











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkLa creciente urbanización es una de las principales preocupaciones para la conservación de la biodi-versidad (Dearborn y Kark 2010), ya que modifica el paisaje y afecta la distribución y abundancia de especies. Las perturbaciones humanas (peatones, animales domésticos, entre otras) pueden tener efectos adversos comparables con los cambios en la estructura y composición de la vegetación (Papouchis et al. 2001, Parris y Schneider 2008, Schlesinger et al. 2008, Rebolo-Ifrán et al. 2015). Por ello, se requiere de una mayor comprensión del impacto que tienen las crecientes perturbaciones humanas sobre la biodiversidad (Blumstein et al. 2005, Tatte et al. 2018).
Al escapar de predadores en ambientes urbanos, las aves que se alimentan en el suelo pueden elegir entre dos estrategias: correr o volar (Rodriguez-Prie-to et al. 2008). Estos comportamientos de huida presentan distintos costos y beneficios para el individuo (Ydenberg y Dill 1986, Blumstein 2019), teniendo el primero un menor gasto energético pero mayor riesgo de depredación y viceversa. Los costos energéticos asociados con correr son más bajos (Butler 1991), ya que las aves pueden luego reanudar rápidamente el forrajeo y permiten al individuo tener un período más largo para evaluar el riesgo real que supone un depredador (Domenici y Blake 1993, Cooper et al.
2007), lo que podría optimizar las decisiones de escape posteriores (Rodriguez-Prieto et al. 2008). Estudios previos señalaron que el correr como estrategia de escape ocurre cuando se percibe un bajo riesgo de depredación (Tatte et al. 2018, 2020). Sin embargo, el individuo está expuesto a un mayor riesgo de depredación, ya que la velocidad de escape es menor y la distancia al depredador es más corta al final del escape (Nuevo 2004). Por otro lado, al volar las presas reducen en gran medida este riesgo, pero a expensas de altos costos energéticos (Butler 1991) y pérdida de oportunidades de forrajeo (Brown y Kotler 2004).
Existen múltiples factores que afectan el comportamiento de escape, incluyendo factores relacionados con el tamaño del ave, el comportamiento del depredador y con el entorno social y físico (Lima 1993, Stankowich y Blumstein 2005, Blumstein 2006, Mikula 2014, Morelli et al. 2019, Santos et al. 2022). A pesar de que los entornos urbanos pueden parecer en un principio homogéneos, son en realidad muy heterogéneos en sus condiciones ambientales, abundancia y diversidad de depredadores y actividades humanas (Mikula 2014, Moller y Tryjanowski 2017, Morelli at al. 2018); como demuestran estudios previos que encontraron diferencias en la distancia de vuelo de las aves ante la aproximación de humanos (FID) en parques y cementerios urbanos (Morelli et al. 2018).
Las condiciones locales afectan directamente el comportamiento de las aves (Botsch et al. 2017, Fer-nández-Juricic et al. 2002). Por ejemplo, árboles, arbustos y lápidas pueden ser utilizados como posaderos y refugios (Cooper 2003, Tryjanowski et al. 2017), lo que para algunas especies significa una disminución del riesgo de depredación (Morelli et al. 2022). A su vez, las aves se han adaptado a los entornos urbanos disminuyendo su temor a los humanos (Moller 2008, Moller et al. 2013, Piratelli et al. 2015, Cavalli et al. 2016, Samia et al. 2017, Biondi et al. 2020). En ambientes urbanos, se observa una disminución en las respuestas anti depredador que pueden estar relacionadas a una habituación a los constantes estímulos no letales causados por el hombre (Mccleery 2009) o a una variabilidad intra-específica del comportamiento de huida, donde los individuos más “audaces” son capaces de tolerar estos ambientes perturbados (Carrete y Tella 2010).
La estacionalidad trae aparejadas variaciones en la temperatura y en la duración de los ciclos de luz-oscuridad, entre otros cambios. Estos factores extrínsecos provocan cambios en los recursos (Leveau 2018), tales como un aumento o disminución en la disponibilidad de frutos o insectos, así como cambios intrínsecos en los individuos, en particular en la conducta debido a la nidificación (Deviche y Davies 2014). A su vez, durante la época reproductiva podría haber un mayor riesgo de depredación de las crías y de los individuos adultos o reproductores (Ciuti et al. 2008).
El forrajeo en grupo trae beneficios en términos de seguridad, generalmente a través de menores posibilidades de ser atrapado debido al efecto de dilución del riesgo de depredación (Roberts 1996, Fer-nández-Juricic et al. 2002, Stankowich y Blumstein 2005, Moller 2015): un individuo en un grupo grande reduce sus posibilidades de ser detectado cuando ocurre un ataque (Fernández-Juricic et al. 2002). La sociabilidad trae aparejada una mayor vigilancia debido al efecto de muchos ojos o “many eyes” (Stankowich y Blumstein 2005) y puede ser un factor clave en la evolución del comportamiento anti depredador (Mo-relli et al. 2019). Estudios previos revelaron que, ante una amenaza, una bandada emprende el vuelo más rápidamente que los individuos solitarios o los individuos en pequeños grupos (Tatte et al. 2018, Morelli et al. 1019, Garcia-Arroyo y MacGregor-Fors 2020). Esto surge porque la amenaza se detecta antes y, por lo tanto, se emprenden acciones evasivas con mayor rapidez para evitar altos costos de vigilancia (Blumstein 2010, Samia y Blumstein 2015). A su vez, las respuestas de miedo pueden transmitirse socialmente (Griffin 2004), de manera que la huida también se podría dar como consecuencia de efectos en cascada o un fenómeno contagioso (Sirot 2006, Morelli et al. 2019).
El comportamiento de escape varía según la especie (Blumstein et al. 2003, Spindler Díaz 2021), lo cual puede deberse al tamaño del cuerpo o a factores intrínsecos propios de la misma. Se ha observado una tendencia a escapar corriendo en aves de menor tamaño (Kullberg y Lafrenz 2007). A pesar de que existen numerosas explicaciones biológicas de por qué varía el comportamiento de huida según el tamaño del cuerpo (e.g. Tatte et al. 2018), esto podría deberse a que los animales más pequeños deben correr mayores riesgos ya que sus requerimientos metabólicos son relativamente más altos (Grant y Noakes 1987).
Los factores que afectan la selección de una u otra estrategia de escape han sido poco estudiados (Rodri-guez-Prieto et al. 2008, Prestes et al. 2018). Por lo tanto, el objetivo de este estudio es analizar la variación de diferentes estrategias de escape en aves en relación a los factores intrínsecos (especie, tamaño promedio del ave) o extrínsecos (cobertura vegetal e infraestructura, disturbios humanos, estación del año, gregarismo, hábitat) recién mencionados.
La cobertura vegetal e infraestructura pueden ser utilizados como refugios (Cooper 2003, Tryjanowski et al. 2017), lo que significaría una disminución en el riesgo de depredación. Por lo tanto, a mayor cobertura e infraestructura, se espera encontrar una tendencia a correr. Por otro lado, dado que el riesgo de depredación y mortalidad por el ser humano es bajo (Mccleery 2009), las aves en los espacios verdes urbanos tenderían a responder disminuyendo en su respuesta de escape (Díaz et al. 2013, Moller y Liang 2013), asumiendo mayores riesgos que aquellas en ambientes no urbanos y utilizando la estrategia de correr. A su vez, se espera que el comportamiento de escape se vea afectado según la especie (Blumstein et al. 2003, Spindler Díaz 2021) y que el escape de vuelo sea mayor durante la época reproductiva debido al mayor riesgo de depredación de las crías y de los propios individuos (Ciuti et al. 2008). Por otra parte, se espera mayor escape en vuelo a mayor tamaño bandada, debido a un mayor efecto de contagio (Sirot 2006, Morelli et al. 2019). El hábitat (parque/cementerio) también afectaría el tipo de escape, siendo la estrategia de correr más frecuente en parques que en cementerios debido al mayor acostumbra-miento de las aves hacia los humanos en los parques. Por último, se espera que aquellas especies de menor tamaño escapen corriendo (Kullberg et al. 2007) debido a que sus requerimientos metabólicos son mayores (Grant y Noakes 1987) y esta estrategia de escape les permitiría reanudar rápidamente el forrajeo.
métodos
Área de estudioEl estudio se llevó a cabo en la Ciudad Autónoma de Buenos Aires (34°35'59"S 58°22'55"O, 25 m.s.n.m., 3 075 646 hab.), Argentina. La temperatura media anual es de 17,9 °C, siendo la época reproductiva (primavera-verano) la estación más calurosa y la no reproductiva (otoño-invierno) la más fría (Servicio Meteorológico Nacional [SMA] s.f.). La ciudad de Buenos Aires se encuentra en la Pampa Ondulada, la cual estuvo originariamente compuesta por una estepa gramínea y parches de bosques xerofíticos (Oyarzabal et al. 2018), aunque actualmente está dominada por cultivos y plantaciones de árboles exóticos. La precipitación media anual es de 1236,3 mm, siendo la época reproductiva la más lluviosa y la no reproductiva la más seca (SMA s.f.). Se estudiaron tres parques y tres cementerios. En total, se exploraron 12 puntos para parques y 16 puntos para cementerios.
Diseño general
Los datos sobre estrategia de escape y las variables ambientales se midieron y recopilaron en los siguientes cementerios y parques de la Ciudad Autónoma de Buenos Aires: Cementerio de la Chacarita, Cementerio de Flores, Cementerio de la Recoleta, Parque Avellaneda, Parque Las Heras y Facultad de Agronomía y Veterinaria.
Los sitios fueron seleccionados según los siguientes criterios: para cada uno de los tres cementerios se eligió un parque cercano de similar tamaño con el cual compararlo, siendo la distancia entre ellos de al menos 1 km para garantizar independencia en los datos. Un solo observador recolectó los datos en distintos puntos de los cementerios y parques. Estos puntos se encontraban separados por al menos 200 m y cubriendo la mayor parte de las áreas dentro de los cementerios y parques, asegurando el muestreo de diferentes tipos de ambientes.
Los mismos sitios y puntos se visitaron dos veces en cada temporada no reproductiva y reproductiva de 2019, con un intervalo de 3 a 4 semanas. Considerando que en los espacios verdes de la ciudad los disturbios humanos varían entre días de semana (lunes a viernes) y fin de semana (sábados y domingos), en cada temporada los sitios fueron visitados una vez en cada momento. De esta manera, se buscó minimizar un potencial sesgo vinculado a los patrones de disturbios humanos.
Recolección de datosLa recolección de datos se realizó en días sin lluvia ni viento fuerte y durante las primeras cuatro horas de la mañana. Los datos sobre estrategia de escape se recopilaron siguiendo un procedimiento estándar para todos los individuos: cuando se localizaba un ave focal individual situada en el suelo, el observador (que vestía ropa de colores apagados para pasar más desapercibido) se movía a una velocidad constante caminando hacia el ave (Díaz et al. 2013) y registraba el tipo de huida (vuelo/caminata). Estudios previos han demostrado que la distancia desde la cual el observador inicia el acercamiento está fuertemente correlacionada con el comportamiento de escape (Moller y Liang 2013), lo que potencialmente causa un problema de colinealidad en el análisis estadístico. Sin embargo, este problema se eliminó al comenzar el acercamiento a la mayoría de los individuos desde una distancia mayor a 30 m del observador, manteniendo las distancias iniciales relativamente constantes (Díaz et al. 2013).
En este trabajo se tuvo también en cuenta si el ave se encontraba en bandada (medido como el número de aves circundantes en un radio de 2 m) y la presencia de predadores/personas en la cercanía (sí/ no). Las bandadas compuestas por múltiples especies también fueron incluidas, ya que se ha encontrado que estas también modifican el comportamiento de escape (Sridhar et al. 2009). Además, se tuvo en cuenta el tamaño promedio de la especie en cm, para ello se utilizaron datos de Canevari et al. (2017).
Se buscó obtener datos de la mayor cantidad de individuos y especies posibles. La recolección de datos sólo se realizó para individuos que se encontraban en un "comportamiento de confort”, como por ejemplo forrajeando, acicalándose o caminando. Por lo tanto, se evitó tomar datos de aves que ya estaban muy alertas (e.g. realizando vocalizaciones de alerta u observación hacia los alrededores) o que se encontraban en sus nidos.
Las características ambientales y datos de actividad humana fueron recolectados dentro de un radio de 100 m alrededor de cada punto (Morelli et al. 2018) y previo a la observación del comportamiento de los individuos. Cada punto fue caracterizado por las siguientes variables ambientales (Tabla 1): a) cobertura vegetal: fue estimada considerando dos niveles (cobertura de los árboles y cobertura a nivel del suelo - cobertura de arbustos, vegetación herbácea, césped y suelo desnudo-), los cuales se consideraron en dos escalas independientes a través de estimaciones visuales del porcentaje de cobertura (del 0 al 100%) dentro de un radio de 100 m. Se consideró arbustos a vegetación leñosa de varios tallos, menor a 2 m de altura.; b) infraestructura humana: incluye el porcentaje de cobertura edificada, lápidas y número de postes de luz; c) disturbios humanos: densidad de personas (estimado como el número de personas registradas en un intervalo de 10min), velocidad de los peatones (número de pasos registrados en 10 segs), proporción hombres/mujeres (en un intervalo de 10min), otros disturbios tales como el número de autos o motos, y de mamíferos; y d) el tamaño de cada sitio (superficie de cada parque / cementerio) en hectáreas, utilizando Google Earth. Por otro lado, se analizó la relación entre la estrategia de escape y el tipo de área verde (parque o cementerio).
Tabla 1: Variables utilizadas para evaluar el comportamiento de escape en la Ciudad de Buenos Aires, Argentina. Abreviaciones: DE - desvío estándar.
Análisis estadísticoCon el fin de evitar la multicolinealidad entre variables ambientales, se realizó una matriz de correlación (Material Suplementario, Anexo A) utilizando el paquete ‘corrplot’ para R (Wei et al. 2017, R Develo-pment Core Team 2017). Sólo aquellas variables con un valor de coeficiente de Pearson menor a 0,6 fueron incluidas en los modelos (Graham 2003). A su vez, se utilizó un test de Student para comparar las variables de disturbios humanos entre parques y cementerios (Gardener 2017). Con el fin de analizar la correlación entre estas variables recién mencionadas, el test de Student fue convertido al índice de Pearson utilizando la ecuación provista por Rosenberg (2010) (Material Suplementario, Anexo B). Para explorar el uso de una u otra estrategia de escape (0 = volar, 1 = correr) en relación a las distintas variables explicativas (Tabla 1), se emplearon modelos lineales generalizados mixtos (GLMMs). Se utilizó el paquete ‘lme4’ (Bates et al. 2014) para llevar a cabo los modelos y la función ‘glmer’. Se realizó una "backward selection” y el criterio de información Akaike (AIC) fue utilizado para determinar el mejor modelo (Burnham y Anderson 2002): luego de evaluar todos los modelos, se seleccionó aquel de menor AIC y que presentaba una diferencia mayor a dos respecto al siguiente modelo. Se analizaron posibles interacciones entre las variables explicativas Época y Bandada, así como entre Época y Hábitat.
Los puntos, sitios y especies fueron incorporados a los modelos como factores aleatorios. Esto es debido a que los puntos se encontraron anidados dentro de los parques y cementerios y, además estos fueron estudiados a lo largo del tiempo. A su vez, se introdujo a la variable especie como factor aleatorio teniendo en cuenta la dependencia filogenética entre especies (Garamszegi y Moller 2011). El modelo se ajustó asumiendo una distribución binomial de errores. Los gráficos finales se elaboraron utilizando el paquete ‘ggeffects’ (Lüdecke 2018). Se utilizó R (R Develop-ment Core Team 2017) para llevar a cabo los distintos análisis y gráficos.
resultados
Se recolectaron datos del comportamiento de huida para 24 especies de aves urbanas, de las cuales 16 especies tuvieron más de 2 muestras y 4 especies más de 30 muestras. Las especies más muestreadas fueron Furnarius rufus con 97 datos de comportamiento de huida, Turdus rufiventris con 91 datos, Zenaida au-riculata con 72 datos y Mimus saturninus con 39 datos.
La probabilidad de seleccionar una u otra estrategia varió entre especies, la época del año y el número de aves que conformaban un grupo (bandada) (Figura 1 y 2). Mimus saturninus, Furnarius rufus, Columba liviay Patagioenaspicazuro presentaron una tendencia a correr, mientras que Myiopsitta monachus presentó una tendencia a volar. A su vez, las aves eligieron preferentemente la estrategia de vuelo durante la época reproductiva y cuando se encontraban formando bandadas de mayor tamaño (Figura 2, Tabla 2).
Tabla 2: Resultados de los modelos lineales generales mixtos entre las variables explicativas y el comportamiento de escape de aves. Los sitios, los puntos y las especies fueron añadidos como factores aleatorios de los modelos. Solo se muestran las variables significativas correspondientes al modelo final. Abreviaciones: OR- odds ratios; IC - intervalos de confianza (mín., máx.); EE - error estándar; P - p-valor.
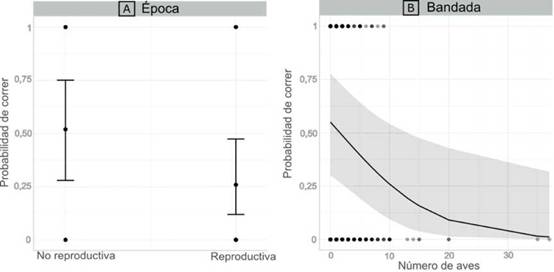
Figura 1: Tendencia de las especies (odds ratio) a realizar una u otra estrategia de escape (correr o volar). El odds ratio es una medida de la magnitud del efecto, y expresa cuántas veces es mayor la probabilidad del evento por unidad de cambio de la variable. El eje Y representa las distintas especies, el eje X los odds ratio. Se muestra el odds ratio (círculo del centro) y los intervalos de confianza (líneas horizontales). Por debajo de uno, las especies tendrán una tendencia a volar, mientras que por arriba de uno las especies tenderán a correr. Para que sea significativo, los intervalos de confianza no deben tocar a este valor (uno).
Figura 2: A. Probabilidad de un ave de correr como comportamiento de huida en función de la época del año. El eje Y representa la probabilidad de correr, el eje X la época (no reproductiva/reproductiva). Se muestran la media (círculo del centro) y el error estándar (líneas verticales). Una probabilidad de 0 indica que se utilizó el vuelo como estrategia de escape. B. Probabilidad de un ave de correr como comportamiento de huida en función del tamaño de la bandada. El eje Y representa la probabilidad de correr, el eje X el tamaño de la bandada (número de aves). Se muestran el ajuste del modelo (línea negra) y el intervalo de confianza del 95% (en gris). Una probabilidad de 0 indica que se utilizó el vuelo como estrategia de escape.
discusión
Los resultados obtenidos en este estudio indican que las aves corren como método de huida en mayor proporción durante la época no reproductiva. No obstante, ellas tienden a volar en mayor proporción a mayor tamaño de bandada. A su vez, ciertas especies presentaron una tendencia a correr como estrategia de escape.
Al igual que en estudios previos (Blumstein et al. 2003, Spindler Díaz 2021), hemos observado diferencias en los comportamientos anti depredadores de distintas especies. A pesar de que estas diferencias no estarían relacionadas con el tamaño corporal del animal, pueden ser explicadas por factores intrínsecos propios de cada especie de ave, tales como diferencias en las habilidades cognitivas y sensoriales (Tisdale y Fernández-Juricic 2009) o al hecho de que la mayoría de los datos fueron tomados de cuatro especies de similar tamaño, con lo cual sería necesario reunir un mayor número de datos para poner a prueba la hipótesis de la influencia del tamaño. A su vez, es importante tener en cuenta que el comportamiento depende de otros factores que no han sido analizados en este estudio, como por ejemplo la edad del individuo (Thornton y Lukas 2012).
Los recursos en las áreas verdes de las ciudades son relativamente estables y abundantes debido al control humano (Shochat et al. 2006, Fuller et al. 2008, Leveau 2018), lo cual significa que las aves que habitan estos espacios pueden adquirir recursos durante todo el año (Zhou y Chu 2012), alterando la es-tacionalidad de las comunidades de aves en comparación a las áreas rurales o naturales (Leveau 2021). Sin embargo, se observaron cambios en el comportamiento de escape entre las épocas reproductiva/no reproductiva, lo que indica que la estacionalidad de los comportamientos anti depredadores no se ve lo suficientemente amortiguada en áreas urbanas, a diferencia de lo observado por Uchida et al. (2016). Es importante mencionar que Zenaida auriculata es una migradora parcial: durante la época reproductiva se encuentra en grandes números en la ciudad de Buenos Aires, mientras que en la época no reproductiva quedan unos pocos individuos (L. Leveau obs pers.). Por lo tanto, no hubo datos recolectados durante la época no reproductiva para esta especie.
La estacionalidad del comportamiento de escape de las distintas especies de aves podría deberse a cambios ambientales propios de las estaciones, tales como variaciones en la disponibilidad de recursos, en la temperatura y en la duración de los ciclos de luz-oscuridad, que afectan tanto factores extrínsecos como intrínsecos de los individuos. De esta manera, la preferencia de un tipo de huida menos riesgosa como el escape en vuelo durante la época reproductiva podría estar relacionadas a un mayor riesgo de depredación de las crías y de los propios individuos (Ciuti et al. 2008). Estos resultados resaltan la capacidad de las aves para adaptar sus comportamientos de escape a las condiciones ambientales locales cambiantes. Por otro lado, el tipo de escape no se relacionó significativamente con la estructura del hábitat, indicando que el tipo de huida es independiente de las diferencias en la cobertura vegetal o infraestructura (Moller et al. 2013). Sin embargo, en nuestro estudio los datos se recolectaron en unidades de 100 m de radio, y sin duda observaciones tomadas a escalas menores podrían arrojar diferentes asociaciones entre los métodos de huida y las variables ambientales. Por ejemplo, la cercanía a perchas o arbustos podría influir en la estrategia de huida.
Como respuesta a la amenaza de depredación, las aves a menudo forman grupos mientras se alimentan (Sirot 2006). Existen dos grandes hipótesis con respecto a los comportamientos anti depredadores y al tamaño de la banda. Por un lado, formar parte de un grupo podría disminuir el riesgo de depredación sobre un ave (efecto de dilución), lo que podría derivar en una disminución en las respuestas anti depredador. Por el otro lado, un número más elevado de individuos podría significar que estos se hallan en un mayor estado de alerta (efecto "many eyes”) y por ello podrían tener respuestas de escape más rápidas (Tatte et al. 2018, Morelli et al. 2019). Nuestros resultados señalaron que las aves eligen preferentemente la estrategia de escape más costosa energéticamente y más rápida (volar) al encontrarse en grupos de mayor tamaño, lo cual respalda la hipótesis del efecto "many eyes” y podría indicar un fenómeno "contagioso”, donde el vuelo de un primer individuo ocasiona el vuelo coordinado del resto (Griffin 2004, Sirot 2006, Morelli et al. 2019). Estos resultados ponen en evidencia que la presencia de numerosos individuos en un grupo puede alterar significativamente el comportamiento de escape de aves y que por ello las especies gregarias pueden ser más susceptibles a la perturbación humana, tanto a nivel individual como poblacional (Weston et al. 2012).
Estos resultados sugieren que las aves tienen un mecanismo para ajustar su comportamiento y hacer frente a los cambios estacionales. Por otro lado, la selección de una u otra estrategia de escape no está vinculada a disturbios humanos, superficie de cada sitio ni a los distintos hábitats (parque / cementerio). Dado que el riesgo de depredación y mortalidad por el ser humano es bajo (Mccleery 2009), las aves en los espacios verdes urbanos tienden a responder mediante una disminución en su respuesta de escape (Díaz et al. 2013, Moller y Liang 2013, Prestes et al. 2018). El hecho de no haber encontrado una asociación con los constantes estímulos creados por los seres humanos y mamíferos parece respaldar la observación general de que los hábitats urbanos presentan un riesgo bajo de depredación (Ibáñez-Álamo y Soler 2010). Finalmente, una limitación de este estudio fue que las observaciones se realizaron en un único año. Por ello, futuras investigaciones debieran considerar realizar muestreos de mayor duración, así como enfocarse en obtener un número más elevado de individuos de diversas especies, particularmente de aquellas que presentan distintos tamaños corporales.
Recibido: 28 de junio de 2022
Aceptado: 4 de octubre de 2022

