











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCTION
Azhdarchids represent a derived clade of long-necked pterodactyloid pterosaurs that were distributed in almost all continents, with excep tion of Antarctica (Averianov, 2014) and probably Australia (Andres, 2021). Most azhdarchids are known from Upper Cretaceous deposits (Andres et al., 2014; Longrich et al., 2018), with some pos sible records from Lower Cretaceous and Upper Jurassic deposits (Costa et al., 2015; but see Andres, 2021). In spite of their wide geographi cal distribution, their record in South America is still very poor. Possible azhdarchid remains were reported from the Lower Cretaceous beds from the Crato Formation, Brazil (Martill & Frey, 1998), but its identification was heavily put into question by subsequent authors that regarded them as indeterminate tapejarids or pterodacty loids (Unwin & Martill, 2007).
More recently, Kellner & Calvo (2017) de scribed the new genus and species of azhdarchoid Argentinadraco barrealensis from the Portezuelo Formation (Late Turonian-Early Coniacian) from Neuquén Province, Argentina, based on an incomplete lower jaw. Based on its general shape, Kellner & Calvo (2017) suggested that it may be allied to azhdarchids. However, this taxon has been recently found to be included in Thalassodrominae among Dsungaripteromorpha by Andres (2021).
The first undoubted record of azhdarchid in South America belongs to Aerotitan su damericanus, a large-sized azhdarchid from Maastrichtian beds of northwestern Patagonia (Novas et al., 2012). Because Aerotitan is known just by the tip of a beak, some authors doubted its affinities (Longrich et al., 2018), but its unique morphology indicates that it belongs to a very large azhdarchid closely related to Quetzalcoatlus (Averianov, 2014; Vullo et al., 2018; Pegas et al., 2022). More recently, Ortiz-David et al. (2018, 2022) described the azhdarchid genus and spe cies Thanatosdrakon amaru, based on postcra nial remains from the Plottier Formation (Late Coniacian-Early Santonian, Neuquén Basin), Mendoza Province, Argentina.
The aim of the present contribution is to describe a single, partial cervical vertebra as signable to Azhdarchidae from Cenomanian beds of the Candeleros Formation (early Late Cretaceous) from northern Patagonia (Fig. 1). This represents the oldest record for this clade in South America.
Fig. 1 Map showing the fossiliferous locality (star) where MPCA-Pv 2003 was found (modified from Novas et al., 2012).
MATERIAL AND METHODS
Locality and horizon
The specimen here reported was found close to the bonebed that yielded the skeletal remains of the coelurosaur theropod Bicentenaria argen tina (Novas et al., 2012). The specimen was found during a field trip performed under the project of relocation and georeferencing of fossil sites in the Río Negro province, carried out by the former Dirección de Patrimonio y Museos from the Río Negro goverment and supported by some profes sionals of the Universidad Nacional de Rio Negro (UNRN) in June, 2019. The fossiliferous locality is placed on the east shore of Ezequiel Ramos Mexía Reservoir, in the northwestern part of the Río Negro Province, Northern Patagonia, Argentina. GPS coordinates of the fossil site are 39° 28’ 9.82” S, 68° 54’ 25.01” W (Novas et al., 2012) (Fig. 1). The specimen here described was found together with some turtle plates belonging to a small Chelidae.
These fossils come from the upper levels of the Candeleros Formation (Early Cenomanian, early Late Cretaceous; Leanza et al., 2004), Río Limay Subgroup, Neuquén Group. The rocks that yield ed the remains of Bicentenaria and the pterosaur here described are composed of red medium-sized sandstones and mudstones, with abundant dark brownish paleosoils (Leanza et al., 2004; Garrido, 2010, 2011). These beds represent a fluvial envi ronment under braided and meandering regimes, as well as aeolian conditions (Garrido, 2010).
SYSTEMATIC PALEONTOLOGY
Pterosauria Kaup, 1834
Pterodactyloidea Plieninger, 1901
Azhdarchoidea Unwin, 1995 (sensu Kellner, 2003)
Azhdarchidae Nessov, 1984
Indeterminate genus and species
Referred material. MPCA-PV 2003, posterior half of a middle cervical vertebra (Fig. 2A-D).
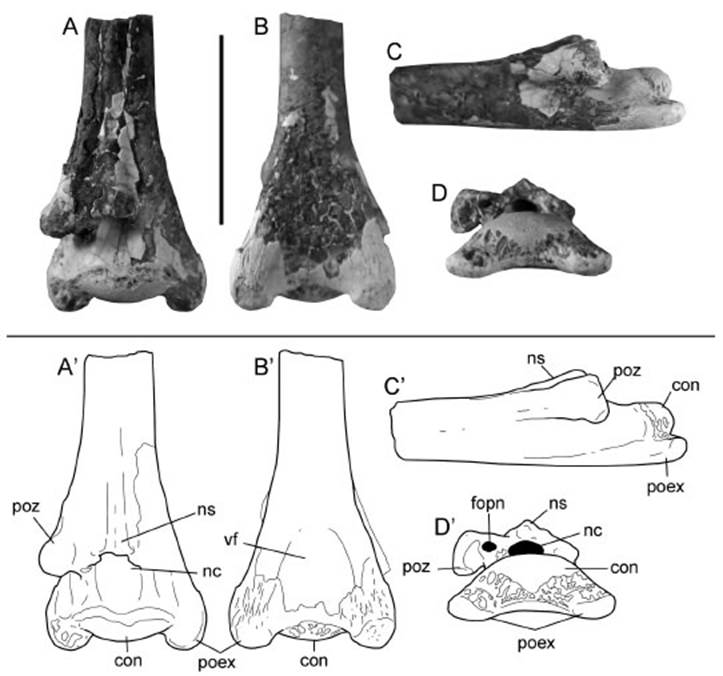
Fig. 2 Azhdarchid cervical vertebra (MPCA-PV 2003) in (A, A’) dorsal, (B, B’) ventral, (C, C’) lateral and (D, D’) posterior views. Abbreviations: con, posterior condyle; fopn, pneumatic foramen; nc, neural canal; ns, neural spine; poex, postexapophyzes; poz, postzygapophysis; vf, ventral fossa. Scale bar: 3 cm.
Description. The specimen MPCA-PV 2003 consists of the posterior half of a cervical verte bra having the surface of the centrum and the left postzygapophysis slightly eroded, and lack ing the right postzygapophysis. The strongly con vex condyle suggests that this element was pro coelous. The notably elongated and compressed centrum, which is tubular in cross section at its mid-length and with a reduced neural spine are features present in cervical vertebrae IV to VI of azhdarchids (see Averianov, 2010), and conse quently, MPCA-PV 2003 is tentatively identified as a middle cervical element.
The neural arch is dorsoventrally low (Fig. 2D’). The neural spine is a very low and narrow ridge, and is confluent with the centrum at mid-length of the vertebra (Fig. 2A, C’). The neural canal is relatively small and subcircular in cross-section. Lateral to the neural canal there is a subcircular, much smaller, pneumatic foramen (Fig. 2D’). The postzygapophysis is relatively robust and ovoidal in cross-section, being sepa rated from the neural spine by a concave dorsal surface. In posterior view, the preserved postzy gapophysis has a deep ventral groove near its base, resulting in a ventrally pendant lateral end (Fig. 2D’). In lateral view, the postzygapophysis is located at the posterior end of a lateral ridge that gets dorsoventrally thicker posteriorly.
The condyle projects posterior to the neural arch and postzygapophyses (Fig. 2A’). In poste rior view, this articular surface is transversely broad and notably dorsoventrally flattened. It is higher than the neural canal and has convex dorsal and ventral margins (Fig. 2D’). In dorsal view, the posterior condyle has a convex articu lar surface, extending onto the lateral surfaces of the centrum on either side (Fig. 2A’). In poste rior view the condyle has an even, convex profile, which slopes ventrally towards the postexapophy ses (Fig. 2D’). The postexapophyses are tongue-shaped and posterolaterally directed (Fig. 2A’, B’). These are located at the posteroventral cor ner of the vertebra and are posteriorly separated from the articular condyle by a shallow groove. The ventral surface of the postexapophyses is flat, while the dorsal one is smoothly convex (Fig. 2D’). The lateral surface of the centrum is shallowly concave (Fig. 2C’), as also the ventral surface between the postexapophyses (Fig. 2B’). Anteriorly, the ventral surface of the centrum be comes straight close to the mid-length of the ele ment. This results in a tubular-shaped centrum at the mid-length of the element. The centrum lacks any foramen on its lateral surface (Fig. 2C’).
The preserved width of MPCA-PV 2003, mea sured across the postexapophyses, is 16.9 mm. In Quetzalcoatlus (specimen TMM 42889-1) the com parable measurement reaches a maximum of ap proximately 70 mm, whereas in Arambourgiania it reaches 90 mm (Frey & Martill, 1996). This indicates that the specimen here described be longed to a relatively small azhdarchid.
DISCUSSION
In spite of its incomplete nature, MPCA-PV 2003, may be nested among azhdarchid ptero saurs. Azhdarchids exhibit notably modified cervical vertebrae that show a unique combi nation of characters absent in other flying rep tiles, which were recognized as unique since the recognition of the clade in 1984 by Lev Nessov. MPCA-PV 2003 is referred to Azhdarchidae because of the occurrence of very elongate and transversely narrow vertebrae with well-de veloped postexapophyses, notoriously reduced neural spine which disappear at mid-length of the centrum, and centrum that is tubular in cross-section at mid-length (Nessov, 1984, 1991; Company et al., 1999; Pereda-Suberbiola et al., 2003; Osi & Weishampel, 2005; Averianov, 2014; Andres & Langston, 2021; Longrich et al., 2018) (Fig. 2). The lack of well-developed neural spine in MPCA-PV 2003 is a feature shared with azh darchids, such as Azhdarcho, Arambourgiania and Quetzalcoatlus. This differs from basal azh darchoids that retained well-developed neural spines as is the case of the African Cenomanian taxon Alanqa (Rodrigues et al., 2011). MPCA-PV 2003 lacks the deep ventral fossa and com pressed posterior half of vertebral centrum pres ent in Quetzalcoatlus (Andres & Langston, 2021 Longrich et al., 2018).
MPCA-PV 2003 is very fragmentary, and thus, a referral to a genus or species is not warranted. Furthermore, direct comparisons with other South American azhdarchids (i.e., Aerotitan, Thanatosdrakon) are not possible because of the lack of overlapping material. Thanatosdrakon preserves the posterior end of a posterior cervical vertebra, which has a higher and more complex neural arch, with dorsoventrally higher neu ral spine and a very large pneumatic foramina lateral to the neural canal, unlike the specimen here studied (Ortiz-David et al., 2022). It should be pointed out that MPCA-PV 2003 belongs to a species much smaller than Aerotitan and Thanatosdrakon, as inferred by its vertebral width.
The record of pterosaurs in the Cretaceous of Patagonia is very patchy and incomplete (Bonaparte, 1996; Kellner et al., 2003, 2006; Codorniú & Gasparini, 2007; Martinelli et al., 2007; Ibiricu et al., 2012; Novas et al., 2012; Kellner & Calvo, 2017; Bellardini & Codorniú, 2019; Díaz-Martínez et al., 2022). For the Cenomanian age (the earliest age of the Upper Cretaceous), pterosaurs are represented by still unpublished remains from the “La Buitrera” locality in Río Negro province (Haluza et al., 2007). Here we expand this patchy record with the first description of a pterosaur coming from Cenomanian beds of Patagonia. Furthermore, it constitutes the oldest record for azhdarchids in the South American continent, which were so far represented by the Coniacian-Santonian Thanatosdrakon amaru (Ortiz-David et al., 2022) and the Maastrichtian Aerotitan sudamericanus (Novas et al., 2012).
In sum, this finding constitutes an important addition to the knowledge of the poorly known pterosaur faunas from completely continental assemblages of the Upper Cretaceous of South America, and also indicates that the diversifica tion and evolution of the clade in South America is still poorly known.

