











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkUrbanization is a major and increasing cause of biodiversity loss worldwide (McDonald et al. 2018). Urban areas are one of the fastest growing habitats in both spatial coverage and ecological impact (Grimm et al. 2008). Therefore, the analysis of the effect of urbanization on biodiversity is imperative to design sustainable cities.
Birds have been widely used in urban studies because they have high taxonomic diversity (Jetz et al. 2012), represent one of the best-known wildli-fe groups (Schulze et al. 2004), are relatively easily to survey (Ralph et al. 1995), and quickly respond to modifications in their habitats (Marzluff et al. 2001, Aronson et al. 2014). In general, studies have analyzed the effect of urbanization on bird commu-nities through the comparison of land uses with di-fferent levels of urbanization (Blair 1996, Leveau and Leveau 2004, Villegas and Garitano-Zavala 2010). Some studies have found a peak of bird diversity at moderately urbanized land uses (Blair 1996, Marzlu-ff 2005, Tratalos et al. 2007, Leveau 2019), probably because of an increase in habitat diversity and re-sources availability in suburban areas (Leveau 2019). Habitat diversity and resources promote high bird diversity, allowing the presence of bird niche specia-lists (Hurlbert 2004). On the other hand, other studies have found a negative relationship between urbanization and bird diversity (Ortega-Álvarez and MacGregor-Fors 2009, Villegas and Garitano-Zavala 2010, Batáry et al. 2018), probably because of a continuous decrease of habitat diversity and resources at higher levels of urbanization. In general, this negative asso-ciation between bird diversity and urbanization has been found in tropical and subtropical areas (Koh et al. 2006, Leveau et al. 2017, Filloy et al. 2019), where urban habitats are compared with forest, which have a higher habitat diversity and primary productivity.
Urbanization has significant effects on bird spe-cies composition (Leveau and Zuria 2017). Although urban bird communities are mainly composed of native species (Aronson et al. 2014), the most urba-nized areas within cities are usually dominated by a few cosmopolitan species, such as the Rock Dove (Columba livia) and the House Sparrow (Passer domesticus) (Leveau and Zuria 2017). These species are able to exploit urban environments given that they feed on food discarded by humans and nest in buildings. The replacement of native species by a few widespread species has been called biotic homogenization (Mc-Kinney and Lockwood 1999).
Among tropical and subtropical regions, Latin America has recently gained presence in the urban bird literature (Ortega-Álvarez and MacGregor-Fors 2011a, 2011b). This region shelters an important proportion of global avifauna (BirdLife International 2013), encompassing important hotspots for birds (Mittermeier et al. 2011), which are currently expo-sed to high urbanization rates. Also, most studies are carried out in big or capital cities ignoring small or medium-sized cities where high population growth rates are expected to take place (United Nations 2015, Kendal et al. 2020). More studies in these types of ci-ties are needed to find useful information to develop management and conservation actions (Allred et al. 2021).
In this context, the aim of the current study was to determine the effect of urbanization on birds’ diversity and composition in a medium-sized Argentine city located in a biogeographic transition zone between an open xerophytic forest and a riparian subtropical forest (Oyarzabal et al. 2018). We expected a negative relationship between urbanization and bird diversity. In addition, we expected a dominance of exotic spe-cies in the most urbanized areas.
methodsStudy Area
The study was performed in the city of Santo Tomé (31°40'0" S, 60°46'0" W) (Fig. 1). The city co-vers an approximate area of 79 km2 and houses 66 133 people (INDEC 2010). The weather is temperate with a mean daily temperature of about 19.5°C and approximate annual rainfall of 990.4 mm (National Meteorological Services, http://www. smn.gov.ar/). The city is bordered by the Salado River at the east. The topography is flat. The major native vegetation types in Santo Tomé are included in the Paranaen-se (Interior Atlantic Forest) and Espinal phytogeo-graphic province confluence. The area is strongly influenced by the flood valley of the Paraná River, which is composed of subtropical wet forest and ga-llery forest. The most abundant species in such area are Salix humboldtiana, Tessaria integrifolia, Nectandra angustifolia, Albizzia inundata, Erythrina crista-galli and different types of flooded savannahs and wetlands (rivers, streams, and ponds) (Cabrera 1994, Arzamendia and Giraudo 2004).
Urban land use types. Firstly, we identified urban land uses with different levels of urbanization in the city of Santo Tomé by visual inspection of satellite images available on Google Earth and our knowledge about the urban matrix. The visual inspection of sate-llite images allowed us to detect potential differences in habitat structure according to the coverage of built surface, vegetation, unpaved soil, and water. Thus, we decided to analyze the following urban land uses which correspond to a decreasing level of urbaniza-tion: (1) the centre of the city, (2) the suburban sector, (3) the peri-urban sector, (4) small urban parks, (5) the military base, (6) the riverside walkway and (7) the urban reserve (Blair 1996, Filloy et al. 2019). The centre of the city consisted of commercial and adminis-trative areas where buildings predominate, and green spaces are scarce. The suburban sector was characte-rized by modérate to high-density, single-family hou-sing within the urban matrix, with lawned sidewalks, yards, and paved roads. The peri-urban sector was located on the boundaries of the city, composed of single-family housing with yards and unpaved roads, near rural zones and green areas. Small urban parks were represented by public green areas smaller than 1.5 ha. The Santo Tomé’s riverside walkway was com-posed of detached houses located within the urban matrix, with lawned sidewalks, yards, and wooded streets. The military base consisted of a large green area with exotic and native vegetation surrounded by urbanized areas. Finally, we selected an urban natural reserve from a locality near to the city of Santo Tomé as a minimally managed site composed of typi-cal habitats from the Paraná valley with native vegeta-tion (D’Angelo et al. 1998, Medrano 2005).
We assessed differences in habitat structure and in the intensity of human activity among the urban land uses. We used the Photoshop CS6 (Adobe) software and the same Google Earth satellite image of each transect to estimate the percentage coverage of built surface, vegetation, unpaved soil, and water as in Gilbert and Butt (2009). The coverage of each variable was confirmed during bird counts. We also me-asured the intensity of human activity represented by (1) vehicle traffic, measured as the number of passing cars per min and (2) pedestrian activity, measured as the number of passing pedestrians per min. We used the Kruskall-Wallis test to find differences between at least one of the pairs of urban land uses.
Bird surveys. We performed bird surveys from Fe-bruary to June 2015. At each urban land use type, we located 10 transects of 100 m long and 50 m wide se-parated by at least 150 m (Fig. 1). Each transect was visited twice. One visit corresponded to the breeding season and the other one, to the non-breeding sea-son. The surveys were performed during the first four hours after dawn, on non-windy, not rainy days and on business days. We visited transects randomly at different moments during the morning to avoid bias in bird detection. We registered birds that were seen or heard, and which were perching, eating, walking, or interacting with another bird within the transect area. Additionally, to ensure our survey effort was comparable between urban land uses, we calculated sample completeness by comparing sample coverage within each urban land use following to Chao and Jost (2012) and using the iNEXT package in R (Hsieh et al. 2020).
Bird diversity and abundance. We measured bird di-versity using Hill numbers, or the effective numbers of equally abundant species (Lou and González-Oreja 2012). Hill numbers differ by a parameter q that re-flects their respective sensitivity to the relative frequency of a species with q = 0, q = 1 and q = 2, corres-ponding to species richness, Shannon-Wiener (H’) and Simpson’s indexes of diversity, respectively (Chao et al. 2014). We used iNEXT package to estímate bird diversity (Hsieh et al. 2020) in R software. iNEXT aims to compare diversity estimates for equally-sized sam-ples by interpolating and extrapolating total species richness (Hsieh et al. 2020). Interpolation and extra-polation curves were computed for each urban land use using bird abundance data and 999 bootstraps. We used the maximum number of individuals for each species during the two visits as a measure of bird abundance. We compared the confidence limits between curves at a minimum sample coverage be-tween urban land uses. Non-overlapping confidence areas indicated significant differences in species di-versity between curves (p < 0.05).
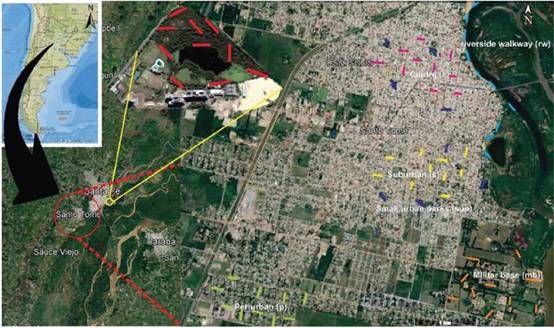
Figure 1. The study area: the city of Santo Tome. Upper left: Argentina. Political boundaries of the province of Santa Fe are shown in light blue. The red circle encompasses the city of Santo Tomé. The yellow circle shows the location of the urban reserve. Transects from different urban land uses are shown in different colours: (pink) the centre, (yellow) the suburban sector, (light green) the peri-urban sector, (blue) the small urban parks, (orange) the military base, (light blue) the riverside walkway and (red) the urban reserve.
We fitted generalized linear models (GLMs) to compare transect values of Hill numbers, total and native abundance, and the ratio of exotic individuals between urban land uses using the glm function from the R software. We used the hillR package (Li 2018) to calculate Hill numbers by transect. Total and native abundance were the mean number of individuals recorded in the two visits for the total species and the native species, respectively. The ratio of exotic individuals was the ratio of the mean number of exotic individuals of the two visits divi-ded the mean number of total individuals. We tested the significance of models by comparing them with null models using likelihood ratio tests (LRT) with the ANOVA function (p < 0.05). To analyse significant differences between urban land uses, we carried out posteriori tests using the emmeans function from emmeans package (Lenth 2022). For species richness, we assumed a Poisson distribution of errors and we checked for over- and sub-dispersion. For the Shannon-Wiener, Simpson index, total and native abundance, and ratio of exotic individuals, we assumed a Gaussian distribution of errors, and ho-moscedasticity and normality were checked. We de-tected and removed outliers for subsequent analysis from the centre and the riverside walkway produced by atypical records of the Brown-chested Martin (Progne tapera) in urban environments. If errors of GLMs had unequal variances, we used generalized least squares (GLS) instead of GLMs using the nlme package (Pinheiro et al. 2022). Spatial auto-correlation of residuals was analysed with the Moran index using “spdep” package from R (Bivand and Wong 2018), and no significant autocorrelation was found (p > 0.05).
Taxonomic composition. We performed non-me-tric Multidimensional Scaling (NMDS) to compare taxonomic composition between urban land uses. NMDS is a multivariate data analysis (Legendre and Legendre 1988) which uses dissimilarity data to or-dinate sites and species in multiple dimensions. We used the Bray-Curtis distance index (Bray and Cur-tis 1957) which takes into account species’ abundan-ces, using the vegan package (Oksanen et al. 2022). We used a Permutation Analysis of Variance (PER-MANOVA) to test differences in species composition between urban land uses. We ran the PERMANOVA with the adonis function from the vegan package.
Results
Urban land uses
Land use characterization. Urban land uses showed significant differences in habitat structure and human disturbance (Fig. 2). The highest percen-tage of built-up area, pedestrians and vehicles were in the centre and suburban sectors, while the lowest percentages, in the military base, riverside walkway and the urban reserve (Built-up area: H = 56.12, p < 0.001, Pedestrians: H = 26.96, p < 0.001, Vehicles: H = 50.18, p < 0.001) (Fig. 2C, E, F). The opposite pattern was observed for the percentage of vegetation and water cover (vegetation cover: H = 53.62, p < 0.001, water cover: H = 27.06, p < 0.001, Fig. 2A, B). The hi-ghest percentages of unpaved soil cover were found in the peri-urban sector and the riverside walkway (Soil cover: H = 35.71, p < 0.001) (Fig. 2D).
Bird diversity. Our survey was enough to record a representative sample of bird assemblages within each urban habitat. The assessment of sample com-pleteness indicated that all samples reached over 80% completeness (89% in urban reserve and > 90% in the rest of urban habitats), allowing for com-parison of bird diversity across habitats (Fig. 3A). A total of 77 bird species were recorded from all ur-ban land uses (Table 1). The most abundant species were the House Sparrow (Passer domesticus, 15.39% of individuals) and the Eared Dove (Zenaida auriculata, 14.8%). The urban centre presented the least number of species (14 species), whereas the urban reserve presented the highest number of species (55 species) (Table 1). The seven urban land uses su-pported different bird composition (Table 1). Based on the three most abundant species in each urban land use, we identified: (i) species that were among the most abundant species in more than one urban land use such as the Eared Dove, the Brown-chested Martin, the House Sparrow and the Monk Parakeet (Myiopsitta monachus) and (ii) species which were among the most abundant species in one urban land use such as the Rufous Hornero (Furnarius rufus) in the suburban sector, the Rufous-bellied Thrush (Turdus rufiventris) and the Gray-breasted Martin (Progne chalybea) in the periurban sector, the Shiny Cowbird (Molothrus bonariensis) and the Guira Cuc-koo (Guiraguira) in the military base, the Rufous-collared Sparrow (Zonotrichia capensis) in the riverside walkway, and the Snowy Egret (Egretta thula) and the Common Gallinule (Gallínula galeata) in the urban reserve. (Table 1).
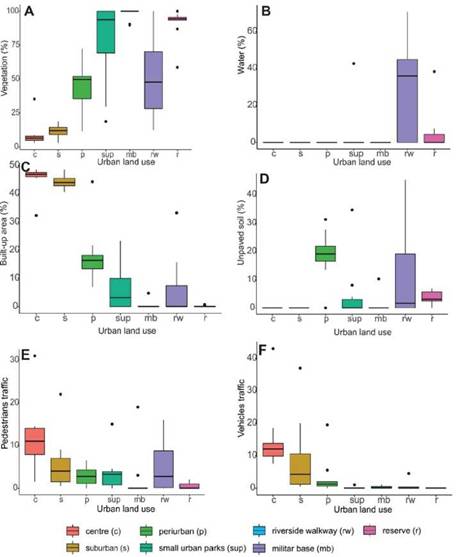
Figure 2. Box-plots of different environmental variables along land uses in Santo Tomé, Argentina. References: the centre of the city (c), the suburban sector (s), the peri-urban sector (p), small urban parks (sup), the military base (mb), the riverside walkway (rw) and the urban reserve (r).
Rarefaction analysis showed significant differences of species diversity between the urban land uses, showing higher diversity in less urbanized land uses (Fig. 3B-D). Such differences were only significant when comparing the urban reserve and the centre with the rest of the urban land uses (Fig. 3B-D). The suburban sector, the peri-urban sector, small urban parks, the military base and the river-side walkway supported intermediate levels of bird diversity (Fig. 3B-D). Estimated species diversity per transect showed similar patterns. Species rich-ness was lower in the city centre, in the suburban and peri-urban sectors than in the urban reserve and the riverside walkway (Fig. 4A). The rest of the urban land uses presented intermediate levels of species richness (Species richness LRT = 105.62, df = 6, p < 0.001) (Fig. 4A). We did not find differences in species diversity (Shannon and Simpson diversity) between the centre of the city, the suburban and the peri-urban sectors, small urban parks and the military base (Fig. 4C, D). Species diversity was higher in the urban reserve and the riverside walkway (Shannon: LRT = 667.4, df = 6, p < 0.001, Simpson: LRT = 336.19, df = 6, p < 0.001) (Table 2, Fig. 4C, D).
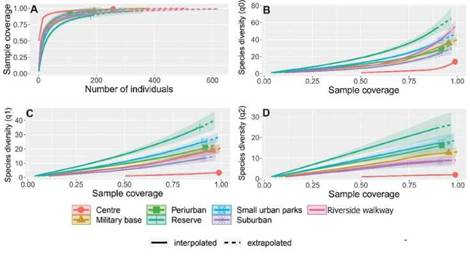
Figure 3. Rarefaction curves showing the sample coverage in relation to the number of individuáis detected (a), and species richness (q = 0, b), the effec-tive number of species for the Shannon-Wiener index (q = 1, c), and the effective number of species for the Simpson index (q = 2, d) in relation to sample coverage for the different land uses in Santo Tomé, Argentina.
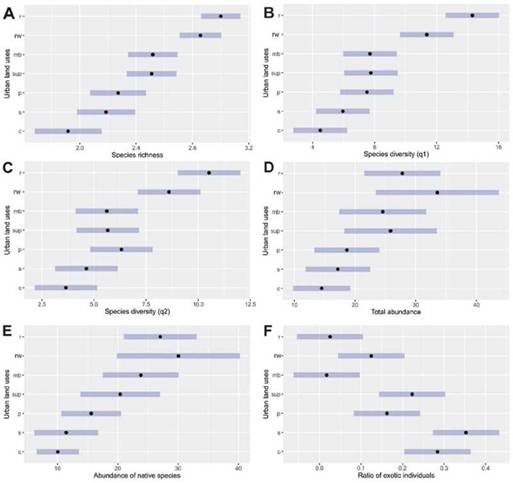
Figure 4. Posthoc differences between land uses for different diversity and abundance valúes. Grey bars indícate confidence intervals. References: the centre of the city (c), the suburban sector (s), the periurban sector (p), small urban parks (sup), the military base (mb), the riverside walkway (rw) and the urban reserve (r).
The total abundance of birds at the centre of the city was lower than in the urban reserve and the ri-verside walkway (LRT = 24.34, df = 6, p < 0.001) (Fig. 4D). However, we did not find significant differences in the rest of pairwise comparisons (Fig. 4D; Table 1). The abundance of native species in the most urba-nized land uses (the centre of the city, the suburban and peri-urban sectors) was lower than in the urban reserve (LRT = 37.55, df = 6, p < 0.001) (Fig. 4E). The rest of urban land uses presented intermediate levels of abundances of native species (Fig. 4E). The ratio of exotic species was lower in the urban reserve and the riverside walkway than in the centre of the city (LRT = 0.95, df = 6, p < 0.001) (Fig. 4F). The rest of urban land uses presented intermediate levels of ratio of exotic species (Fig. 4F).
Bird composition. The PERMANOVA showed signifi-cant differences of bird composition between land use types (F = 4.47, p = 0.001, r2 = 0.30). The NMDS (Stress = 0.20) showed distinct bird assemblages across urban land uses (Fig. 5). The House Sparrow, the Rock Dove (Columba livia), the Chalk-browed Mockingbird (Mimus saturninas) and the Rufous Hornero were more associa-ted to the centre and residential land uses (the suburban and peri-urban sectors). The Grayish Saltator (Saltator coerulescens), the White-barred Piculet (Picumnus cirra-tus), the Wattled Jacana (Jacana jacana), the White-ban-ded Mockingbird (Mimus triurus) and the Greater Thor-nbird (Phacellodomus ruber) were more associated to the urban reserve. The Rufous-collared Sparrow, the House Wren (Troglodytes aedon), the Picui Ground Dove (Columbina picui) and the Great Kiskadee (Pitangus sulphuratus) were more associated to moderately urbanized land uses such as the suburban and the periurban sectors and the small urban parks. The Masked Gnatcatcher (Polioptila dumicola), the Rufous-bellied Thrush, the Grayish Baywing (Agelaioides badius) and the Sayaca Tanager (Thraupis sayaca) were more associated to the riverside walkway and the urban reserve. The Campo Flicker (Colaptes campestris), the Green-barred Woodpecker (Co-laptes melanochloros), the Picazuro Pigeon (Patagioenaspi-cazuro), the Spot-winged Pigeon (Patagioenas maculosa), the Monk Parakeet and the Guira Cuckoo were more associated to the military base (Fig. 5).
Figure 5. Non-metric multidimensional scaling showing the relationship of land uses and bird species in Santo Tomé, Argentina. Large circles indicate the centroid for each land use, whereas small circles indicate each transect. References: the centre of the city (c), the suburban sector (s), the peri-urban sector (p), small urban parks (sup), the military base (mb), the riverside walkway (rw) and the urban reserve (r).
discussionThe results obtained showed that bird diversity and abundance declined with increasing urbanization. Moreover, bird composition showed significant changes between land uses, with a dominance of exotic species in the most urbanized areas. In the city of Santo Tomé, bird species diversity and abundance decreased from highly to scarcely urbanized land uses, a pattern seen in many studies in the northern hemisphere (e.g. Clergeau et al. 2006, McKinney 2008, Vignoli et al. 2013) and within the neotropics (e.g. Ortega-Álvarez and MacGregor-Fors 2009, Leveau et al. 2017, Escobar-Ibáñez et al. 2020). Residential land uses supported similar bird diversity to the city centre and some recreational land uses such as the riverside walkway supported similar bird diversity to the urban reserve. According to some medium-sized cities in the Pampean region, the abundance of total bird species and native bird species decreased from highly to scarcely urbanized land uses (Garaffa et al. 2009). According to previous findings worldwide, bird assemblages in the city of Santo Tomé were composed mainly by widely distributed native species (Leveau and Leveau 2004, Juri and Chani 2009). However, two exotic bird species were identified: the House Sparrow and the Rock Dove. Contrary to previous findings, the first concentrated at the more urbanized land uses whereas the latter was represented by a few individuals at highly and moderately urbanized land uses. Our knowledge of the effect of urbanization is still incipient and more re-search is needed to understand the underlying ecologi-cal processes that shape the taxonomic bird diversity in the city of Santo Tomé. However, our research provides valuable information and constitutes a framework for future studies.
Cities are heterogeneous landscapes and bird as-semblages respond accordingly (e.g. Juri and Chani 2005, Leveau et al. 2017). Using the gradient approach to study these variations, in humid forests, the bird diversity often increases from highly urbanized land uses (commercial and administrative areas) towards natural protected/unprotected areas (Ortega-Álvarez and Mac-Gregor-Fors 2009, Filloy et al. 2019, Escobar-Ibáñez et al. 2020). Although we reported an increase in the bird diversity from highly to scarcely urbanized land uses, we observed that some moderately urbanized land uses, such as the riverside walkway, supported similar bird diversity to the reserve. In Argentina, the first was reported in a previous study performed in an Argenti-nian city but in an urban-rural gradient (e.g. Leveau et al. 2017). High levels of bird diversity in moderately urbanized land uses are often associated with a high diversity and abundance of habitat and resources available to birds (Blair 1996, Hurlbert 2004, Leveau 2013, 2019). In moderately urbanized land uses, humans create public and private urban green spaces with vegetation strata: lawn, shrubs, and trees. The presence of a variety of shrub and tree species usually provides a diversity of flora and fruit resources for birds (Frohlich and Ciach 2020, Curzel and Leveau 2021, Lerman et al. 2021). In addition, preferences for colourful and showy plants which are easy to maintain often influence vegetation management and habitat heterogeneity (Avolio et al. 2018, Cavender-Bares et al. 2020). The introduction of watering regimes, fertilizers and regular maintenance in these urban areas may also increase primary produc-tivity (Blair 1996, Lepczyk et al. 2004, Cavender-Bares et al. 2020). The mixture of non-vegetated and vegeta-ted features at moderately urbanized land uses may increase the amount of edge habitat available, allowing a higher species richness than in highly urbanized land uses (Blair and Johnson 2008, MacGregor-Fors and Schondube 2011, Leveau 2013). On the other hand, the high bird diversity observed in sparsely urbanized land uses (the urban reserve) may be associated to both: (i) the highest levels of primary productivity and habitat diversity produced by continuous coverage of highly heterogeneous habitats dominated by native vegeta-tion (Garaffa et al. 2009, Leveau et al. 2017, Filloy et al. 2019), and (2) the lowest levels of human activity since the replacement of green areas by impervious surface, the human population density and vehicle traffic and noise act as selective pressures to species in more ur-banized land uses (Ortega-Álvarez and MacGregor-Fors 2009, Leveau et al. 2017, da Silva et al. 2021). Therefore, our results suggest that similar levels of bird diversity to protected/unprotected lands at moderately urbanized land uses are possible. However, future studies aimed to disentangle the effect of habitat features across the city are encouraged since it has been shown that habitat structure can shape bird assemblages at different levels of urbanization, even in highly urbanized land uses (Pe-llissier et al. 2012, Curzel et al. 2021).
The total energy available in ecosystems shape the abundance of individuals that they support (Storch et al. 2018). Since primary productivity varies across ur-ban systems, changes in the abundance of individuals in bird assemblages are expected (Chiari et al. 2010, Le-veau 2019, Leveau et al. 2020). In Argentina, changes of total bird abundance along urban/non-urban gradients reported in the literature are variable (Leveau and Le-veau 2004, Garaffa et al. 2009, Leveau and Zuria 2017). In contrast, an increase of the abundance of native bird species towards the non-urban extreme was often reported in Argentinian cities (Garaffa et al. 2009, Leveau 2019). Regarding the total abundance of bird species and of native bird species, our results agree with the findings of Garaffa et al. (2009) and Leveau and Leveau (2004) for some medium size cities given that the most urba-nized land uses (centre and suburban sector) presented lower levels of abundance than the less urbanized land uses (the riverside walkway and the urban reserve). The number of individuals that a habitat can support is often expected to be positively associated to primary produc-tivity (Storch et al. 2018, Leveau 2019). It is important to note that the increase in the abundance of birds from hi-ghly to scarcely urbanized land uses in the city of Santo Tomé was more evident when only native species were considered. Exotic bird species concentrated mainly at highly urbanized land uses (Leveau et al. 2017). The urban exotic bird species are able to thrive in highly urbanized land uses since they are able to feed on discarded food provided by humans and nest on buildings in an environment where the predation pressure is often low (McKinney 2006). These results reinforce the relevance of those urban land uses in which resources for native species are available to favour the abundance of nati-ve species in urban systems. In addition, some gaps in the knowledge of the underlying process that shape the abundance of exotic species in the city of Santo Tomé emerged. In Argentinian cities the increase in the abun-dance of exotic bird species towards the more urbanized sites are mainly represented by two Eurasian species, the House Sparrow and the Rock Dove (Germain et al. 2008, Juri and Chani 2009, Cristaldi et al. 2017, Leveau et al. 2017), even in intermediate size cities (Ramirez et al. 2016, Gorosito and Cueto 2020). The relative low abundance of the Rock Dove in the city of Santo Tomé with respect to nearby cities (Cristaldi et al. 2017) may need further studies. As it is a species that is able to fly the entire distance between Santo Tomé and nearby cities in a single day (Carlen and Munshi-South 2021), its relative low abundance may be associated to histo-rical factors (e.g., the year when the city was colonized by birds) or environmental features of the city of Santo Tomé (e.g. the lack of high-rise buildings) (Hetmañski 2007, Evans et al. 2010, Meller et al. 2012, Przybylska et al. 2012, Tang et al. 2018, Carlen and Munshi-South 2021). This knowledge may contribute to urban plan-ning to regulate populations of exotic species in urban systems.
Bird assemblages within the city of Santo Tomé are composed of a lower number of native species than in surrounding non-urban areas (rural and natural areas) (Rossetti and Giraudo 2003, de la Peña 2013, Frutos et al. 2020). It has been shown that urban systems can fil-ter bird species according to their ecological functions (Croci et al. 2008, Silva et al. 2016, Leveau 2021). For instance, although insectivorous species were often identified across all urban land uses such as the House Wren, the Rufous Hornero and the Great Kiskadee, other insectivorous species such as the White-barred Piculet, the Great Antshrike (Taraba major) and the Black-capped Warbling Finch (Microspingus melanoleucus) appeared only in the urban reserve, which was dominated by nati-ve vegetation (Cristaldi et al. 2017). In addition, the suc-cess of colonizing urban systems may also be associated with the abundance of species in non-urban areas (e.g. Leveau et al. 2022). In this sense, species present in the city of Santo Tomé are among the most abundant spe-cies in the surrounding non-urban areas as it was reported in previous studies (Rossetti and Giraudo 2003, de la Peña 2013, Frutos et al. 2020). Therefore, future studies should shed light on the underlying processes that sha-pe the urban bird assemblage in the city of Santo Tomé in order to enhance the occurrence of those native species that seem to avoid urban systems and constraint population dynamics of exotic bird species.
final remarksAlthough it has been widely shown that large urban areas have a significant effect on bird assemblages, our results suggest that intermediate size cities like Santo Tomé can also have a significant effect. In our study, bird species diversity and abundance increased from highly to scarcely urbanized land uses. Exotic species concentrated their abundance on the most urbanized land uses. These declines in bird diversity suggest that several native bird species that inhabit the surrounding non-urban habitats could not thrive in the city. Howe-ver, we observed that most of urban land uses can still retain native species and some of them may enhance bird diversity and contribute to the heterogeneity of bird assemblage composition. In this sense, high levels of species diversity and heterogeneity of bird composition were associated to urban land uses with natural resour-ces according to the ecological requirement of species, relative low levels of human disturbance and vegetation management for conservation or aesthetic purposes. Given that the city of Santo Tomé is expected to grow, our results should serve as a base knowledge for an urban planning aimed to enhance the diversity and hete-rogeneity of urban bird assemblages.
Table 1. Máximum abundance of species recorded in each urban land use type throughout the study period.
Table 2. Generalized linear model and Generalized least squares model results showing differences in Species richness, Shannon-Wiener and Simpson index, total abundance, abundance of native spe-cies, ratio of exotic individuals between urban land uses of Santo Tomé, Argentina. z/t indicates z or t tests. Reserve is in the intercept.
Acknowledgments
The following supported the work: CONICET (PIP 2011-0355), Universidad Nacional del Litoral (CAID-2016-UNL), ANPCYT (PICT 2016-1415 and PICT-2013-2203 FONCYT). We thank the Consejo Nacional de Investigaciones Científicas y Técnicas and the Instituto Nacional de Limnología (CONICET- UNL) which facilitated our work.
Recibido: Recibido: 21 de julio de 2022
Aceptado: 24 de noviembre de 2022

