











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
Las revisiones taxonómicas y las diferentes fuentes de información biológica (catálogos, inventarios, bases de datos), son esenciales y constituyen los insumos básicos para entender los patrones biogeográficos y la organización espacial de las comunidades vegetales (Funk & Richardson, 2002; Funk, 2006; Borges et al., 2010). En este sentido, la búsqueda de distintos tipos de patrones es un tema central y recurrente en biogeografía y ecología (Ferro & Barquez, 2014). La visión tradicional de una disminución lineal del número de especies con la altitud fue interpretada alguna vez como un patrón universal, semejante al observado en los gradientes latitudinales (Ferro & Barquez, 2014). Sin embargo, estudios recientes muestran que no existe un único patrón altitudinal de riqueza de especies, y que contrariamente al supuesto de una disminución monotónica en función de la altitud, el caso más común es un pico de riqueza en altitudes intermedias (Ferro & Barquez, 2014), referido como el “efecto del dominio medio” (Colwell & Lees, 2000; Adams, 2009; Giraldo-Cañas, 2014).
En este contexto, el conocimiento detallado de la variación altitudinal de la riqueza de especies en diferentes regiones del mundo es un requisito fundamental a la hora de indagar los procesos que producen dichos patrones (Colwell & Lees, 2000; Grytnes & Vetaas, 2002; Adams, 2009; Borges et al., 2010; Ferro & Barquez, 2014). Por lo tanto, los gradientes altitudinales proveen un importante escenario para estudiar las relaciones de la distribución de especies y el clima, debido a la gran variación climática en pequeñas distancias geográficas (Lloret & González-Mancebo, 2011; Giraldo-Cañas, 2014). En este sentido y dadas la riqueza y la amplia expresión ambiental de sus especies, la familia Malpighiaceae Juss. sería un modelo para estudiar procesos de distribución en el neotrópico.
Las Malpighiaceae constituyen una familia de 77 géneros y ca. 1300 especies (Almeida et al., 2016; Davis et al., 2020a), distribuidas en regiones tropicales y subtropicales de ambos hemisferios, con cerca del 90% de las especies presentes en el Nuevo Mundo (desde Arizona, Nuevo México, Texas y Florida en los Estados Unidos de América, el Caribe hasta la Argentina y Chile) y el restante 10%, se encuentra en el Viejo Mundo (África, Australia, China, Filipinas, India, Indonesia, Madagascar, Malasia, Micronesia, Nepal, Nueva Caledonia, Nueva Guinea, Pakistán, Península Arábiga, Sri Lanka, Taiwán, Tailandia y varias islas del Pacífico) (Anderson et al., 2020). Al respecto, las Malpighiaceae se dispersaron desde Sudamérica vía el Caribe hacia Norteamérica y de ahí a Eurasia por el Atlántico norte y subsecuentemente, alcanzaron diferentes áreas tropicales del Viejo Mundo, cuando las condiciones paleoambientales así lo facilitaron (Davis & Anderson, 2010).
La familia Malpighiaceae se caracteriza por variados hábitos de crecimiento, desde árboles, arbustos, sufrútices y muy raramente hierbas perennes en Aspicarpa Rich. (Anderson, 2016), hasta especies trepadoras, tanto leñosas como herbáceas, las cuales se distribuyen en una amplia gama de hábitats. Cabe destacar que las Malpighiaceae están entre las familias con más diversidad de especies trepadoras en el neotrópico, junto con las Apocynaceae, las Asteraceae, las Bignoniaceae, las Fabaceae, las Passifloraceae y las Sapindaceae, entre otras (Linares, 2001; Prósperi et al., 2001; Acevedo-Rodríguez, 2005; Abadía Bonilla et al.,
2015). En este contexto, el objetivo de este trabajo es documentar los patrones de variación altitudinal en la riqueza, el endemismo y la diversidad filogenética de las Malpighiaceae en Colombia.
MATERIALES Y MÉTODOS
La circunscripción y la nomenclatura de los géneros y las especies está basada en Anderson (2006; 2013), Anderson & Davis (2007; 2013), Davis & Anderson (2010), Anderson & Anderson (2016), Anderson et al. (2020), Davis et al. (2020b) y Almeida & van den Berg (2021). Los datos de la representatividad de la familia Malpighiaceae en Colombia provienen de muestreos personales, de la revisión de las colecciones depositadas en los herbarios ACAM, AFP, ANDES, AS, BA, BAF, BRG, CAUP, CDMB, CEN, CEPEC, COAH, COL, CR, CUVC, CHOCO, EAN, F, FAUC, FMB, G, HECASA, HFAB, HORI, HPUJ, HUA, HUC, HUQ, IAN, IBGE, INPA, IPA, JAUM, LLANOS, MEDEL, MO, NY, PSO, QCA, RSA, SI, SURCO, TOLI, UDBC, UIS, UPTC, US y VALLE (Thiers, 2020), así como de Cuatrecasas (1958), Giraldo-Cañas (1996; 2011) y Anderson & Anderson (2016). A los ejemplares de herbario se les tomó toda la información de sus respectivas fichas, tales como departamento, municipio, localidad, altitud, coordenadas (aunque en la mayoría de los ejemplares de herbario este dato no figuraba), usos, nombres populares, estado reproductivo, fechas de colección, recolectores, números de colección y herbario. A la localidad de los ejemplares analizados se la clasificó por unidad biogeográfica en el contexto colombiano. Las unidades biogeográficas colombianas están basadas en Bernal et al. (2016). La consideración de las especies nuevas está basada en Anderson & Anderson (2016). Por su parte, los términos riqueza/diversidad y endemismo están basados en Badii et al. (2007) y Noguera-Urbano (2017), respectivamente. La diversidad filogenética se basa en los postulados de Llopis-Belenguer et al. (2018) y Hernández-Ruedas et al. (2019) y aquí se hace una mirada desde la representatividad de los clados de la familia en Colombia. La configuración y los nombres de los clados de las Malpighiaceae está basada en Anderson et al. (2020). Se utilizaron intervalos altitudinales con una amplitud de 500 m, amplitud frecuentemente utilizada en estudios de gradientes altitudinales (e.g. Stevens, 1992; Lægaard, 1999; Etter & van Wyngaarden, 2000; Grytnes & Vetaas, 2002; Kessler, 2002; Oommen & Shanker, 2005; Giraldo-Cañas, 2014; 2018), lo cual facilita las comparaciones de riqueza, diversidad y endemismo entre diferentes grupos biológicos. Adicionalmente, con el fin de identificar el patrón de variación con la altitud para la riqueza de especies y el endemismo, se usan los números de riqueza o endemismo por cada intervalo altitudinal, a partir de un análisis de regresión lineal simple (Daniel, 1990).
RESULTADOS Y DISCUSIÓN
Riqueza, endemismo y distribución. Las Malpighiaceae están representadas en Colombia por 34 géneros y 246 especies (Tablas 1 y 2); así, en Colombia está representado el 19% de las especies de la familia a nivel mundial y el 44,2% de los géneros. Cabe destacar que Cuatrecasas (1958) halló para Colombia 21 géneros y 168 especies, así, en los 63 años entre ese trabajo y el actual, se ha incrementado en 78 el número de especies de Malpighiaceae para la flora de Colombia. El endemismo corresponde a 47 especies, lo cual constituye el 19,1% de las especies presentes en Colombia. Así, Colombia es uno de los países más ricos en esta familia junto con Brasil (584 especies), Venezuela (189), México (169) y Perú (137) (Mamede et al., 2010; Villaseñor, 2016; Ulloa Ulloa et al., 2017; Almeida et al., 2021). Por su parte, los géneros más diversificados en Colombia son Byrsonima Kunth (36 especies), Heteropterys Kunth (28), Hiraea Jacq. (28), Stigmaphyllon A. Juss. (28), Tetrapterys Cav. (25), Bunchosia Kunth (23) y Mascagnia (DC.) Bertero (15). Este patrón de riqueza por género se ajusta, en líneas generales, a los patrones encontrados en otros países con numerosas Malpighiaceae (Mamede et al., 2010; Villaseñor, 2016; Ulloa Ulloa et al., 2017). En cuanto al hábito, las formas lianescentes suman 162 especies, los árboles y los arbustos representan 83 especies, y sólo hay en Colombia una especie correspondiente a un sufrútice, Byrsonima verbascifolia (Tabla 2).
La mayor riqueza de especies se da en las tierras bajas; así, en el primer intervalo altitudinal (0-500 m s.m.) se han encontrado 212 especies, lo cual representa el 86,2% de las especies presentes en Colombia, seguido del segundo intervalo (500-1000 m s.m.) con 94 especies (38,2%) (Tabla 2), mientras que el límite altitudinal de las Malpighiaceae en Colombia (3200 m s.m.) sólo lo alcanzan dos especies, Stigmaphyllon bogotense, en los Andes y la Sierra Nevada de Santa Marta, y Stigmaphyllon columbicum, en los Andes.
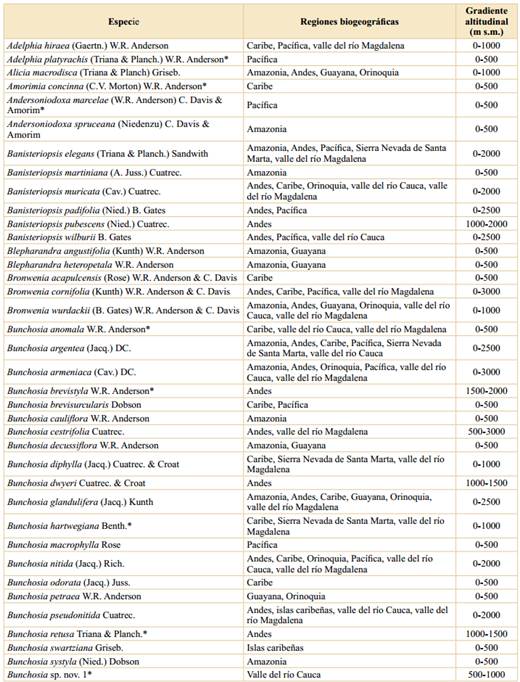
Tabla 1 Las Malpighiaceae de Colombia y su distribución por regiones biogeográficas y gradientes altitudinales.
*: Especie endémica de Colombia.
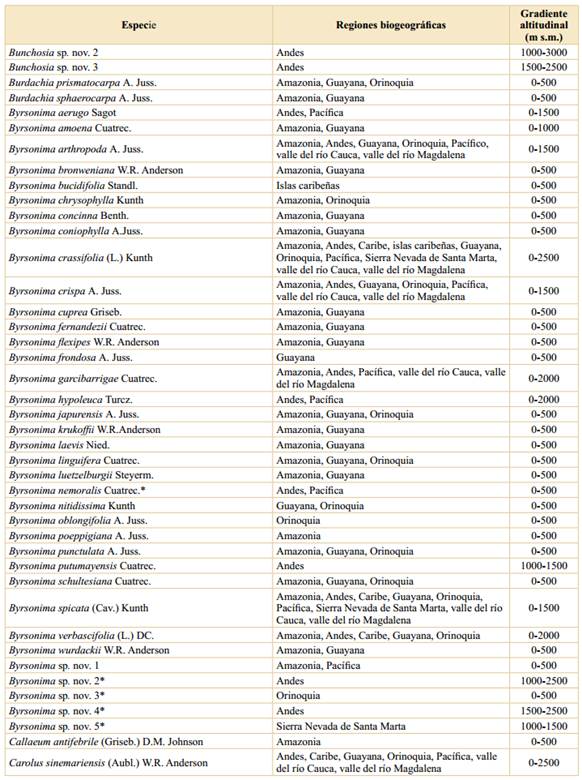

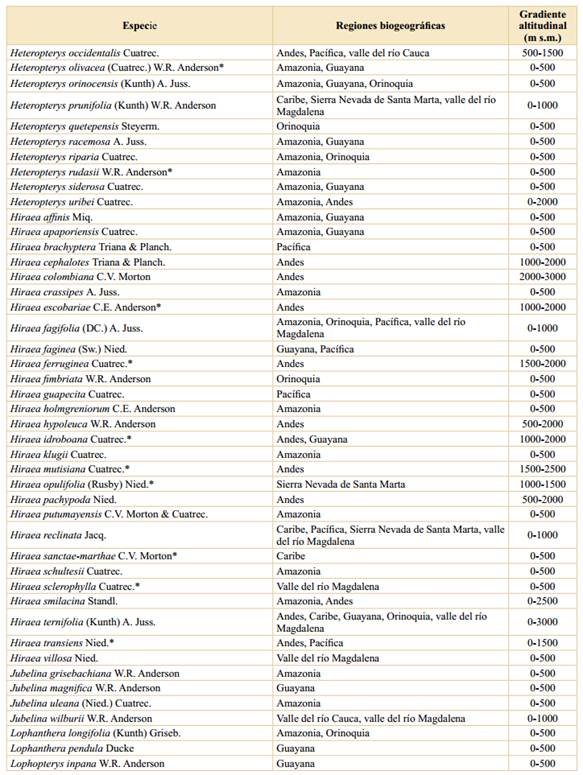
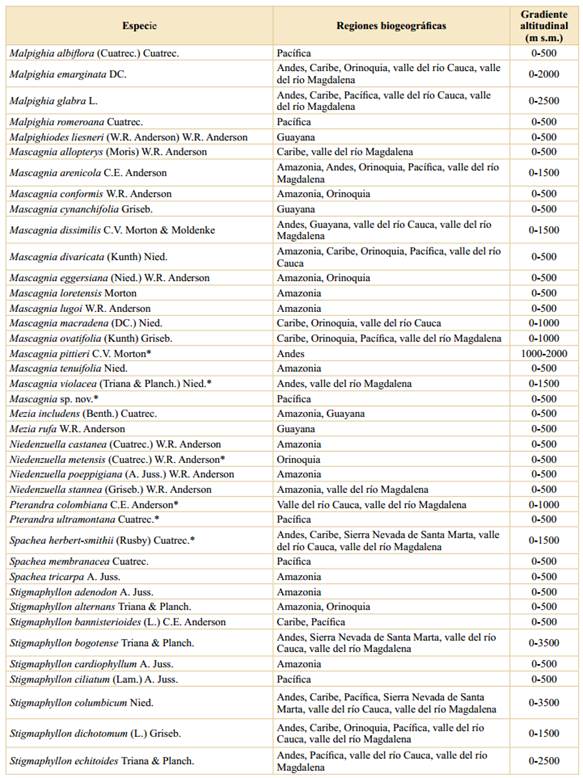
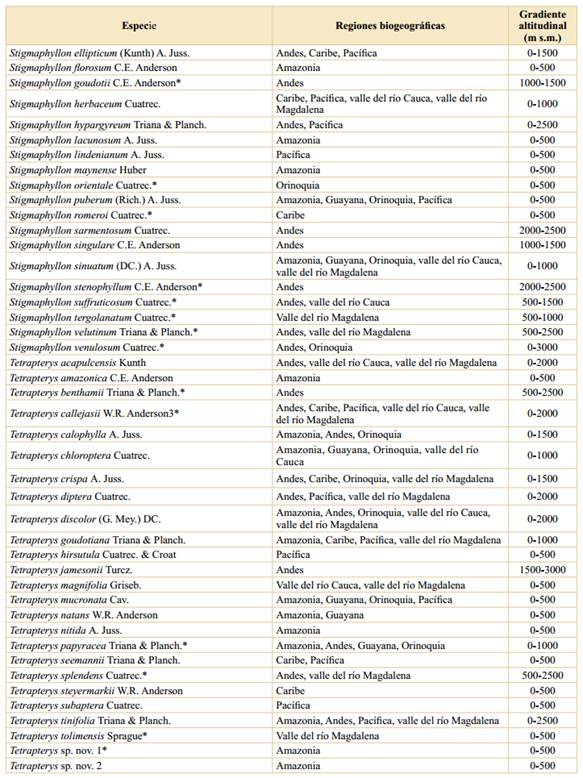
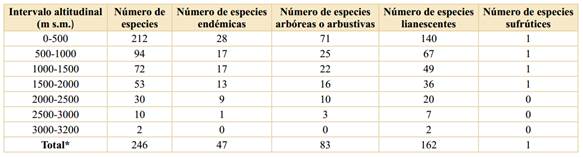
Tabla 2 Riqueza y endemismo de las Malpighiaceae en Colombia, sobre la base de gradientes altitudinales. *: La suma de las columnas no es aritmética, ya que hay varias especies que se distribuyen en más de un intervalo altitudinal.
Asimismo, el número de especies endémicas por intervalo de altitud muestra el mismo patrón, en donde las tierras bajas (0-500 m s.m.) son las que presentan el mayor número de especies endémicas (28), seguido del segundo intervalo altitudinal con 17 especies y el endemismo va disminuyendo a medida que se asciende en el gradiente altitudinal (Tabla 2). Estos datos se ajustan a los encontrados para otras familias vegetales en Colombia (Rangel-Ch., 2006; 2015; Giraldo-Cañas, 2010; 2018) estudiadas en un contexto de gradientes altitudinales, así como en Ecuador (Lægaard, 1999; Kessler, 2002) y Venezuela (Giraldo-Cañas, 2014), en las cuales las tierras bajas y medias (0-1500 m s.m.) tienen la mayor riqueza, no sólo en número de especies sino también en géneros y clados (Giraldo-Cañas, 2014).
El patrón de riqueza altitudinal, así como el patrón de endemismo altitudinal, muestran una disminución con la altitud (y = -0,061x + 173,57; R2 = 0,82; y = -0,009x + 27,76; R2 = 0,95, respectivamente, donde “y” es el número de especies y “x” la altitud). Este patrón de riqueza de especies es exhibido por otros grupos vegetales en el neotrópico, tales como Poaceae (Giraldo-Cañas, 2010; 2014), Marcgraviaceae (Giraldo-Cañas, 2018), así como por la riqueza de especies en numerosos bosques neotropicales en diferentes gradientes altitudinales (Gentry, 1995). Por otra parte, los datos aquí analizados para las Malpighiaceae de Colombia, no se ajustan al “efecto del dominio medio” (Colwell & Lees, 2000; Adams, 2009; Giraldo-Cañas, 2014), el cual contempla que la mayor riqueza de especies, en un contexto de gradientes altitudinales, se encuentra en las alturas intermedias, como es el caso de la riqueza de Acanthaceae, Bromeliaceae, Pteridophyta, Solanaceae) (Kessler & Kluge, 2008; Adams, 2009) o el caso de las especies de la familia Frullaniaceae de las hepáticas en áreas montanas neotropicales (Gradstein & Uribe, 2011). Esta situación se ajusta a la naturaleza ecológica y biogeográfica de la mayoría de las especies de Malpighiaceae, las cuales prefieren los hábitats húmedos y cálidos de tierras bajas, principalmente de la cuenca amazónica, de variadas formaciones vegetales de los escudos precámbricos sudamericanos y de la Mata Atlántica (Anderson, 1981; 2001; Mamede et al., 2010; Rangel-Ch., 2015; Almeida et al., 2016).
Los límites altitudinales de la familia Malpighiaceae en Colombia, alcanzan los 3200 m s.m. y tan sólo diez especies se distribuyen en las tierras altas (2500-3200 m s.m.), como en algunos bosques andinos, tanto primarios como secundarios, arbustales, bordes de bosque y en zonas de transición entre bosques altoandinos y páramos, a saber, Bronwenia cornifolia (≤3000 m s.m.), Bunchosia armeniaca (≤2800 m s.m.), Bunchosia cestrifolia (≤2950 m s.m.), Bunchosia sp. nov. (≤2700 m s.m.), Hiraea colombiana (≤2800 m s.m.), Hiraea ternifolia (≤2750 m s.m.), Stigmaphyllon bogotense (≤3200 m s.m.), Stigmaphyllon columbicum (≤3200 m s.m.), Stigmaphyllon venulosum (≤2800 m s.m.) y Tetrapterys jamesonii (≤2600 m s.m.) (Tabla 1).
La Amazonia, los Andes y el valle del río Magdalena, exhiben la riqueza de especies más alta, con 116, 89 y 68, respectivamente (Figura 1, Tabla 3). Cabe resaltar que en la Sierra Nevada de Santa Marta (la montaña costera más alta del mundo, Alvear et al., 2015), sólo se han registrado 22 especies, pero claramente esta baja riqueza se debe a que dicha sierra está pobremente muestreada (Alvear et al., 2015), así que se cree que su real riqueza biológica y endemismo, no sólo de Malpighiaceae sino de muchos otros grupos de plantas, sería muy destacada (Alvear et al., 2015), y esto se basa en la “Hipótesis de la heterogeneidad ambiental”, la cual sustenta la idea de que ambientes heterogéneos permiten ser colonizados por un mayor número de especies, con lo cual aumentan la riqueza y la diversidad biológicas (Badii et al., 2007).
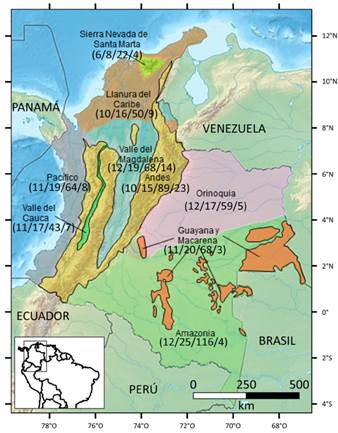
Fig. 1 Diversidad filogenética, riqueza genérica y específica y endemismo de las Malpighiaceae en las regiones biogeográficas de Colombia (número de clados/ número de géneros/número de especies/número de especies endémicas). En esta figura, por cuestiones de escala, no aparecen las islas caribeñas colombianas (2/2/4/0) (mapa modificado de Bernal et al., 2016). Figura en color en la versión en línea http://www.ojs.darwin.edu.ar/index.php/ darwiniana/article/view/923/1206
El mayor número de especies endémicas se encuentra en los Andes y en el valle del río Magdalena, con 23 y 14 especies, respectivamente (Figura 1, Tabla 3). Es curioso el bajo endemismo en la Guayana colombiana, toda vez que las Malpighiaceae presentan una riqueza y un endemismo muy destacados en toda esta región biogeográfica (Anderson, 1981), con 160 especies, 100 de las cuales son endémicas (Anderson, 1981; Giraldo-Cañas, 2001; 2008), y más concretamente, la Guayana venezolana presenta 153 especies, 32 de las cuales son endémicas (Anderson, 2001). Así, al igual que para la Sierra Nevada de Santa Marta, los inventarios vegetales son escasos y fragmentarios en la Guayana colombiana (así como en otras áreas colombianas, e. g. Amazonia, Pacífica y varios valles intra-andinos, entre otras), por lo tanto, estos datos actuales de riqueza y endemismo, no mostraría su real expresión en esta área. Adicionalmente, las formaciones vegetales de la Guayana se asientan en sustratos muy antiguos, ya que dicha región es de origen precámbrico (véanse Giraldo-Cañas, 2001; 2008) y así, en este contexto se podría recurrir a la “Hipótesis del Tiempo” para esperar una mayor riqueza y diversidad de especies en la Guayana colombiana, dado que esta hipótesis relaciona antigüedad geológica con una mayor riqueza y diversidad biológicas (Badii et al., 2007).
Disyunciones anfiatlánticas. De las especies presentes en Colombia, Heteropterys leona y Stigmaphyllon bannisterioides exhiben una distribución natural en América tropical y en la costa atlántica de África tropical (Anderson & Anderson, 2016). Estas especies poseen frutos adaptados a la dispersión hidrócora, como la presencia de aerénquima en las paredes del fruto, lo cual le permite una mayor flotabilidad (Anderson, 2001). Así, recientemente estas dos especies pudieron haber llegado a África por medio del océano Atlántico (Davis & Anderson, 2010).
Disyunciones neotropicales. Entre las especies presentes en Colombia, hay algunas que muestran disyunciones intra-americanas, como por ejemplo Byrsonima bucidifolia (presente en la isla caribeña colombianadeProvidencia,asícomoenlapenínsulade Yucatán, México), Ectopopterys soejartoi (conocida en Colombia únicamente en el norte de Antioquia, así como en algunas áreas de Ecuador y Perú), Hiraea villosa (conocida de algunas áreas colombianas del valle del río Magdalena, así como en algunas áreas del Perú), Stigmaphyllon bogotense (conocida únicamente de los Andes septentrionales y de Costa Rica), Stigmaphyllon sarmentosum (conocida únicamente del departamento colombiano de Cundinamarca en el centro del país, así como en algunas áreas de Ecuador y nordeste del Perú) y Tetrapterys magnifolia (conocida de algunas áreas de los departamentos de Antioquia y Cesar, en el norte de Colombia, así como en algunas áreas de Ecuador y Perú) (Anderson & Anderson, 2016). Estas disyunciones se deben analizar con cautela, ya que son evidentes los vacíos de inventarios florísticos en muchas áreas de Colombia. Así, esta distribución aparentemente disyunta quizás se deba a problemas y vacíos de muestreos e inventarios en varias regiones colombianas, como se mostrará más adelante.
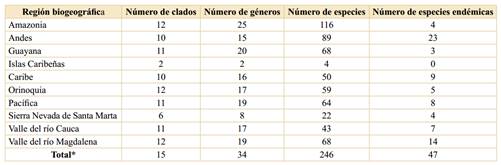
Tabla 3 Riqueza de las Malpighiaceae en las regiones biogeográficas de Colombia. *: La suma de las columnas no es aritmética, ya que hay varios clados, géneros y especies que se distribuyen en más de una región biogeográfica.
Diversidad filogenética de las Malpighiaceae en Colombia. En Colombia están representados 15 de los 19 clados reconocidos para la familia (véase Anderson et al., 2020), con 34 de los 77 géneros descritos (tablas 3 y 4). Los clados más diversificados en Colombia son el clado Stigmaphyllon con cinco géneros, 48 especies, diez de las cuales son endémicas de Colombia (5/48/10), el clado Byrsonima (3/39/5), el clado Hiraea (3/31/9), el clado Heteropterys (1/28/3), el clado Tetrapterys s. str. (2/27/6) y el clado Bunchosia (1/23/5) (Tabla 4). La diversidad filogenética en los gradientes altitudinales es similar al patrón de la riqueza específica por intervalo altitudinal, ya que las regiones naturales referidas a las tierras bajas (Amazonia, Orinoquia, valle del río Magdalena, Guayana, Pacífica, valle del río Cauca, Caribe), son las que presentan la mayor diversidad en clados. Si bien la Amazonia es la región más rica en especies, presenta el mismo número de clados (12) que la Orinoquia (una región que tiene la mitad de la riqueza específica de la Amazonia) y que el valle del río Magdalena; le siguen en cantidad de clados, la Guayana, la Pacífica y el valle del río Cauca, con once clados cada una (Tabla 3). A pesar de ser los Andes la segunda región natural más rica en especies (89), es la que ocupa el séptimo lugar en cuanto al número de clados (diez de los quince clados registrados en Colombia, faltando tanto clados basales como más recientes) (Tabla 3). Estas cifras son diferentes a las presentadas por Anderson & Anderson (2016), toda vez que aquí se amplía el número de géneros para Colombia, de 32 a 34, y el número de especies y por ende, de clados para varias unidades biogeográficas colombianas, como por ejemplo, pasar de diez especies para la Sierra Nevada de Santa Marta a 22, pasar de 57 especies para la Guayana a 68, de 42 especies para el valle del río Cauca a 43 y de 67 especies para el valle del río Magdalena a 68 especies, además, se actualizaron algunos datos para la distribución altitudinal de varias especies. No obstante, con nuevas exploraciones en varias áreas del territorio nacional, se esperaría que la Guayana colombiana presente el mayor número de especies y de clados, seguida, probablemente, por la Amazonia, ya que en ambas regiones sudamericanas se encuentran la mayor riqueza de especies y la mayor cantidad de linajes.
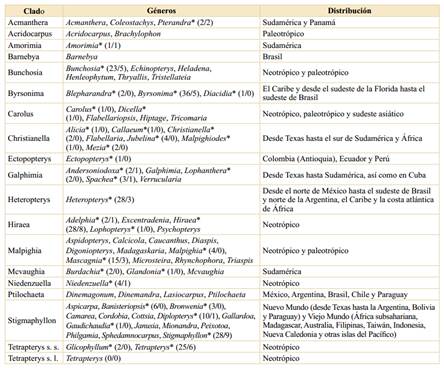
Tabla 4 Diversidad filogenética de las Malpighiaceae y su representatividad en Colombia. La configuración de los clados y su distribución geográfica están basadas en Anderson et al. (2020). *: Género presente en Colombia. Los números entre paréntesis corresponden al número de especies por género presentes en Colombia (totales/endémicas).
CONCLUSIONES
Colombia es el segundo país más rico en Malpighiaceae del mundo (34 géneros, 246 especies y un endemismo del 19,1%), después de Brasil [44 géneros, 584 especies y un 61% de endemismo (Mamede et al. 2010; Almeida et al., 2021)]. No obstante, son evidentes los pocos muestreos botánicos en muchas áreas colombianas (Giraldo- Cañas, obs. pers. realizadas durante casi 30 años de exploración en Colombia), por lo tanto, persisten los vacíos en torno de la riqueza biológica del país, ya que falta por explorar cerca de un 40% del territorio colombiano (Giraldo-Cañas, obs. pers.). En este sentido, son prioritarias las expediciones a la Sierra Nevada de Santa Marta (en el Caribe), la Guayana (principalmente en los departamentos de Caquetá, Guainía y Vaupés), la Orinoquia (principalmente en los departamentos de Arauca y Vichada), la región Pacífica (incluidas varias de sus serranías), la región del Catatumbo (Norte de Santander), varias zonas de las vertientes andinas (principalmente los cañones intra-andinos) y a pesar de que la Amazonia colombiana es la más rica, es evidente la necesidad de explorar varias áreas amazónicas de los departamentos del Amazonas, Caquetá, Guaviare y Putumayo. Es necesario destacar que gran parte de estas áreas estaban “prohibidas” para la investigación científica, debido a la guerra interna colombiana, la cual duró cerca de 55 años, y que tras la reciente firma de un acuerdo de paz están “abiertas” a variadas iniciativas de exploración e investigación (Baptiste et al., 2017).
El patrón de riqueza de especies, endemismo y diversidad filogenética de las Malpighiaceae en el contexto altitudinal colombiano muestra una alta riqueza en las tierras bajas. Este patrón coincide con varias familias, especialmente diversificadas en las tierras bajas neotropicales (0-1500 m s.m.), como Annonaceae, Apocynaceae, Arecaceae, Bignoniaceae, Chrysobalanaceae, Convolvulaceae, Euphorbiaceae, Fabaceae, Lamiaceae, Malvaceae, Marantaceae, Marcgraviaceae, Moraceae, Myristicaceae, Myrtaceae, Passifloraceae, Piperaceae, Poaceae, Rubiaceae, Sapotaceae, entre otras (Rangel- Ch., 2006; 2015; Bernal et al., 2016). No obstante, este comportamiento es diferente al exhibido por otros grupos vegetales, los cuales muestran un patrón correspondiente al “efecto del dominio medio” (Acanthaceae, Araceae, Bromeliaceae, Ericaceae, Gesneriaceae, hepáticas, líquenes, Orchidaceae, Pteridophyta, Solanaceae), toda vez que su mayor riqueza de especies se alcanza en altitudes intermedias (1500-2500 m s.m.) (Rangel-Ch., 2006; 2015; Bernal et al., 2016) y contrario a otras familias, las cuales alcanzan su riqueza más destacada en alturas mayores (≥ 2500 m s.m.), como en el caso de varios grupos (tribus, géneros) de las familias Apiaceae, Asteraceae, Calceolariaceae, Cunoniaceae, Hypericaceae, Juncaceae, Orchidaceae, Plantaginaceae, Poaceae, Ranunculaceae, Rosaceae, entre otras (Rangel-Ch., 2006; 2015; Bernal et al., 2016).
La consideración de regiones con elevada riqueza de especies, endemismo destacado y una apreciable diversidad filogenética, junto con la combinación de la representatividad de otros grupos biológicos, tanto vegetales como animales, podrían constituir insumos para definir y delimitar nuevas áreas prioritarias para la conservación en Colombia, y varias de estas regiones no cuentan con la suficiente representatividad de áreas biogeográficas con categorías de conservación en el “Sistema Nacional Ambiental”. Por otra parte, se recomienda evaluar, en un futuro cercano, el recambio, la abundancia, el desplazamiento y la extinción de especies en intervalos altitudinales, con miras a proporcionar nuevas evidencias en torno de los efectos del calentamiento climático global.

