











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
Las comunidades muscinales de las áreas australes de la Patagonia argentina y chilena, Tierra del Fuego e islas subantárticas, tienen una composición florística similar a las que crecen en el sur de Australia, Nueva Zelanda y Tasmania (Matteri, 1986), y parecerían ser “relictos” actuales del ancestral Gondwana (Wardle et al., 2001; Blöcher & Frahm, 2002; Matteri, 2003; Blöcher et al., 2006; Klazenga, 2012a, 2012b). Numerosas especies de musgos presenta este patrón de distribución fitogeográfica (Potrony Hechavarría et al., 2011).
Acrocladium Mitt. y Lepyrodon Hampe, son géneros de musgos pleurocárpicos, que habitan las regiones templado-frías del sur de Australia, Nueva Zelanda y América del Sur (Matteri & Schiavone, 1988, 2002; Klazenga, 2012a, 2012b; Larraín, 2016; Fife, 2018). La especie L. tomentosus (Hook.) Mitt., es la única que crece en Sudamérica y, además, en las áreas alto-andinas del Norte de México y el Caribe (Bowers & Sharp, 1975; Churchill & Linares, 1995; Fife, 2018).
El género Acrocladium presenta una controvertida posición taxonómica a nivel de familia. Inicialmente fue ubicado en “Stereodonteae” (Mitten, 1869), luego en Lembophyllaceae (Fleischer, 1923; Brotherus, 1925; Crum, 1991), Amblystegiaceae (Walther, 1983; Ochyra & Matteri, 2001), Plagiotheciaceae (Pedersen & Hedenäs, 2002) hasta ser incluido en una nueva familia, Acrocladiaceae por Tangney et al. (2010). Recientemente Goffinet & Buck (2020), lo han incluido nuevamente en la familia Lembophyllaceae, sin haber aún, acuerdo entre los autores. Lepyrodon en cambio, pertenece a la familia Lepyrodontaceae Broth. (Klazenga, 2012b; Goffinet & Buck, 2020).
Con base en estudios moleculares, Tangney et al. (2010), establecieron para Acrocladium una relación de grupo hermano con Lepyrodon. Sin embargo, se observó que los datos moleculares, no coinciden con las diferencias morfológicas observadas entre las especies.
Según las descripciones de varios autores, estos taxa muestran significativas diferencias. Acrocladium presenta gametofitos con características ramas cilíndricas terminadas en punta, que lo hacen fácilmente distinguible en el campo, son autoicos, amarillo-verdosos, con tallos poco ramificados, hojas oblongo- ovadas de ápices obtusos y margen entero, con células porosas y alares bien diferenciadas, en general sin nervio, y si está presente es muy corto, simple, rara vez doble; las cápsulas de los esporofitos son ovoideo-cilíndricas, arqueadas, inclinadas (Karczmarz, 1966; Ochyra & Matteri, 2001; Vanderpoorten et al., 2002a, 2002b; Hedenäs & Vanderpoorten, 2006; Klazenga, 2012a). El género Lepyrodon presenta plantas dioicas, amarillo a marrón- verdosas, con tallos muy ramificados y tomentosos, hojas oblongo-lanceoladas a triangulares, de ápice cuspidado y margen aserrado en la mitad superior de la lámina, con células vermiformes, sin células porosas, ni células alares, ni aurículas basales, nervio corto, simple o doble, a menudo bifurcado, cápsulas oblongo-cilíndricas y erectas (Allen, 1999; Klazenga, 2012b; Fife, 2018).
En la región fueguina de Argentina, Acrocladium, está representado por una única especie: A. auriculatum (Mont.) Mitt., y el género Lepyrodon por 2 especies: L. lagurus (Hook.) Mitt. y L. tomentosus (Matteri & Schiavone, 2002). Sin embargo, como especies dominantes del sotobosque de Nothofagus Blume (Lencinas et al., 2008), A. auriculatum y L. lagurus crecen a menudo asociadas entre sí, cubriendo a modo de tapices ásperos, con tallos entretejidos y plagiótropos, los distintos sustratos: troncos, ramas caídas, suelo y también como epífitas (Matteri & Schiavone, 1988, 2002). Para ambas especies, se han descripto las morfologías de sus gametofitos y esporofitos, pero no se han considerado, ni estudiado aún en detalle, las esporas, y solo se brindan datos de sus diámetros.
Está comprobado que en Bryophyta, las esporas de determinados taxa han aportado caracteres de interés en estudios sistemáticos. En 1979, Buck separó las familias Bruchiaceae y Dicranaceae (Bryophyta) a partir de la ornamentación de las esporas. Esta separación fue confirmada años más tarde, para especies de Brasil, por Luizi-Ponzo (1995) y Luizi-Ponzo & Barth (1998). Estébanez et al. (2006) encontraron para cuatro especies de Ptychomitrium Fürnr. de Japón, diferencias a nivel de la ultraestructura de las esporas. Khoshravesh & Kazempour (2007) destacaron el valor diagnóstico de la ornamentación de las esporas, tanto a nivel genérico, como específico en taxa del norte de Tehran (Irán). Piñeiro (2017) encontró diferencias en el tamaño y escultura de las esporas de Entosthodon laxus (Hook. F. & Wilson) Mitt. y Funaria hygrometrica Hedw., de los bosques fueguinos de Argentina.
En relación con las esporas de briofitas, si bien existen glosarios de términos palinológicos (Malcolm & Malcolm, 2006; Punt et al., 2006; Halbritter et al., 2018), con el avance de la microscopía electrónica, Reyre (1968, 1975) planteó la dificultad con el uso de una adecuada terminología para describir determinados tipos de esculturas “ultrafinas”. Por ejemplo, una verruga con gránulos sobre su superficie se describe como “verruga finamente granulada”. El autor propone para estos casos tipos de “ultra-escultura”, y si bien, su trabajo se refiere a granos de polen de Gimnospermas, esta situación se da muy a menudo al describir determinadas esculturas en esporas de briofitas.
Si bien hay publicaciones sobre esporas de musgos, tales como Medina & Estébanez (2014), Savaroğlu (2015), Savaroğlu et al. (2017), Costa Silva-e-Costa & Luizi-Ponzo (2019), entre otros, todavía son escasas las obras de consulta general y específica para las esporas de estas plantas. Se destaca el atlas de Boros et al. (1993), como una de las fuentes de información más completas hasta el momento, que brinda descripciones de las esporas de los musgos europeos, con microscopio óptico y algunas fotos con microscopio electrónico de barrido. Pero en particular, para las especies fueguinas Acrocladium auriculatum y Lepyrodon lagurus, no hay descripciones de las esporas, por lo que planteamos en este trabajo, hacer un análisis comparativo para establecer el valor diagnóstico de los caracteres que las esporas podrían aportar a la sistemática de estos taxa.
MATERIALES Y MÉTODOS
Para el estudio se seleccionaron y emplearon ejemplares de herbario de Acrocladium auriculatum y Lepyrodon lagurus del Museo Argentino de Ciencias Naturales “Bernardino Rivadavia” (BA), Ciudad Autónoma de Buenos Aires, Argentina.
Las esporas se estudiaron con Microscopio Óptico (MO) y Microscopio Electrónico de Barrido (MEB). Las esporas fueron acetolizadas (Erdtman, 1960) durante 2 minutos. Las observaciones, sobre 25-30 esporas, se hicieron sobre preparados montados en gelatina-glicerina, utilizando un microscopio óptico, Olympus CH-2. Para el análisis con MEB, las esporas fueron hidratadas en agua destilada (24 hs), fijadas en formaldehído, alcohol, ácido acético- FAA (20 días), deshidratadas en series graduadas de alcohol etílico (20-70%) y secadas por punto crítico. Las muestras fueron metalizadas con oro-paladio y montadas sobre cinta bifaz. Las fotografías se tomaron con un microscopio JEOL, JSMT-100.
Para la descripción de las esporas se utilizó la terminología empleada por Kremp (1965), Reyre (1968, 1975), Clarke (1979) y Punt et al. (2006), y la utilizada en los trabajos de Olesen & Mogensen (1978), Mogensen (1981, 1983) y Brown & Lemmon (1990), para las esporas de musgos en particular.
Material examinado
Acrocladium auriculatum (Mont.) Mitt.
ARGENTINA. Tierra del Fuego. Depto. Río Grande, Hito XIX, 6-II-1942, Castellanos s/n (BA 32041). Depto. Tolhuin, Tolhuin, 23-IV-1994, Roig 15690 (BA 46561). Depto. Ushuaia, Lapataia, 11- I-1974, Matteri 1724 (BA 25577); Ruta J, 16-I-1974, Matteri 1825 (BA 25649); Rancho Hambre, 23-IV-2006; Fuertes & Rodriguez s/n, (BA 45580); Pto. Ushuaia, Río Olivia, 27-IX-1979, Matteri 2142 (BA 30838).
Lepyrodon lagurus (Hook.) Mitt.
ARGENTINA. Tierra del Fuego. Depto. Ushuaia, Pto. Harberton; 24-XII-1932, Castellanos s/n (BA 1326); Lapataia, 11-I-1974, Matteri 1726 (BA 25549); Lapataia, 11-I-1974, Matteri 1740 (BA25593); Ruta de la Costa a 12 km al O de Ea. Moat, 16-II-1984, Matteri & Schiavone 2585 (BA 2585, 2536); Bahía Buen Suceso, 19-I-1984, Matteri & Schiavone 3516 (BA 3516).
RESULTADOS
Para cada especie estudiada, se describieron con MO y MEB los siguientes caracteres de las esporas: forma, presencia o no de lesura, tamaño, y ornamentación de la superficie (Tabla 1, Fig. 1).
En Acrocladium auriculatum, las esporas son esferoidales, aletes, de 10 a 15 µm de diámetro, y la superficie de la perina es ornamentada. Se distingue una escultura formada por un nivel inferior de procesos pequeños menores a 0,25 µm, con forma de gránulos y verrugas, irregularmente distribuidos, y un nivel superior de elementos baculado-gemados, separados, de 0,7-1 µm de altura, sobre los que se observa una ultra-escultura de verrugas, gemas y gránulos (Fig. 1: A, C, E, G). Es común observar esporas abolladas sobre una de sus caras.
En Lepyrodon lagurus, las esporas son esferoidales, aletes, de 12 a 20 µm de diámetro, con superficie de la perina ornamentada. La escultura presenta procesos baculado- crestados, de 0,3-0,5 µm de altura, densamente distribuidos, sobre los que se observa una ultra-escultura de gránulos y verrugas que se agrupan en “conglomerados” terminales (Fig.1: B, D, F, H). Es común observar esporas abolladas sobre una de sus caras.
DISCUSIÓN Y CONCLUSIONES
El estudio comparativo entre las esporas de las especies fueguinas, Acrocladium auriculatum y Lepyrodon lagurus, nos ha permitido establecer similitud en la forma y en la ausencia de lesura, ya que en ambos casos son esferoidales y aletes.

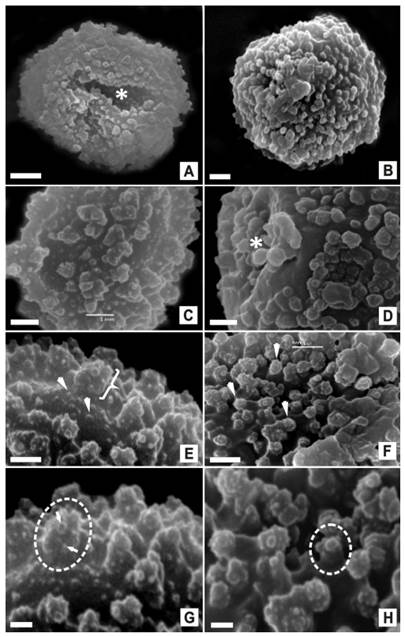
Fig. 1 Esporas de Acrocladium auriculatum y Lepyrodon lagurus con MEB. A. auriculatum (A, C, E, G). A, vista proximal de una espora esferoidal, alete, de superficie ornamentada, en la que se observa cómo la espora se abolla sobre esta cara, posible leptoma (Asterisco). C, vista general de la superficie con procesos ornamentales en dos niveles. E, superficie de la espora en la que se observan procesos pequeños irregularmente distribuidos, en un nivel inferior (Flechas), y procesos de mayor tamaño, separados, en un nivel superior (Llave). G, detalle de un proceso baculado-gemado (Círculo), sobre el que se observa la ultra-escultura de pequeños procesos con forma de verrugas, gemas y gránulos (Flechas). Sobre el nivel inferior gránulos y verrugas distribuidos. L. lagurus (B, D, F, H). B, vista distal de una espora esferoidal, alete y superficie ornamentada. D, vista ecuatorial en la que se observa cómo la espora se abolla sobre una de las caras, posible leptoma (Asterisco). F, superficie de la espora en la que se observan procesos baculado-crestados, densamente distribuidos, sobre los que se visualizan pequeños procesos terminales (Cabezas de flecha). H, detalle de uno de los procesos baculado-crestado (Círculo) sobre el que se observa la ultra-escultura de procesos redondeados y agrupados en “conglomerados terminales”. Escalas: 2 µm (A, B); 1 µm (C, D, F, G); 0,5 µm (E, H).
En relación con esta última característica, si bien no se visualizan lesuras, la frecuencia de esporas abolladas sobre una de sus caras, hace sospechar la presencia de una posible zona adelgazada a nivel de las paredes, lo que varios autores han observado y definido en esporas de otras especies de musgos (Funaria muhlenbergii Turn., Funaria hygrometrica Hedw., Hypnum vaucheri Lesq., Tortula atrovirens (Sm.) Lendb.), como un leptoma (Boros et al., 1993; Ghi-sen et al., 1997; Lashin, 2011; Piñeiro, 2017).
En Acrocladium auriculatum, las esporas son en general más pequeñas, no superan los 15 µm, mientras que en Lepyrodon lagurus pueden llegar a medir hasta 20 µm de diámetro. Se diferencian por el tipo de ornamentación. A. auriculatum, presenta una escultura doble, formada por procesos esculturales ubicados en dos niveles (Fig.1: E, G). En L. lagurus la escultura está formada por procesos densamente distribuidos sobre la superficie de la espora (Fig.1: B, F). En ambas especies, sobre los procesos esculturales, es posible observar una ultra-escultura, representada por verrugas, gemas y gránulos en las esporas de A. auriculatum (Fig.1: G), y gránulos y verrugas agrupados en conglomerados terminales, en las esporas de L. lagurus (Fig.1: F, H).
El diámetro de las esporas de Lepyrodon, estudiadas por Allen (1999) para la región fueguina, parece ser un carácter variable, ya que en L. lagurus miden entre 10-14 µm y en L. tomentosus entre 15-25 µm. En nuestras muestras estudiadas, se advierte que las esporas de L. lagurus presentan un mayor rango de tamaño (12-20 µm), similar al que presentan las esporas de Nueva Zelanda, con diámetros entre 12-17 µm (Fife, 2018).
A partir de las características observadas, se destaca que los caracteres de las esporas contribuirían a reforzar las diferencias morfológicas, gametofíticas y esporofíticas, entre las plantas de las dos especies estudiadas. Este resultado coincide con la nueva clasificación de Goffinet & Buck (2020), quienes ubican a estos géneros en familias diferentes. Podemos concluir que las esporas contribuyen como una potencial y complementaria fuente de información para la sistemática y biología de estos taxa.

