process. 24: 3011-3025. [ Links ]
14. Ghimire, C. P.; Bruijnzeel, L. A.; Lubczynski, M. W.; Bonell, M. 2012. Rainfall interception by natural and planted forests in the Middle Mountains of Central Nepal. J. Hydrol. 475: 270-280. [ Links ]
15. Guevara-Escobar, A.; Gonzalez-Sosa, E.; Veliz-Chavez, C.; Ventura-Ramos, E.; Ramos-Salinas, M. 2007. Rainfall interception and distribution patterns of gross precipitation around an isolated Ficus benjamina tree in an urban area. J. Hydrol. 333: 532-541. [ Links ]
16. Herwitz, S. R. 1985. Interception storage capacities of tropical rainforest canopy trees. J. Hydrol. 77: 237-252. [ Links ]
17. Holder, C. D. 2007. Leaf water repellency of species in Guatemala and Colorado (USA) and its significance to forest hydrology studies. J. Hydrol. 336: 147-154. [ Links ]
18. Huanhua, P.; Chuanyan, Z.; Zhaodong, F.; Zhonglin, X.; Chao, W.; Yang, Z. 2013. Canopy interception by a spruce forest in the upper reach of Heihe River basin, Northwestern China. Hydrol. Process. 28: 1734-1741. [ Links ]
19. Ibrahim, M.; Rapp, M.; Lossaint, P. 1982. Economie de l’eau d’un écosystème à Pinus pinea L. du litoral Méditerranéen. Ann. For. Sci. 39: 289-306.
20. Klaassen, W.; Bosveld, F.; de Water, E. 1998. Water storage and evaporation as constituents of rainfall interception. J. Hydrol. 212-213: 36-50. [ Links ]
21. Klos, P. Z.; Chain-Guadarrama, A.; Link, T. E.; Finegan, B., Vierling, L. A.; Chazdon, R. 2014. Throughfall heterogeneity in tropical forested landscapes as a focal mechanism for deep percolation. J. Hydrol. 519: 2180-2188. [ Links ]
22. Levia, D. F.; Frost, E. E. 2003. A review and evaluation of stemflow literature in the hydrologic and biogeochemical cycles of forested and agricultural ecosystems. J. Hydrol. 274: 1-29. [ Links ]
23. Leyton, L.; Reynold, E. R. C.; Thompson, F. B. 1967. Rainfall interception in forest and moorland. International Symposium on Forest Hydrology. Pennsylvania State University, Pergamon Press, W. E. Sopper, H. W. Lull (Eds.). p. 163-178. [ Links ]
24. Limousin, J. M.; Rambal, S.; Ourcival, J. M.; Joffre R. 2008. Modelling rainfall interception in a Mediterranean Quercus ilex ecosystem: Lesson from a throughfall exclusion experiment. J. Hydrol. 357: 57-66. [ Links ]
25. Llorens, P.; Poch, R.; Latron, J.; Gallart, F. 1997. Rainfall interception by a Pinus sylvestris forest patch overgrown in a Mediterranean mountainous abandoned area. I. Monitoring design and results down to the event scale. J. Hydrol. 199: 331-345. [ Links ]
26. Llorens, P.; Domingo, F. 2007. Rainfall partitioning by vegetation under Mediterranean conditions. A review of studies in Europe. J. Hydrol. 335: 37-54. [ Links ]
27. Lloyd, C. R.; Gash, J. H. C.; Shuttleworth, W. J.; Marques-Filho, A. 1988. The measurement and modelling of rainfall interception by Amazonian rain forests. Agr. Forest Meteorol. 43: 277-294. [ Links ]
28. Monteith, J. L. 1965. Evaporation and environment. In: Gash, J. H. C., Shuttleworth, W. J. (Eds.). Evaporation. Benchmark Papers in Hydrology. IAHS Press. Wallingford. p. 337-366. [ Links ]
29. Moreno-Pérez, M. F.; Roldán, J.; Cienfuegos, I. 2012. Interceptación de la lluvia por la vegetación en una cuenca hidrológica con clima mediterráneo. XXV Congreso latinoamericano de hidráulica. San José. Costa Rica. [ Links ]
30. Pereira, F. L.; Valente, F.; David, J. S.; Jackson, N.; Minunno, F.; Gash, J. H. 2016. Rainfall interception modelling: Is the wet bulb approach adequate to estimate mean evaporation rate from wet/saturated canopies in all forest types?. J. Hydrol. 534: 606-615. [ Links ]
31. Pérez-Arellano, R.; Moreno-Pérez, M. F.; Roldán-Cañas, J. 2016. Comparación de modelos de interceptación de agua de lluvia en individuos aislados de Pinus pinea y Cistus ladanifer. Ingeniería del Agua. 20.3: 153-168. [ Links ]
32. Ponce-Donoso, M.; Vallejos-Barra, O. 2016. Valoración de árboles urbanos, comparación de fórmulas. Revista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo. Mendoza. Argentina. 48(2): 195-208. [ Links ]
33. Rodrigo, A.; Avila, A. 2001. Influence of sampling size in the estimation of mean throughfall in two Mediterranean holms oak forest. J. Hydrol. 243: 216-227. [ Links ]
34. Rutter, A. J.; Kershaw, K. A.; Robiens, P. C.; Morton, A. J. 1971. A predictive model of rainfall interception in forests. 1) Derivation of the model form observations in a plantation of Corsican pine. Agr. Meteorol. 9: 367-384. [ Links ]
35. Rutter, A. J.; Morton, A. J.; Robins, P. C. 1975. A predictive model of rainfall interception in forests. II: Generalization of the model and comparison with observations in some coniferous and hardwood stands. J. Appl. Ecol. 12: 367-380. [ Links ]
36. Saxena, R. K. 1986. Estimation of Canopy Reservoir Capacity and Oxygen-18 Fractionation in Throughfall in a Pine Forest. Nord. Hydrol. 17: 251-260. [ Links ]
37. Simões, M. P.; Madeira, M.; Gazarini, L. 2009. Ability of Cistus L. shrubs to promote soil rehabilitation in extensive oak woodlands of Mediterranean areas. Plant Soil. 323: 249-265. [ Links ]
38. Xiao, Q.; McPherson, E. G. 2011. Rainfall interception of three trees in Oakland, California. Urban Ecosyst. 14: 755-769. [ Links ]
39. Zhang, Y.; Wang, X.; Hu, R.; Pan, Y.; Paradeloc, M. 2015. Rainfall partitioning into throughfall, stemflow and interception loss by two xerophytic shrubs within a rain-fed re-vegetated desert ecosystem, northwestern China. J. Hydrol. 527: 1084-109. [ Links ]
40. Zimmermann, A.; Zimmermann, B. 2014. Requirements for throughfall monitoring: The roles of temporal scale and canopy complexity. Agr. Forest Meteorol. 189-190: 125-139. [ Links ]
ACKNOWLEDGEMENTS
This study was developed as part of the project "Surface water balance in the El Cabril watershed funded by ENRESA and the University of Cordoba. The authors wish to thank the staff of the Department of Soil Engineering of ENRESA for their continued and effective cooperation.

