











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCTION
Algae are a heterogeneous group of autotrophic organisms that are the basis of the trophic webs, although the massive development of cyanobacteria and contamination by cyanotoxins constitute an environmental problem in aquatic systems, due to the various negative impacts they can cause (Bordet et al., 2017).
Algae removal, particularly those with toxicogenic potential, from water collected for supply and human consumption constitutes one of the indispensable requirements to be met by companies responsible for the use of surface water for potabilization (Aguilera et al., 2018).
In pilot plants designed to evaluate water disinfection by ozone, the algae that enter the system in the raw water and those that remain after the treatment process are monitored on a daily basis. The concentration of algae entering the system depends on the environmental variables that affect the water body from which the liquid is extracted (e.g. reservoirs) and, obviously, after treatment, on the system’s own operating variables: flow rate used in the pilot test, ozone dose used (g/m3), mixture and contact time, and the value of the hydraulic flow used. Obtaining the optimum working point is evaluated in different operating scenarios of the ozone pilot plant. These vary according to the inlet water flow rate, ozone flow rate gO3/h, contact time in minutes and residual dose of ozone in mg/l.
The recording of phytoplankton diversity, abundance and fluctuations in phytoplankton composition and the effects of ozonation treatment are usually considered technical aspects used to size the operation of the final plants and therefore are not usually published. This work details the variation in the quali-quantitative composition of the microalgae that entered and exited an ozone pilot plant, at the outlet of the Campo Alegre reservoir in the province of Salta (Argentina), during the period of operation, which covered the winter, spring and summer months of 2019 and autumn 2020. This information, together with other technical aspects of the operation of the pilot plant, will be used for the implementation of the definitive treatment plant that will allow a production of 4,000m3.h-1 which, through an aqueduct, will supply water to a population of 200,000 inhabitants of several adjacent towns and large sectors of the city of Salta, characterized by deficient service and lack of coverage, which have caused different environmental problems that have an impact on anthropic communities.
The objective of the work was to evaluate the removal capacity of microalgae in an ozone pilot plant as a function of two cell size classes: MDL (maximum linear dimension, in μm) and biovolume (in μm3) between the autumn of 2019 and the autumn of 2020, and to establish seasonal differences in the structure and diversity of the algal communities present.
MATERIAL AND METHODS
The Campo Alegre reservoir, located in the central part of the province of Salta, Argentine (24°33’42’’S 65°22’05’’W), is an important surface water reserve for supplying an estimated 200,000 inhabitants of the metropolitan area of Salta Capital and adjacent towns. This reservoir, located at 1444 meters above sea level, was built in the 1970s and has a capacity of 48 hm3 and a surface area of 322 ha with a maximum depth of 40 meters. The dam is fed by the waters of the Santa Rufina and San Alejo streams. The current water uses are irrigation (main) and recreational use. The period of maximum water consumption for irrigation occurs in the four-month period August-November.
The intake of water from the dam and adduction to the definitive drinking water treatment plant, which is in the final stage of construction, is expected to perform the complete treatment of raw water with coagulation, flocculation, sedimentation and filtration, including the removal of algae from the reservoir. The study covered from June 2019 to June 2020, with a total of 320 samples, distributed as follows; 2019: autumn 34 samples, winter 88 samples, spring 100 samples, summer 66 samples, and 32 samples in the autumn of 2020. Microalgae concentration 50% were collected at the inlet and 50% at the outlet of an ozone pilot plant. Each sample was preserved in situ with Lugol in 300 ml plastic bottles and stored at 4°C until quantitative analysis. Algal identification was made from samples collected by 25-micron plankton net, concentrated and fixed with 4% formalin in 300 ml bottles. Images of the species were captured with an Axio Cam 1Cc3 digital camera, in a Zeiss AxioVert 4L microscope, with 40x, 63x and 100x magnification. For diatoms, organic matter was removed according to the methodology of Battarbee (1986), using hydrogen peroxide and heat. The classification system adopted was based on algae base (Guiry & Guiry, 2020).
The abundance of microalgae (individuals.ml-1) was quantified after sedimentation in combined chambers, with a Zeiss L inverted microscope, using the technique of Utermöhl (1958). Colonies, filaments and individual cells were considered as counting units.
Numerous reference literature was used for taxonomic determinations including Bicudo & Meneses, 2006; Joosten, 2007; Komárek & Fott, 1983; Komárek & Anagnostidis, 1999, 2005; Krammer & Lange-Bertalot, 1986, 1991; John et al., 2011; Sant’ Anna et al., 2012; Mc Gregor, 2013.
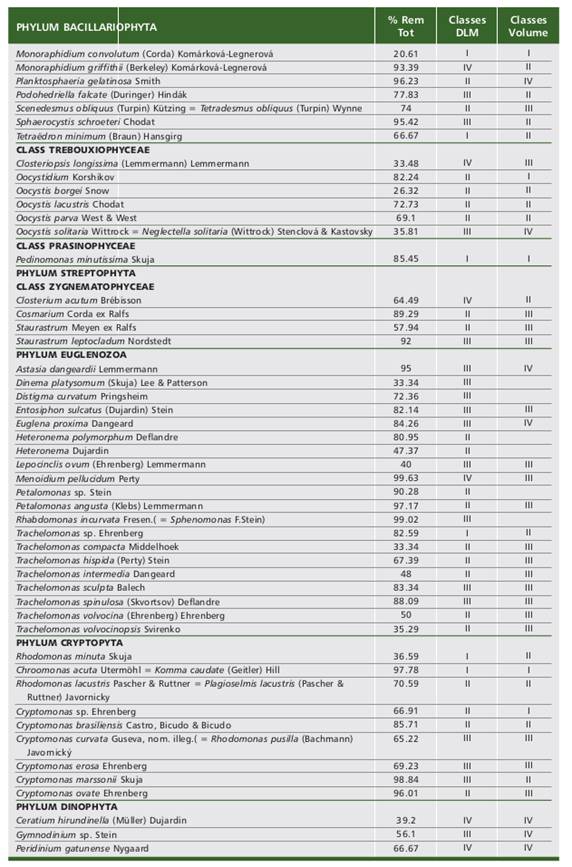
The phytoplankton structural variables (density, species richness, Shannon-Weaver diversity log2 and composition of main taxonomic groups) were compared to analyze the variability between phytoplankton input and output in the pilot plant during the different climatic periods of the 2019-2020 study cycle. To this end, a non-parametric analysis of variance was performed using the Kruskal-Wallis (H) test, since the variables analyzed did not meet all the requirements of normality or homogeneity of variance. In addition, a group of species that were present in at least 10% of the sampling was selected, and for each one, the average maximum linear dimension (MLD, in μm) and the average biovolumes (in μm3) were estimated to correlate them (Pearson’s correlation coefficient) with the total removal percentages of each species in the treatment.
The cell metrics used were extracted from Fernández (2010), Kruk et al. (2010), Medeiros Fonseca et al. (2014). Species were catalogued according to the categories suggested by Crossetti & Bicudo (2008) who classify both DML (I < 10 μm, II: 11-20 μm, III of 21 and 50 μm and IV > 50 μm) and biovolumes (I <102 μm3, II 102-103 μm3, class III 103to 104 μm3 and class IV >104 μm3) into four classes. Infostat statistical software (Di Rienzo et al., 2018) was used.
RESULTS
Phytoplankton in plant inlets
In the first year, a total of 153520 ind.ml-1 entered the plant, and an average of 994. 91±82ind.ml-1 of phytoplankton, with a maximum value of 7722ind.mL-1 on June 6, 2019, and a minimum of 47ind.mL-1 on November 26, 2019.
The abundance of phytoplankton showed seasonal differences with higher records in the winter of 2019 compared to the other periods sampled (Table 1).
In the structural conformation of phytoplankton, a large seasonal variation was observed (Fig. 1). Trebouxiophyceae were abundant only in the first fall with a maximum of 57% and then presented minimum percentages equal to or less than 1.12%. Bacillariophyceae (diatoms) predominate in the winter with a maximum of 64% and minimum of 4.42% in the summer of 2019. Cyanobacteria only dominated in the summer (70.8%) and Chlorophyceae in the second autumn with 47.12%. Cryptophyceae ranged from 1.66 to 5.74% in autumn and winter respectively, they were also constant in the other periods. The remaining minority groups (Prasinophyceae, Pyramonadophyceae and Pedinophyceae) together only had a percentage representation of 1.5% in spring 2019 and 3.7% in autumn 2020, and in the remaining three seasons they did not exceed 0.09% of the total
Table 1 Tabla 1 Phytoplankton abundance (ind.ml-1) in the pilot plant inflow, discriminated by climatic season. Abundancia del fitoplancton (ind.ml-1) en el ingreso a la planta piloto, discriminado por estación climática.
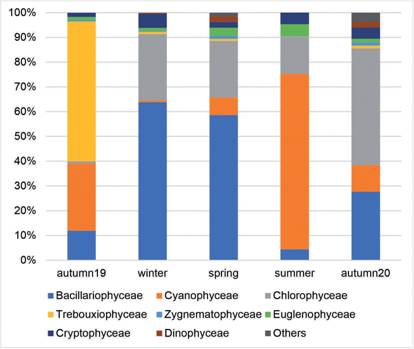
Fig. 1 Fig. 1 Phytoplankton groups in percentages represented in each climatic season at the pilot plant inlet. Grupos del fitoplancton en porcentajes representados en cada estación climática al ingreso a la planta piloto.
Statistical analysis determined a significantly higher abundance of Bacillariophyceae, Chlorophyceae, Trebouxiophyceae and Cryptophyceae in winter, while Dinophyceae were significantly higher in spring and Cyanobacteria in summer (Table 2).
The diatoms that entered the plant presented a very low density except for Cy-clotella ocellata (66921 ind.ml-1), which was the most abundant species, and Dicostella pseudostelligera (2270 ind.ml-1). The chlorophycean Monoraphidium convolutum was the second most abundant species (22784 ind.ml-1). In general, the species recorded at entry presented low values compared to those at exit and in decreasing order were: Rhodomonas minuta (2244 ind.ml-1), Pseudanabaena mucicola (1690 ind.ml-1), Ceratium hirundinella (1393 ind.ml-1), Cryptomonas ovata (1302 ind.ml-1) and Pseudanabaena catenata (1228 ind.ml-1).
Some cyanobacteria such as Aphanothece smithii (216 ind.ml-1), Synechococcus elongatus (237 ind.ml-1) and Synechococcus lividus (1239 ind.ml-1) had higher densities in spring, while others such as Merismopedia tranquila (227 ind. ml-1), Schizothrix facilis (251 ind.ml-1) and Pseudanabaena catenata (603 ind.ml-1) were more abundant in autumn 2020. Pseudanabaena mucicola represents the only case of cyanobacteria that reached comparatively higher values (1665 ind.ml-1) in summer.
Phytoplankton in the plant outflows
In 2019, an average of 763.80±69 ind.ml-1 of phytoplankton exited the plant, with a maximum of 3853 ind.ml-1 on 20 August 2019 and a minimum of 0 ind.ml-1 on 11 November 2019. There were also seasonal differences in the amount of algae detected at the treatment outlet over the study period with higher densities during winter (Table 3).
Table 3 Tabla 3 Seasonal differences in phytoplankton density (ind.ml-1) at discharge from the pilot plant in the period 2019-2020. Diferencias estacionales en la densidad del fitoplancton (ind.ml-1) en la descarga de la planta piloto en el período 2019-2020.
The winter period was the most productive in biomass, particularly because of the presence of small diatoms that were able to elude treatment effects. The different classes of algae recorded at the plant outlet differed between periods of the climatic cycle, being more abundant in winter: Bacillariophyceae (1223±47) (F=112.68); Chlorophyceae (462.89 ±18) (F=120.15); Cryptophyceae (66.23±11) (F=8.31) and Euglenophyceae (14.43±1.46) (F=9.61) (with highly significant p<0. 00001) and Dinophyceae (9.08±1.28) (only significant p=0.0069).
On the other hand, Cyanobacteria presented higher densities in autumn (185±11) (F=73.81), with no differences in the remaining groups between seasons in the study.
The remaining classes were poorly represented in all periods and did not exceed 2%, except for Trebouxiophyceae with 12.22% in autumn 2019, Cryptophyceae in winter (3.65%) and Dinophyceae in spring (2.82%) (Fig. 2).
In all monitoring, few species exited the plant in appreciable quantities: Cyclotella ocellata (58725 ind.mL1), Monoraphidium convolutum (18088 ind.ml-1) and M. circinale (1049 ind.ml-1).
Phytoplankton comparison between pilot plant input and output
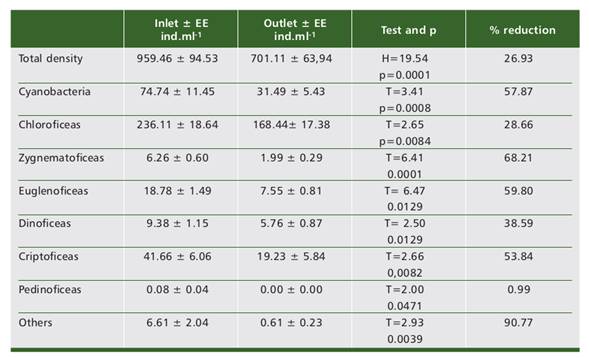
The treatment applied in the pilot plant produced a significant reduction in the average algal density (26.94%), which increased even more in some particular groups: Zygnematophyceae (68.21%), Euglenophyceae (59.80%), Cyanobacteria (57.87%),
Cryptophyceae (53.84%), Dinophyceae (38.59%) and Chlorophyceae (28.66%) (Fig. 3). A total of 326 species were recorded in the monitoring, although only 126 had a frequency of occurrence in at least 10% of the dates and some significance in the relative densities being listed some selected species in Table 4. The distribution of species among groups was: Bacillariophyceae 56 spp, Cyanobacteria 105 spp, Chlorophyceae 65 spp, Euglenophyceae 55 spp, Trebouxiophyceae 14 spp, Zygnematophyceae 11 spp, Cryptophyta 11 spp, Prasinophyceae 5 spp, Dinophyceae 3 spp, and Chrysophyceae 1 species.
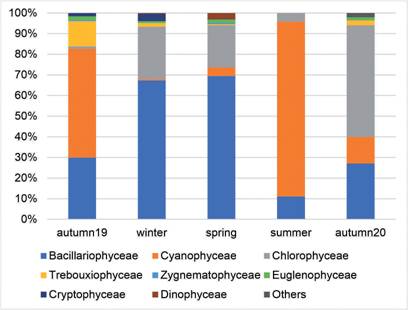
Fig. 2 Fig. 2 Phytoplankton groups in percentages represented in each climatic season in the pilot plant discharge Grupos del fitoplancton en porcentajes representados en cada estación climática a la salida de la planta piloto.
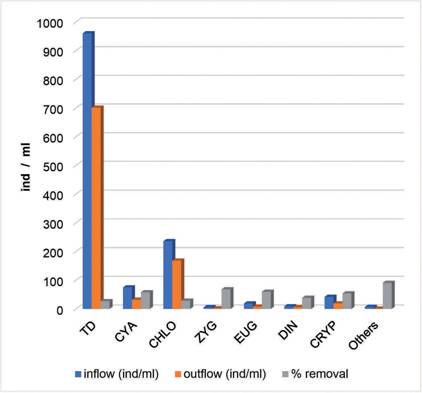
Fig. 3 Fig. 3 Comparison of relative abundance of algae groups at plant inflow and outflow and % removal efficiency per group Comparación de las abundancias relativas de los grupos de algas entre el ingreso y el egreso de planta y % de eficiencia de remoción por grupo.
Table 4 Tabla 4 (1/2) Table 4 (2/2) Tabla 4 (2/2) (1/2). Removal of selected species with cell size classes. Remoción de especies seleccionadas con clases de tamaños. Removal of selected species with cell size classes. Remoción de especies seleccionadas con clases de tamaños.
Although there was consistency between the amount of algae entering versus that exiting the plant, it can be seen that only in the winter and spring period of 2019 there were significant increases, which occurred mainly at the expense of Cyclotella ocellata. It should be clarified that the procedures employed from October 2019 onwards involved the stages of coagulation, flocculation, sedimentation and filtration in addition to the application of variable doses of ozone.
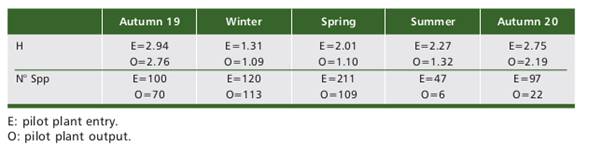
In the different climatic seasons, the number of species and algal diversity at the entrance and exit of the ozonation plant (Table 5) varied significantly, with winter being the phase with the lowest diversity value and summer the phase with the lowest relative number of species.
Although a total of 297 species entered the plant with a total average diversity of 2.26 during the entire monitoring period, at the end of the process and as a result of this, only 187 species exited with a final average total diversity of 1.34.
Removal efficiency versus cell dimensions
The phytoplankton removal percentages did not show significant correlations with the maximum linear dimension (MLD) or with the biovolumes of the selected species (Table 4). However, the removal of categories III and IV was slightly higher for both cell dimensions.
The percentages of removal according to MLD of categories I (63.21±31.29) and II (67.54±24.83) were lower than the removals of classes III (72.97±22.26) and IV (71.00±26.25).
In the case of biovolumes also Classes I (68.27±28.02) and II (64.23±29.57) had lower removal percentages than Categories III (72.74±22.66) and IV (70.34±20.62).
On the other hand, the two MLD classes (in μm) with the highest number of species were Class II with 34 spp. and Class III with 33 spp; whose life habits corresponded mostly to unicellular flagellated or non-flagellated colonial individuals. For example, species of the genera Cryptomonas, Petalomonas, Trachelomonas, in the first case and Desmodesmus, Coelomoron, Scenedesmus, Merismopedia in the group of colonial /cenobial species.
In the case of the classes established according to cell biovolumes (in μm3), a greater number of species was also observed in Classes II with 33 spp. and III with 38 spp., whose morphologies were also predominantly non-flagellated colonial forms in group II and flagellated unicellular forms in group III with the latter class comprising practically the majority of species with this life habit.
In summary, the average cell dimensions of MLD =31.62 ±8.79 μm and average biovolume=4019.69 ±2711.49 μm3, both Class III° had the highest removals in the ozone treatment (72.97% and 72.74%, respectively), although they were not statistically different from the remaining classes.
DISCUSSION
Conventional treatment of raw water through physical and chemical clarification processes (coagulation, flocculation, sedimentation and filtration) can be complemented with ozone disinfection. Ozone is a disinfectant of bacteria, viruses, parasites and an aid in micro-flocculation and reduction of suspended solids. It has also been shown to control algae and pathogens by oxidation of cell wall elements and in the interior by oxidizing cell membranes, enzymes and nucleic acids (Zhang et al., 2014). Its oxidizing and disinfecting power, greater than that of chlorine, makes it more effective than chlorine in removing odor, taste and color from water. It does not produce trihalomethanes and eliminates their precursors, requires a lower concentration and contact time to eliminate microorganisms (0.4 ppm for 4 minutes), does not alter the pH of the water, improves coagulation and facilitates the elimination of iron and manganese (Barrado Moreno et al., 2016).
In the Campo Alegre reservoir, raw water enters the ozone pilot plant from the dam discharge valve, located at the base of the dam wall at a depth of 10 meters, through an adduction aqueduct of 1.770 meters long. Obtaining the optimum working point in the pilot plant was evaluated in different operating scenarios that varied according to the inlet water flow rate from 4 to 10 m3.h-1 being the ozone flow rate from 20 to 30 gO3.h-1, and the contact time from 2.5 to 6 minutes with a residual dose of 3 to 7.50 mg.l.-1.
The treatment had a variable microalgae removal efficiency. Among the diatoms, except for Dicostella pseudostelligera with 82.2% efficiency, the removal percentages of the smaller species were low, e.g. Cyclotella ocellata (12.25%), and the same was true for species with large maximum linear dimensions (MLD) such as Aulacoseira granulata (31%) and Nitzschia palea (54.7%).
Species included in the picoplankton (DLM<2μm) such as Cyanobium gracile and in the nannoplankton (2-20 μm) such as the genera Synechocystis and Synechococcus belonging to Synechococcales of Class I were abundant in both plant entry and exit and their frequency of removal was variable (Table 4). Picocyanobacteria are abundant in tropical freshwater lenitic environments (Callieri, 2010).
Zygnematophyceae of Classes III and IV such as Staurastrum leptocladum and Closterium acutum, Cosmarium sp. and Staurastrum sp. presented intermediate to high removal efficiencies.
Large filaments such as some Nostocales and Oscillatoriales of group IV (Planktothrix, Oscillatoria or Phormidium) were not relevant in the sampling probably because of their low sinking rate which did not facilitate their entry to the plant.
Flagellate euglenoids with loricae of the genus Trachelomonas, or without heterotrophic cell covers such as members of the genera Dinema, Heteronema and Menodium also presented removal values in a wide range (30-97 %), their typical habitat being sediments (Al-Qassab et al., 2002).
Dinoflagellates such as Ceratium hirundinella, and the genera Gymnodinium and Peridinum of group IV (> 50 μm) presented low removal percentages. Ceratium during the study period presented a constant presence in the photic zone of the reservoir, and at the plant outlet it appears fragmented in multiple pieces.
CONCLUSION
The algal removal capacity by the ozonation process in a pilot plant that captures water from the discharge valve of the Campo Alegre dam produced a significant reduction in algal density, particularly of Cyanobacteria, Zygnematoficeae, Euglenophyceae and Cryptophyceae, which in all cases exceeded 50%. At the level of genera and species, the reduction in density at the end of the process was variable and did not present a significant correlation with the two cellular dimensions DLM and phytoplankton biovolumes, influenced by the variation of the flow and period of contact with the chemical during the test.

