











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkLos impactos antrópicos, tales como la pérdida y/o fragmentación de hábitat, la introducción de especies exóticas y/o la defaunación, son considerados una gran amenaza para la biodiversidad y el funcio namiento de los ecosistemas. La biodiversidad ha sido tradicionalmente medida como la riqueza de especies presentes en un lugar; sin embargo, esta necesita ser evaluada desde una perspectiva comunitaria y a través de componentes más estrechamente ligados a las funciones ecosistémicas per se (Donoso Cuadrado 2017). Conocer la diversidad o la riqueza de especies no es equivalente a entender el funcionamiento del ecosistema; es decir, qué hace cada especie y cómo se asocian entre sí para mantener el ecosistema funcionando (del Val de Gortari 2022). En los últimos años las redes ecológicas, es decir, los conjuntos de especies vinculadas por interacciones ecológicas, son consideradas una herramienta más apropiada para explicar la dinámica de las comunidades y la estabilidad de los ecosistemas (Montoya et al. 2006, Donoso Cuadrado 2017). Es decir, conocer y entender dichas interacciones es fundamental en el ámbito de la biología de la conservación, ya que el funcionamiento de los ecosistemas depende de dichas interacciones y no de la riqueza de especies (Montoya et al. 2006, Bascompte y Jordano 2007, 2008, del Val de Gortari 2022).
Las redes de interacción mutualista formada por las plantas y los animales son componentes de gran importancia en numerosos ecosistemas. Por ejemplo, en los ambientes tropicales el 50% de las aves consumen frutos al menos durante alguna parte del año (Stiles 1985, Loiselle y Blake 1990) y el 90% de las especies arbóreas y el 98% de los arbustos del soto-bosque son dispersados por animales (Gentry 1982, Howe y Smallwood 1982, Stiles 1985). Estas interacciones mutualistas con beneficios recíprocos conforman la arquitectura de la biodiversidad (Montoya et al. 2006, Bascompte y Jordano 2007). Estudiar dichas interacciones frugívoras, en su sentido amplio de red, es fundamental para conocer y evaluar la respuesta de los ecosistemas frente a diferentes perturbaciones.
En Argentina el bosque xerófilo del Espinal ha sido fuertemente destruido por el avance de la frontera agrícola. Debido a ello en la actualidad solo se conservan algunos relictos con vegetación nativa en medio de ambientes antropizados y expuestos a un gran número de presiones como el sobrepastoreo y las quemas descontroladas (Rainer Cinti 1998). A pesar de ello, aún se conservan relictos de dicho bosque los cuales albergan una importante riqueza de plantas nativas, con al menos 9 especies endémicas (Arana et al. 2021), y una amplia variedad de especies de aves (Morello et al. 2012, Verga et al. 2019). A día de hoy no existen estudios científicos relacionando las redes de interacción formadas por las plantas y las aves del bosque del Espinal; debido a ello, el presente trabajo tuvo por objetivos: a) ampliar el conocimiento de las especies de aves frugívoras/omnívoras presentes en un bosque del espinal, b) determinar qué especies de semillas están siendo dispersadas por las aves a través de las fecas, c) conocer si la dispersión es legítima; es decir si las semillas se encuentren sanas/sin daños físicos (Bustamante et al. 1992, Bustamante y Canals 1995) y d) describir la conectividad y el grado de ani-damiento de la red de interacción generada.
MÉTODOS
Área de estudioEl presente estudio se llevó a cabo en el área protegida "Bosque Autóctono El Espinal” ubicada en el campus de la Universidad Nacional de Río Cuarto a los 33°10’S y 64°20’O. Esta área protegida fue creada en 2002 y posee una superficie de 7 ha.
La temperatura máxima media anual es de 20°C en el mes de enero y la mínima media anual es de 3°C en el mes de julio. Las precipitaciones máximas ocurren en los meses de diciembre y enero (130 mm) y las mínimas en los meses de junio y julio (13 mm).
La formación vegetal que caracteriza a este relicto, representativo de la provincia fitogeográfica del Espinal, es el bosque xerófilo abierto, donde domina un estrato arbóreo con especies leñosas como "Chañar” (Geoffroea decorticans), "Tala” (Celtis ehrenbergiana) "Moradillo” (Schinus fasciculatus) junto con especies exóticas como la "Mora” (Morus nigra) y el "Olmo” (Ul-mus pumila). En el estrato arbustivo se encuentran "Chilcas” (Baccharis salicifolia), "Palo Amarillo” (Aloy-sia gratissima) y "Poleo” (Lippia turbinata) entre otras. Acompañan a las especies leñosas distintas herbáceas, en su mayoría compuestas y gramíneas; de las 198 especies catalogadas el 76.26% son autóctonas y 23.73% introducidas (Vischi y Oggero 2002).
Actualmente el Bosque Autóctono El Espinal cuenta con 126 especies de aves registradas pertenecientes a 31 familias diferentes (eBird 2022) y únicamente dos de ellas son especies exóticas: la Paloma Doméstica (Columba livia) y el Estornino Pinto (Sturnus vulgaris). Del total del ensamble de aves, aproximadamente el 55% son insectívoras, el 17% son omnívoras, el 10% son granívoras y el 2% son frugívoras (el porcentaje restante corresponde a otros tipos de dietas más especializadas) (de la Peña 2016, Tobias 2021).
Muestreo de aves
Se realizaron muestreos mensuales de aves desde diciembre de 2018 hasta diciembre de 2019 con redes de niebla (malla de 36 mm, 12 x 2.5 m) abiertas generalmente entre las 06:00 y las 13:00hs en el interior y en el borde del Bosque Autóctono El Espinal. Durante los muestreos se colocaron de cuatro a seis redes de niebla separadas a una distancia de entre 70 y 100 metros. Las posiciones de las redes de niebla se seleccionaron estratégicamente para facilitar el muestreo de aves en todos los tipos de vegetación representativos y en sus diferentes combinaciones. Las aves capturadas fueron anilladas con un anillo de aluminio y el procesamiento como la toma de datos de los ejemplares se realizó siguiendo las instrucciones Ralph et al. (1996) y las normas internacionales propuestas por el North American Banding Council (NABC 2001 a,b). Se tomaron medidas morfométricas estándar de todas las aves capturadas; la edad y el sexo se asignaron siguiendo los criterios de Pyle (1997).
Caracterización de la dieta de las aves y análisis de semillasEl estudio de la dieta se realizó mediante la recolección de fecas. Para ello las aves capturadas en las redes se mantuvieron en sacos de tela hasta obtener la muestra - lo cual sucede generalmente en menos de 10 min (Brandolin obs. pers.) - garantizando así la procedencia (especie) de cada muestra (Hernandez Ladrón et al. 2012). Las fecas recolectadas fueron colocadas en tubos eppendorf con la descripción del lugar, fecha y posición geográfica. Posteriormente, en el laboratorio fueron suavemente desarmadas, con la ayuda de una pinza diente de león, y se analizaron bajo lupa estereoscópica con el objetivo de identificar los distintos ítems presentes. Las semillas fueron separadas y se identificaron a nivel específico cuando fue posible. La identificación específica de las semillas se realizó con la ayuda de un catálogo de referencia elaborado previamente, mediante la ayuda de expertos y de bibliografía específica (Bianco et al. 2000, Zubizarreta y Díaz Panizza 2014). Además, dichas semillas se revisaron en detalle para observar si presentaban algún daño físico (quiebres, exposición del embrión, vaciamiento, etc.).
Análisis de la red de interacción frugívoraSe calculó el número de especies de plantas (P), el número de aves frugívoras (A) y el tamaño de la red (S). Seguidamente se determinaron las interacciones potenciales (AxP) y el número observado de interacciones (L). Las interacciones se dan entre pares de especies de cada conjunto de nodos y representan un vínculo que implica una reciprocidad. Estos datos fueron usados para describir la conectividad con dos parámetros: conectancia y densidad de la red. La conec-tancia, C= L/(AxP), se interpreta como la proporción de interacciones registradas del total de interacciones potenciales. La densidad de la red L/(A+P), como la media del número de interacciones por especie.
El grado de anidamiento se calculó usando el algoritmo NODF propuesto por Almeida-Neto et al. (2008) implementado en el programa ANINHADO 3.0 (Guimaraes y Guimaraes 2006). Los valores de NODF oscilan entre 0 y 100, donde N toma valores próximos a 100 en matrices muy anidadas. La significancia de NODF se estimó usando un procedimiento de Monte Carlo. El grado de anidamiento obtenido se comparó con un modelo nulo (Ce en ANINHADO) generando 1000 matrices aleatorias del mismo tamaño que la registrada, donde la probabilidad de interacción entre una planta y un frugívoro es proporcional a su número total de interacciones (Bascompte et al. 2003). El valor de P se definió como la fracción de matrices aleatorias que tuvieron un NODF igual o mayor que la matriz real. Cuando ninguna matriz tenía un valor mayor de NODF que la matriz real, se definió p<0.001(Mello et al. 2011).
RESULTADOS
Muestreo de avesSe capturaron 58 ejemplares de aves pertenecientes a 31 especies diferentes, de las cuales en 14 de ellas se ha observado que incluyen frutos/semillas en su dieta: Tordo Músico (Agelaioides badius) (n= 1), Tacuarita Azul (Polioptila dumicola) (n= 2), Sietevesti-dos Serrano (Poospiza whitii) (n= 2), Torcacita Picuí (Columbina picui) (n= 1), Juan Chiviro (Cyclarhis gujanensis) (n= 1), Fiofío Silbón (Elaenia albiceps) (n= 6), Fiofío Pico Corto (Elaenia parvirostris) (n= 3), Gorrión (Passer do-mesticus) (n= 1), Corbatita (Sporophila caerulescens) (n= 4), Pijuí Frente Grís (Synallaxis frontalis) (n= 1), Ratona Común (Troglodytes aedon) (n= 9), Zorzal Chalchalero (Turdus amaurochalinus) (n= 4), Zorzal Colorado (Tur-dus rufiventris) (n= 2) y Chingolo (Zonotrichia capensis) (n= 8).
Caracterización de la dieta y análisis de semillasDel total de muestras de fecas analizadas (n= 58) el 78% (n= 45) fueron de las especies consumidoras de frutos y semillas y el 21% de ellas contenía semillas. Se contabilizaron 90 semillas pertenecientes a 10 familias y 12 especies diferentes de las cuales cinco fueron especies autóctonas: Galium sp., Solanumpseu-docapsicum, Celtis erherbergiana, Poligonum acuminatum y Stipa sp.; cinco exóticas: Chenopodium álbum, Morus nigra, Leonurus sibiricus, Medicago sp., Anagalis sp. y dos especies no fueron identificadas. El 90% de las semillas no presentaron ningún tipo de daños físicos luego de pasar por el tracto digestivo de las aves.
Las muestras de fecas con semillas pertenecieron a 6 especies (4 familias) de aves: Torcacita Picuí, Fio-fío Silbón, Fiofío Pico Corto, Piojito Trinador, Zorzal Chalchalero y Tordo Músico.
Análisis de la red de interacción frugívoraA nivel de red se observó un total de 6 especies de aves (A) que consumieron frutos de 12 especies de plantas (P) para un tamaño de red de 18 especies (S). El número de interacciones potenciales (AxP) fue de 72 y el número de interacciones observada (L) fue de 16 para una conectancia (C) del 22% (Tabla 1). La densidad de la red fue estimada en 0.88 vínculos por especie.
Las plantas registradas pertenecen a 10 familias botánicas de las cuales 2 fueron árboles, uno autóctono (C. erherbergiana) y uno exótico (M. nigra), y 8 corresponden a especies herbáceas, 4 exóticas (C. álbum, L. sibiricus, Medicago sp., Anagallis sp.) y 4 autóctonas (Ga-lium sp., S. pseudocapsicum, Stipa sp. y Polygonum acumi-natum) (Tabla 1).
El grado de anidamiento de la red de frugivoría en el bosque del espinal fue de NODF= 28.09, P<0.01 con un mayor grado de anidamiento en el caso de las aves NODF= 30.3, P<0.01 que para las plantas NODF= 18.33; P<0.08 (Fig. 1).
DISCUSIÓN Y CONCLUSIONES
Este trabajo confirma que las aves realizan una importante dispersión de semillas a través de las fe-cas ya que la mayoría de ellas no presentaron daños físicos luego de pasar por el tracto digestivo de las mismas. Las semillas dispersadas corresponden tanto a especies nativas como a especies exóticas que están presentes en el área de estudio. Algunas de las especies observadas (C. erherbergiana y S. pseudocap-sicum) ya han sido citadas como dispersadas por las aves (Dellafiore 2016) y varias de ellas germinan luego de ser defecadas como es el caso de C. erherbergiana (Dellafiore y Rosa 2017), S. pseudocapsicum, M. nigra y C. album (Dellafiore obs.pers.).
Las especies de plantas que presentaron el mayor número de interacciones con las aves se corresponden a especies con una elevada producción de frutos/ semillas (Medicago sp., M. nigra y P acuminatum) las cuales han sido mencionadas como parte de la dieta de las aves y/o han sido mencionadas como especies dispersadas por ellas (Aramburu 1997, Mancina 2006, Góes-Silva et al. 2012).
Tabla 1: Número de interacciones (las interacciones se dan entre pares de especies de cada conjunto de nodos y representan un vínculo que implica reciprocidad) para las distintas especies de aves y de plantas. En negrita las especies autóctonas.
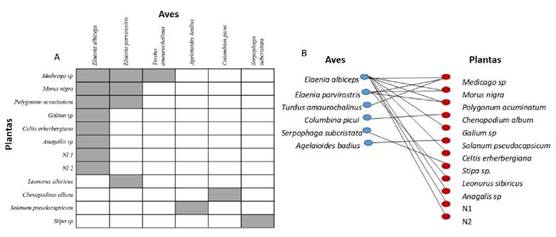
Figura 1: A: Matriz de interacciones frugívoras. Cada cuadro representa una interacción potencial entre un ave y una planta ornitócora, donde los cuadros con color rojo representan una interacción registrada. Las filas y columnas se organizaron con las especies siguiendo un orden de mayor a menor número de interacciones para facilitar la visualización del anidamiento. B: Gráfica bipartita con las aves de color azul y las plantas de color rojo.
Las especies de aves con mayor número de interacciones fueron Fiofío Silbón y Fiofío Pico Corto, las cuales son migrantes del sistema migratorio neotro-pical austral (Jimenez et al. 2016, Hosner 2020), y pasan el invierno en latitudes tropicales de Sudamérica. Para reproducirse el Fiofío Silbón migra a un estrecho tramo de bosque a lo largo de la Cordillera de los Andes, entre el norte de la Patagonia y el extremo sur de Sudamérica (Capllonch et al. 2011). Por el contrario, el Fiofío Pico Corto se reproduce desde Bolivia hasta el centro de Argentina y Uruguay (Hosner 2020). Durante el verano ambas especies son frugívoras y muy abundantes representando más del 20% de las aves capturadas durante los periodos de fructificación de las plantas en la Reserva del Bosque Autóctono del Espinal y llegando a superar el 80% en algunos sitios (ver Bravo et al. 2017). Las especies de fiofíos serían unas de las principales dispersoras de semillas en el bosque del espinal y, además, debido a su abundancia y al espectro de semillas dispersadas podrían jugar un papel clave en la dinámica de las especies que dispersa. Más estudios son necesarios para evaluar si las semillas dispersadas conservan su poder germinativo, luego de pasar por el tracto digestivo, y si posteriormente pueden establecerse efectivamente.
El Zorzal Colorado a pesar de ser considerada una especie principalmente frugívora, no presentó semillas en sus fecas encontrándose exclusivamente insectos. En cambio, en el Piojito Trinador, que ha sido caracterizado principalmente como una especie insectívora de follaje (Milesi et al. 2002), encontramos en uno de los ejemplares cinco semillas de Stipa sp. En sus fecas. Esta ingesta, aunque probablemente sea accidental, puede resultar de importancia para la dispersión de dicha especie ya que las semillas fueron encontradas sin daños físicos.
Analizando los patrones de la red de interacción generada observamos que en el bosque del espinal la conectividad resulta ser bastante baja con 0.88 vínculos por especie, pero la conectancia fue elevada (22%) teniendo en cuenta los valores reportados en la literatura (10-19%) (Albert et al. 2000, Palacios Arce 2014). La conectancia es una medida de comunidad básica del número de interacciones y cuando su valor es más grande se incrementa la robustez de la red (Staniczenko et al. 2013, Palacios Arce 2014, Poisot y Gravel 2014). Para las especies individuales, el número de interacciones es un excelente predictor de su persistencia en la comunidad (James et al. 2012). Los estudios de redes muestran que las especies más vulnerables tienen menos interacciones, y a su vez que un ataque dirigido a las especies más conectadas pone en riesgo a la red entera (Memmot et al. 2004).
La red de frugivoría en el bosque del espinal posee valores de anidamiento algo más bajo (NODF= 28.09) que los citados en la literatura para bosques nublados (NODF= 31-46) (Bascompte et al. 2003, Palacios Arce 2014) lo que indica que no hay un grupo de especies generalistas con más interacciones que interactúan con un subconjunto de las especies de plantas con menos interacciones. Sin embargo, se observa un mayor grado de anidamiento en el caso de las aves (NODF= 30.3, P<0.01) que para las plantas (NODF= 18.33; P<0.08); es decir, que se observa que algunas aves son más generalistas e interactúan con un subconjunto de especies de plantas con menos interacciones.
Una red de interacción representa las conexiones y dependencias entre especies, y su estudio es fundamental para comprender cómo estas interacciones sustentan la biodiversidad y funcionalidad de ecosistemas (Jordano 2009). Hasta el momento se desconoce el ensamble total de especies que pueden estar presentes en este relicto de Espinal, por lo que la red de interacción descripta en este trabajo podría ser limitada y no reflejar la totalidad de las relaciones que pudieran existir. Por otro lado, sólo contamos con las semillas colectadas en una feca, con lo cual no necesariamente sea representativo de lo que come esa especie a lo largo del tiempo. De todas maneras, la red presentada nos permitió conocer cómo está conformada la comunidad de aves dispersoras de semillas en el bosque del espinal e identificar especies de aves insectívoras que podrían también dispersar semillas, aunque el consumo de las mismas pueda ser accidental. Una semilla dispersada sana y que luego germina resulta ser una interacción positiva para la planta dispersada. La información presentada en este trabajo nos permite tener una visión más completa de la dinámica ecológica y la estructura de los ecosistemas, y cómo estas interacciones pueden verse afectadas por factores como la pérdida de hábitat o el cambio climático (Uribe Botero 2015).
La continuidad de este tipo de estudios aumentará sin duda la representatividad y nos permitirán conocer el funcionamiento del ecosistema e implementar medidas de conservación más realistas; es decir, basadas en la dinámica del ecosistema y no en especies individuales. Este enfoque permitirá obtener información adicional sobre las especies de aves y plantas presentes en el ecosistema, así como las interacciones que mantienen entre sí, sus cambios en el tiempo y respuestas a las perturbaciones.
AGRADECIMIENTOS
Agradecemos a los revisores anónimos y a la Dr. Susana Peluc por los aportes y sugerencias realizados los cuales ayudaron a mejorar el presente trabajo. Agradecemos a la Secretaría de Ciencia y Técnica de la Universidad Nacional de Río Cuarto por el apoyo financiero.
Recibido: 27 de febrero 2023
Aceptado: 17 de julio 2023

