











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Fruit phenology studies are abundant in the scientific literature 22. However, although tree pruning is a widespread practice performed by small and large producers to optimize plant growth, experiments on the ideal branch size for maximum plant development are still incipient, especially in fruit species, and the divergence of most authors 4,5,17 with regard to this parameter raises concerns about its actual influence on plant development.
The introduction of temperate species has contributed to diversifying fruit production. Figs stand out in this regard due to their edaphoclimatic adaptability, becoming a viable production alternative as verified by previous studies conducted in northeastern Brazil, especially in the state of Ceará, generating high yields under the climatic conditions of the Apodi Plateau region 9, highlighting the importance of studies on the growth and phenology of temperate crops and their adaptability to warmer conditions 19. From this perspective, pruning also plays an essential role in favoring fig development by improving the canopy structure, controlling fructification, maintaining vigor, and increasing production in new branches.
Pruning management in fruit species is essential for crop development as it may stimulate production and accelerate or anticipate harvest depending on the climatic conditions of the cultivation area 3,6,16. However, studies on the ideal pruning intensity of fig branches are still incipient, and this parameter is hypothesized to interfere with fig development and production.
Growth curves are used to analyze phenological data, and since variations may occur among the individuals of a population, it is possible to use a Generalized Linear Mixed Model (GLMM), allowing to include variance components due to non-observed effects, including random effects in the linear predictor, in addition to fixed effects. Introducing random effects in the linear predictor allows modeling the correlation structure of observations performed on the same individual 11. The GLMM aims to describe changes in the mean response of each individual and their relationships with the covariables.
From this perspective, this study aimed to evaluate the growth and phenology of the fig cultivar ‘Roxo de Valinhos’ subjected to formative and production pruning.
Material and methods
Description of the study area
The study was performed from April 2018 to February 2019 in an orchard belonging to the Fruit Growing Study Group (FRUTAGRO) of the Federal University of Piauí (UFPI), Campus Professora Cinobelina Elvas (CPCE), in the municipality of Bom Jesus, Piauí, Brazil, 9°4’55” S and 44°19’39’’ W, at an elevation of 228 m above sea level. The experimental area, located in the semi-arid region of the state of Piauí, has a hot and humid climate with summer-autumn rainfall, with a rainy period from December to May and a dry period from June to November. The climate is classified as Awa according to the Köppen classification, with a mean annual temperature of 26.2°C and mean annual rainfall ranging from 900 to 1,200 mm year-1 (15. Figure 1 shows the meteorological data during the experimental period.

Conduction of the experiment
The area has a Dystrophic Yellow Latosol (Oxisol), corresponding to well-drained, slightly undulating, less fertile deep soils whose characteristics are shown in Table 1 (soil analysis of the experimental area), determined according to Teixeira et al. (2017).
Table 1: Physical and chemical characteristics of the soil in the 0-20 cm layer.

Organic matter - Walkley-Black method; Soil Analysis Laboratory, Campus Professora Cinobelina Elvas - CPCE, Bom Jesus, Piauí.
Materia orgánica - Método Walkley-Black; Laboratorio de análisis de suelos, Campus Professora Cinobelina Elvas - CPCE, Bom Jesus, Piauí.
Plant growth conditions
The fig plants were spaced at 2 m x 1.5 m. A drip irrigation system was adopted, with daily watering for two hours and one emitter per plant (flow rate of 40 L/h). Weed control was performed every 30 days using a hoe and a backpack brush cutter. Fertilization was split into three applications after soil analysis using ammonium sulfate (90 g per plant), single superphosphate (100 g per plant), and potassium chloride (120 g per plant).
In April 2018, formative pruning was performed in 27 fig plants at 50 cm from the ground to allow a single stem per plant. After emergence, the three shoots with the best conditions for canopy formation (vigor and healthiness) were chosen, while others were removed. Finally, one branch per plant was selected for phenological evaluation in the 27 plants of the fig cultivar ‘Roxo de Valinhos.’
Phenological analysis was performed over 12 evaluations considering the following periods: from pruning to the beginning of sprouting, at the beginning of harvest and at its end, and during the harvest period. The beginning of sprouting was defined by the emergence of the first shoots; the beginning of harvest was determined by the onset of fruit ripening, with fruit color changing from green to purple; finally, the harvest period ended when most harvested fruits were abnormally colored (uneven and low-intensity purple color) and showed a fibrous pulp, considered unfit for fresh consumption 14.
The following variables were evaluated weekly starting from 30 days after formative pruning: branch length (cm), measured from the base of the branch to its apex with a millimeter ruler; branch diameter (cm), measured at the base of the branch with a portable digital caliper (150 mm, accurate to 0.01 mm); number of leaves per branch; number of nodes per branch; number of shoots per branch; and number of inflorescences per branch. In addition, the number of fruits on each branch was counted every seven days (evaluation days), from their formation until reaching 100% maturation. Harvest was halted when green and small fruits that would not resume ripening were observed.
Production pruning was performed in the 27 fig plants in September 2018. The process consisted of three pruning treatments (10, 20, and 30 cm) with nine replications each. The analyses performed during the present experimental stage were similar to those of the previous stage, but their frequency was fortnightly, not weekly, totaling eight evaluations.
Statistical analysis
The quantitative variables were evaluated using a generalized linear mixed model (GLMM). The model assumed that the response variables could be affected by the following fixed factors: branch size after pruning (10, 20, and 30 cm), time after pruning (30, 60, 90, and 120 days), and the interaction between these two factors (branch size X time after pruning). The effect on each plant was considered a random factor to model the correlation structure between observations of the same individual.
The generalized linear mixed model with a Poisson distribution was specified as follows:

The relationship between the expectation and the linear predictor was given by the log link function
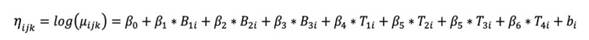
The generalized linear mixed model with a normal distribution was specified as follows:
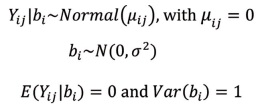
In this case, the relationship between expectation and linear predictor was given by an identity link function unlike the one previously described, in which Y ij is the response variable (branch length, branch diameter, number of leaves, nodes, flowers, fruits, and shoots) of the i-th plant with branch size j in moment k , with i = 1,?, 27, j = 10, 20, 30, and k = 30, 60, 90, 120; μ i is the mean number expected in plant I with branch size j and in moment k ; b i is the random intercept, and σ b is the estimate of variability of the response variable in plant i. It should be noted that branch length and diameter (measurable characteristics - continuous data) were evaluated by the normal model, while the other characteristics (counting characteristics - discrete data) followed the Poisson model.
All statistical procedures mentioned below are contained within the Statistical Analysis Systems software (University Edition version). Descriptive statistics of the data on the response variables were obtained by the MEANS procedure. These data were analyzed by considering a model of analysis of variance using the Normal and Poisson distributions with the GLIMMIX procedure, and the means were adjusted using the lsmeans command (least-squares means). Statistical significance was checked by the F-test, and the means were compared by the Tukey-Kramer test. In all analyses, significance was established at p ≤ 0.05. Spearman’s rank correlation coefficient was calculated using the CORR procedure. The SigmaPlot 11.0 software was used to generate the plots of the characteristics evaluated.
Results and discussion

In general, the earlier the beginning of sprouting, the shorter it takes for flowering to occur 12. Although a typically temperate fruit species, figs have minimal low-temperature requirements and sprout almost immediately after leaf fall if the temperature remains high 8, as verified at the time of sprouting.
The phenology of fruit species is affected by temperature, and phenological stages such as sprouting, vegetative growth, and harvest can be anticipated at higher temperatures. Under these conditions, plants can absorb and transport nutrients more effectively, favoring their development and vigor.
When studying the fig cultivar ‘Roxo de Valinhos’ subjected to pruning in the region of Lavras, MG, at 28°C and relative humidity of 67%, Norberto et al. (2001) observed 162 days between pruning and harvest for plants pruned in July, whereas, in the present study, 109 days were observed for plants pruned in April. This result suggests that, under the semi-arid conditions of the state of Piauí, the high temperatures shortened the production cycle of the fig cultivar ‘Roxo de Valinhos’.
The values found in the present study for the time between pruning and harvest are close to those found by Silva et al. (2017) when studying fig phenology and production at 27°C, relative humidity of 68.9%, and July pruning, obtaining 130 days for plants grown in the western region of Rio Grande do Norte.
The crop cycle variations observed may have been due to differences in temperature oscillation, as observed by Souza et al. (2009) when studying basal temperatures and the thermal sum of fig plants pruned at different times, resulting in 139 days from pruning to the beginning of harvest for plants pruned in August and grown at 36°C in Botucatu, São Paulo. These results corroborate the present findings as the fig cycle was shortened with the increase of temperature (Figure 1, page 15).
Fig fructification requires full sunlight as vegetative buds are quickly differentiated into reproductive buds when temperature increases, increasing the demand for leaf carbohydrates produced by photosynthesis. From this perspective, the high rainfall in April, coinciding with pruning, stimulated the storage of water and nutrients, essential components for bud differentiation.
Plant phenology is dependent on water availability, as clearly observed in semi-arid regions, which alternate between dry and wet periods 19. In Brazil, especially in its northeastern semi-arid region, the conditions and phenological patterns of plants tend to follow the seasonal rainfall fluctuation, causing mild and short-term stresses that result in anticipated flowering and fructification, thus modifying the plant cycle 1.
Branch length, branch diameter, the number of leaves, nodes, and shoots showed a linear growth trend. Their values increased gradually as a function of the evaluation times, reaching the mean values of 31.26 cm per branch, 0.97 cm per branch, 17.56 leaves per branch, 14.24 nodes per branch, and 11.05 shoots per branch after 77 days, respectively (Figure 2, page 19).

Figura 2: Fenología del cultivar de higo ‘Roxo de Valinhos’ sometido a poda formativa en función del número de días después de la poda. Longitud de la rama (3A); Diámetro de la rama (3B); Número de hojas (3C); Número de nodos (3D); Número de brotes (3E).
Branch length showed a nonlinear growth trend (Figure 2A, page 19). This variable increased by 0.64 cm per day, reaching the final value of 1.15 cm per branch after 77 days. According to Figure 2B (page 19), branch diameter reached its maximum growth rate after 63 days, corresponding to 32.61 mm. Considering the initial branch diameter, this variable increased by 17.88%.
High vegetative growth rates occur due to high temperatures after pruning (Figure 1, page 15), resulting in competition between processes that demand photoassimilates, such as branch and shoot growth and fruit development.
The number of leaves also showed a nonlinear growth trend as evaluations succeeded, reaching 33.70 leaves per branch after 77 days (Figure 2C, page 19). These values are higher than those reported by Silva et al. (2011) when studying fig growth under different conditions, obtaining the lowest and highest numbers of leaves per branch of 13.32 and 15.87, respectively.
Celedonio et al. (2013) analyzed three types of fig cultivation in Limoeiro do Norte, CE, and observed that plants grown in the field had 44 leaves per branch after 90 days. However, it should be noted that the climatic conditions of Limoeiro do Norte differ from those of the present study.
The number of leaves decreased in the period from 42 to 49 days after pruning (Figure 2C, page 19), consequently affecting the number of nodes (Figure 2D, page 19) and shoots (Figure 2E, page 19) as these structures emerge from the leaf axil. This reduction may have been due to the high temperatures and low relative humidity of the region (Figure 1, page 15), reducing plant metabolic activity for ± 2 months, as evidenced by leaf fall and the presence of fewer nodes and shoots.
With regard to the number of fruits per branch, despite the value of 0.67 for this parameter (Figure 3, page 20), which is considered below-average compared to studies such as the one by Silva et al. (2017), it should be noted that the plants were still in early development.

Moreover, the primary purpose of formative pruning is not to produce marketable fruits immediately but rather to develop good canopy architecture and vegetative vigor, stimulating nutrient uptake and storage for the reproductive period and preparing the plants for production pruning.
Vegetative development depends on the uptake and processing of essential materials for plant growth, such as water, energy, carbonic gas, and nutrients. This process implies changes in the internal relationships of the crop with the external environment and is directly associated with the cultivation area, rainfall, temperature, humidity, and severe stresses to which plants may be subjected, such as pruning 2.
The fig plants showed uniformity and typical branch growth after production pruning, although with late sprouting and fruit formation compared to the period after formative pruning, with an average of 70 days. There was a 49-day period from fruit formation to the beginning of harvest, one day shorter to formative pruning, after which the fruits were fit for fresh consumption (Table 3, page 20).

The high temperatures explain the late sprouting and fruit development observed, inducing physiological delay as the plants responded with slower growth regardless of branch size.
Phenological studies conducted by Ferraz et al. (2017) with the fig cultivar ‘Roxo de Valinhos’ pruned at 20.3°C and relative humidity of 58.3% in the region of Botucatu, SP, showed that the period between pruning and sprouting was 13.69 days for plants pruned in August, while the period between the beginning of sprouting and the beginning of harvest was 155 days. These values are above those of the present study and highlight that the production cycle of the fig cultivar ‘Roxo de Valinhos’ was shortened due to the high temperatures of the semi-arid region of Piauí.
The ratio between the generalized chi-square statistics and its degrees of freedom is close to 1 (Table 4, page 21).
Table 4: Analysis of variance of the effects included in the model for the studied characteristics in fig plants.

χ2 - chi-square; DF - degree of freedom.
χ2 - chi-cuadrado; DF - grado de libertad.
This indicates that data variability was adequately modeled and there was no residual overdispersion (variance higher than the mean).
No significant difference was observed (p>0.05) for branch size and the branch size x day interaction with regard to the characteristics evaluated. However, there was a significant difference for the day factor (Table 5, page 21).
Table 5: Adjusted means of fig characteristics as a function of the number of days after pruning.

Means followed by different lowercase letters, per characteristic, differ from each other by the Tukey-Kramer test (p<0.05).
Medias seguidas de diferentes letras minúsculas, por característica, se diferencian entre sí por la prueba de Tukey- Kramer (p <0,05).
The variance component model was adopted to structure the residual (co)variance of measurement variations for each individual.
The Tukey-Kramer test highlighted that the number of days after pruning influenced plant development. This was expected as the longer the time, the more the crop develops. However, there was no significant difference in the number of leaves, nodes, and shoots from 90 to 120 days after pruning, implying that branch size reached a ‘standard value’ 90 days after pruning. In other words, the plants reached statistically similar measurements, growing and developing as if they had not been subject to production pruning.
In the present experience, the optimal climatic conditions for plant development should also be noted, with well-distributed rainfall, high temperatures, and high relative air humidity, promoting uniformity among plants even when subjected to the stress caused by production pruning.
Branch length showed a linear growth trend (Figure 4A, page 22), with a constant daily increase of 0.34 cm.

Figura 4: Fenología del cultivar de higo ‘Roxo de Valinhos’ en función de mediciones repetidas a lo largo del tiempo.
After 120 days, the mean growth rate was 0.99 cm per branch. On the other hand, Figure 4B (page 22), shows that branch diameter reached its maximum growth rate of 32.61 mm after 60 days. Based on the initial branch diameter, this variable increased by 14.66%. Branch length and diameter showed linear growth trends, and their values increased gradually as a function of the evaluation times, reaching 41.18 cm per branch and 1.24 cm per branch, after 120 days. A similar situation occurred for the number of nodes (Figure 4D, page 22) (19.66 nodes per branch).
A positive response was verified for the number of leaves (Figure 4C) and shoots (Figure 4E), which showed quadratic growth curves. The measurements reached their maximum potential 90 days after production pruning, with 18.54 leaves per branch and 14.91 shoots per branch. These observations show that, in general, the responses of closer times in repeated measures experiments are more strongly correlated than those of more distant times. The low rainfall rates after pruning may have delayed fruit formation and contributed to the non-significance of the number of fruits observed in the present study, especially for the cultivar ‘Roxo de Valinhos,’ which fructifies well under ideal rainfall condi tions. However, the results for the vegetative phase were promising.
Some of the characteristics evaluated were correlated, such as branch length with branch diameter and the number of nodes with the number of shoots (Table 6, page 23).

Vigorous fig branches develop both in length and diameter, supporting the leaves properly.
The correlation between the number of nodes and the number of shoots was high as nodes are necessary for shoot emergence. However, in cases of very severe pruning, shoots may emerge in random spots of the plant, which did not occur in the present study.
Conclusion
The fig cultivar ‘Roxo de Valinhos’ was well adapted to the semi-arid region of Piauí and showed precocity at all phenological stages compared to fig plants cultivated in temperate regions.
With regard to production pruning, branch size as a function of time did not influence fig development according to the characteristics evaluated.

