











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkLa evidencia actual sugiere que el sistema climático está cambiando e incrementando la frecuencia de eventos meteorológicos extremos (e.g., huracanes, tormentas, inundaciones), que se caracterizan por su intensidad en un periodo corto de tiempo y su rareza (van de Pol et al. 2010, Bailey y van de Pol 2016, Ummenhofer y Meehl 2017). Los eventos extremos tienen un gran impacto en los sistemas biológicos al influir en la biología reproductiva de las especies e incrementar sus tasas de mortalidad (Moreno y M0ller 2011). Estos efectos, pueden producirse tanto de forma directa como indirecta a través de impactos en las poblaciones y sus hábitats (Dinsmore y Farnsworth 2006, Newton 2007, Martin y Maron 2012).
Las tormentas severas son eventos meteorológicos extremos que tienen la capacidad de producir daños materiales importantes y/o muertes. Generalmente, las tormentas severas se conforman por lluvias intensas y vientos fuertes, pueden producir granizo, rayos, inundaciones repentinas e, incluso, tornados (Prieto-Gonzales et al. 2014). Las aves son uno de los taxones de vertebrados más afectado por eventos de mortandad masivos producidos por tormentas severas, las cuales han incrementado su frecuencia en las últimas décadas (Fey et al. 2015a, Yang et al. 2021). Uno de los parámetros poblacionales más afectados es la supervivencia, asociada a la intensidad del evento (Newton 1998, Sther y Bakke 2000, Ju-lliard et al. 2004). Las consecuencias implican una reducción del tamaño de las poblaciones reproductoras que podrían tener efectos profundos en la dinámica poblacional de especies de comportamiento gregario (S^ther y Bakke 2000, Dionne et al. 2008, Zuberogoi-tia et al. 2016).
El crecimiento de la población humana ha propiciado que los ecosistemas naturales sean reemplazados por paisajes urbanos (Bolund y Hunhammar 1999). Varias especies de aves han colonizado las ciudades y algunas de ellas son más exitosas en ambientes urbanos que en sus hábitats naturales (Luniak 2004). Las aves urbanas muestran mayor plasticidad ecológica, fisiológica y comportamental que sus congéneres de ambientes rurales, lo cual les permite tolerar una mayor amplitud en las condiciones ambientales incluyendo hábitats alterados o degradados (Bonier et al. 2007). La cobertura vegetal y la superficie de los espacios verdes en las ciudades son factores claves que determinan la abundancia y composición de las comunidades de aves urbanas (Melles et al. 2003, Faggi y Perepelizin 2006, Moller et al. 2012). Se ha observado que en espacios verdes con mayor diversidad de especies vegetales se incrementa la riqueza de aves, ya que se benefician de los recursos que ofrecen las ciudades (Sovrano 2017, Guachamín-Paladines 2021). Los árboles de gran tamaño, por ejemplo, son recursos muy utilizados para pernoctar en las urbes (Leveau y Leveau 2004, Fey et al. 2015b).
El paisaje urbano presenta un mosaico de espacios verdes y aislados inmersos en una matriz de estructuras antropogénicas (Niemela 1999). Estos espacios verdes, consisten en parches de vegetación de diferente tamaño, forma, disposición, estructura, composición, diversidad, y calidad de hábitat para la avifauna urbana (Davis et al. 2014, Wang et al. 2017). Los parches representan un suministro de recursos esenciales para numerosas especies de aves, ya que aumentan las probabilidades de que las mismas encuentren hábitats adecuados para alimentación, nidi-ficación y refugio (Sulaiman et al. 2013). Los estudios de ecología urbana han descrito la diversidad de aves y los requerimientos de hábitat en distintos tipos de parches (Bolund y Hunhammar 1999, Barbosa et al. 2011, Oliver et al. 2011, Pineda-López et al. 2013, Strohbach et al. 2013, Nielsen et al. 2014). Estos parches de vegetación son muy importantes porque ayudan a mantener una riqueza de especies de diversos taxones y contribuyen a la calidad ambiental de los ecosistemas urbanos (Bolund y Hunhammar 1999, Knapp et al. 2008, Nielsen et al. 2014).
Actualmente existe escasa información sobre la mortalidad de aves que producen las tormentas en Sudamérica. Además, la información existente suele ser anecdótica o incompleta (Marzluff 2017). Los trabajos apuntan en general a áreas costeras del hemisferio norte, evaluando el efecto de huracanes sobre especies migratorias (Tavares et al. 2020). Los objetivos de este estudio fueron: i) cuantificar individuos muertos de aves afectadas por una tormenta severa en la ciudad de Tulumaya, Mendoza, Argentina, e ii) identificar dormideros comunales para estimar la mortalidad de individuos del área urbana impactada por la tormenta. Debido a que se proyecta un incremento en los eventos meteorológicos extremos como consecuencia del cambio climático (Grimm 2011, Cai et al. 2014), se espera que la supervivencia de las aves urbanas a dichos eventos esté condicionada por la calidad del hábitat y sus rasgos de historias de vida, en particular la etología y demografía de las especies (Fernández-Juricic 2004, Dionne et al. 2008, Zubero-goitia et al. 2016).
métodosÁrea de estudio
El estudio se desarrolló en la ciudad de Tulumaya, departamento de Lavalle, provincia de Mendoza, Argentina (Fig. 1; 32°43’18’’S 68°35’44’’O). La ciudad se encuentra a 600 msnm, en la ecorregión del Monte de Llanuras y Mesetas (Oyarzabal et al. 2018). El clima es árido a semiárido, con temperaturas medias de 22°C en verano y 8°C en invierno (González-Loyarte et al. 2009). Las precipitaciones son de régimen estival con una media de 200 mm anuales (Norte 2000). El 4% de la superficie de la provincia está cubierta por oasis (zonas cultivadas por riego artificial). Tulumaya se encuentra inserta en el oasis norte, alimentado por los ríos Mendoza y Tunuyán (Balter 2015). Tuluma-ya es el distrito más poblado de Lavalle, con cerca de 8000 habitantes (Griffone 2019).
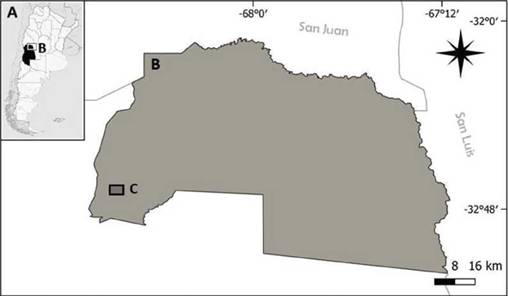
Figura 1. Ubicación del área de estudio en la ciudad de Tulumaya, provincia de Mendoza, Argentina. En el polígono delimitado (c) puede observarse los parches forestales (verde) identificados mediante teledetección.
En la ciudad, se delimitó un polígono de 154 ha (Fig. 1) que comprende el casco céntrico del área urbana, excluyendo de esta manera la matriz agrícola y de monte nativo de los alrededores. En los espacios verdes urbanos predominan las especies arbóreas exóticas (Martínez-Carretero 2022). El desarrollo de dichas especies en una región con escasas precipitaciones, fue posible debido al sistema de regadío por acequias y canales que trazan la ciudad, de forma análoga al resto de los asentamientos de la provincia de Mendoza (Prieto et al. 2008).
Descripción de la tormenta severa
La tormenta, cuyos efectos se analizan en el presente trabajo, se inició a las 20:57 h y finalizó a las 21:10 h el día 16 de diciembre de 2021 (obs. pers. Ta-llei). La misma presentó precipitaciones en forma de agua y granizo, y ráfagas de viento huracanado (obs. pers. Tallei). Según la clasificación de la intensidad de los vientos del Sistema Meteorológico Nacional, la velocidad del viento podría haber superado los 89 km/h (temporal muy fuerte) ya que entre las consecuencias se produjeron daños en edificios y caídas de árboles (Anexo 1). Las precipitaciones fueron de 33.79 mm/ día y se registró una temperatura media de 17.7°C (NASA).
Colecta de datos
Mortalidad de aves. Se realizó una parcela de 100 x 100 m en un parche forestal a la mañana siguiente de la tormenta (17 de diciembre de 2021) para registrar individuos muertos de aves (Anexo 2). Se cuanti-ficaron todos los individuos que se encontraron en el suelo y se identificaron a nivel de especie. El parche forestal tenía una superficie total de 1.88 ha, una riqueza de 14 especies arbóreas y una densidad de 92.5 individuos/ha de árboles.
Identificación de parches forestales. Se utilizó una imagen satelital del sensor Sentinel-2, European Space Agency, correspondiente a la fecha del evento meteorológico (diciembre 2021). Se aplicó un índice espectral denominado índice de clorofila verde (GCI = NIR / GREEN - 1) para identificar los parches forestales. GCI se utiliza para estimar el contenido de clorofila de las hojas, reflejando el estado fisiológico de las plantas y siendo útil para discriminar tipos de vegetación (Sarker et al. 2021). Se realizó una clasificación no supervisada, ya que esta técnica permite definir clases considerando la similitud entre píxeles y calcular la superficie (Paruelo 2014). Se definieron tres tipos de clases o coberturas: parche forestal, espacio verde general, y urbana (infraestructura y área edificada). El suelo > 0.1 ha de cobertura de árboles se clasificó como parche forestal (Wang et al. 2017). El suelo con la combinación de distintos elementos vegetales (i.e., herbáceas, arbustos, y árboles) y suelo desnudo, se clasificó como espacio verde general. En cada parche forestal, se procedió a validar dicha clasificación en el terreno y a cuantificar la abundancia absoluta y composición de las especies arbóreas > 20 cm de DAP (diámetro a la altura del pecho). Las imágenes fueron procesadas utilizando las herramientas del sistema de información geográfica del software Quantum GIS versión 3.16.4 (QGIS Development Team 2021).
Dormideros de aves. Una vez identificados los parches forestales mediante teledetección, se procedió a validarlos como dormideros. Se definió como dormideros comunales a aquellos parches forestales que fueron usados para pernoctar por las especies de aves de interés (i.e., aquellas que fueron registradas muertas por la tormenta: Patagioenas maculosa, Zenaida au-riculata, Myiopsitta monachus, Columbina picui y Passer domesticus), independientemente si eran usados por otras especies de aves, y en los cuales se cuantificó un mínimo de 163 individuos de aves por ha (valor de referencia = valor observado de individuos muertos por la tormenta). Se realizaron cinco revisitas en distintos días (10, 11, 13, 16 y 17) durante el mes de mayo de 2022 a cada parche forestal (visitando el total de parches por día). Los parches fueron recorridos por dos observadores a una velocidad mínima constante a pie, trazando una transecta de longitud variable por 100 m de ancho en el área central de cada parche (Bi-bby et al. 2000). Se identificaron las especies de aves vistas u oídas, y se registró la abundancia de las especies de interés de este estudio por cada árbol (Ralph et al. 1996). Los muestreos se realizaron entre las 18 y 19:30 h, en coincidencia cuando las aves arriban a los dormideros para pernoctar. En cada revisita, se alteró el orden de recorrido de los parches para evitar sesgos en los horarios de llegada de las aves.
Análisis estadísticos
Se estimó el número y superficie de área (ha) de los parches forestales, y de cada tipo de cobertura. Se utilizó un análisis de escalamiento multidimensional no métrico (NMDS) para analizar diferencias en los parches forestales que eran usados como dormideros de aquellos que no. Las ordenaciones se llevaron a cabo utilizando datos cuantitativos (basados en la abundancia de cada especie de árbol) y el índice de Bray-Curtis. Se aplicó un análisis de similitud (ANO-SIM) para evaluar diferencias significativas en la composición y abundancia de árboles entre los parches. Para comparar la diversidad de especies arbóreas, se utilizaron curvas de rarefacción basadas en el número de individuos y la interpolación/extrapolación de los números de Hill (número efectivo de especies; Chao et al. 2014). Éste método, permite analizar una o varias comunidades a partir de la riqueza de especies (q = 0), diversidad de Shannon (q = 1, exponencial de entropía de Shannon) y diversidad de Simpson (q = 2; inversa de la dominancia de Simpson) (Moreno et al. 2017).
Se estimó la proporción de árboles usados por las aves como dormideros y la disponibilidad de los mismos en los parches. Los datos de disponibilidad y uso fueron analizados utilizando el índice de selección de Manly (Wi). Un índice Wi > 1 indica selección a favor de ese recurso, un índice Wi = 1 indica que no hay selección, mientras que un índice Wi < 1 indica evitación o rechazo (Manly et al. 2002). El uso aleatorio o selectivo se evaluó mediante intervalos de confianza del 95%. Debido a que las condiciones de mínima luz al atardecer disminuyen la detectabilidad de las aves generando sesgos en los conteos (Bibby et al. 2000, Esquivel y Peris 2008), se utilizaron datos de presencia/ausencia para evaluar el uso y la selección de los árboles como dormideros a nivel de ensamble de aves.
La estimación de la abundancia de cada especie de ave se expresó como individuos por hectárea (ind/ ha). Se extrapoló el número de individuos muertos a la superficie total de los dormideros identificados. Los análisis fueron realizados mediante los paquetes stats, tidyverse, vegan, iNEXT y ggplot2 en el software R versión 4.0.1 (R Development Core Team 2020).
resultadosSe registraron 163 individuos muertos de cinco especies de aves (Tabla 1). El 83.4% de los individuos afectados correspondieron a Paloma Manchada (Pata-gioenas maculosa), el 8.6% a Torcaza (Zenaida auricula-ta), el 5.5% a Cotorra (Myiopsitta monachus), el 1.9% a Torcacita Picuí (Columbinapicui) y por último el 0.6% a Gorrión (Passer domesticus). La mortalidad estimada fue de 628 individuos para el total de los dormideros (3.85 ha) identificados en el área de estudio.
Se identificaron en total 13 parches forestales los cuales representaron el 3.68% de cobertura del área de estudio. La cobertura urbana y la cobertura espacio verde general, representaron el 65.09% y el 31.23% respectivamente. Del total de los parches forestales, siete resultaron dormideros de aves y representaron el 67.81% de la superficie; mientras que seis parches no resultaron dormideros. En los dormideros se registraron en total 996 individuos de las especies de aves de interés para este estudio, de los cuales el 91.06% fueron palomas manchadas, 4.92% torcazas, 2.51% cotorras y 1.51% gorriones.
En cuanto a la vegetación, se registraron en total 595 individuos de 19 especies arbóreas, y la composición y abundancia de los parches entre tipo de uso (dormidero vs no dormidero) fue diferente (Fig. 2; ANOSIM, R = 0.28; P = 0.02). Los dormideros tuvieron una superficie de 0.52 ± 0.23 ha (media ± EE) y una densidad de árboles de 126.92 ± 29.37 ind/ha. Los parches que no resultaron dormideros tuvieron una superficie de 0.34 ± 0.06 ha y una densidad de árboles de 111.17 ± 27.38 ind/ha. La diversidad de árboles fue mayor en los parches que resultaron dormideros para los índices 0D = 19 ± 3.57 (IC 19-27) y 1D = 8.27 ± 0.46 (IC 7.58-9.37), en comparación a 0D = 10 ± 1.42 (IC 10.2-15.77) y 1D = 5.18 ± 0.38 (IC 4.63-6.11) de los parches que no lo fueron (Fig. 3).
Tabla 1. Densidad de individuos muertos de aves registrados y estimados a partir de los efectos de una tormenta severa ocurrida en la ciudad de Tulumaya, Mendoza, Argentina.
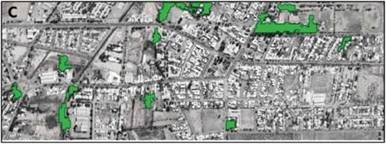
Figura 2. Diagrama de ordenamiento mediante un análisis de escalamiento multidimensional no métrico (NMDS) de la composición y abundancia de especies de árboles de los parches forestales clasificados según si resultaron dormideros de aves o no, en la ciudad de Tulumaya, Mendoza, Argentina. Referencias: euca = Eucalyptusspp. (Eucalipto); plat = Platanushispanica (Plátano); pinu = Pinusspp. (Pino); palb = Populus alba (Álamo blanco); pnig = Populus nigra (Álamo negro); sali = Salix babylonica (Sauce); schi = Schinus molle (Aguaribay); moru = Morus alba (Morera); ulmu = Ulmus spp. (Olmo); meli = Melia azedarach (Paraíso); olea = Olea europaea (Olivo); tama = Tamarix galica (Tamarindo); pros = Prosopis spp. (Algarrobo); cupr = Cupressus spp. (Ciprés); ficu = Ficus carica (Higuera); brac = Brachychitonpopulneus (Brachichito); tipu = Tipuana tipu (Tipa); sene = Parasenegalia visco (Acacia); pdel = Populus deltoides (Álamo Carolino).
Diez especies de arboles fueron usadas cómo dormideros (Fig. 4). Las especies que presentaron mayores proporciones de disponibilidad en los parches forestales fueron Eucalipto (Eucalyptus spp.), Álamo Blanco (Populus alba) y Pino (Pinus spp.). Las especies de árboles que fueron seleccionadas como dormideros fueron Platanus hispánica (Wi = 3.15 [IC 1.759.41]), Ulmusspp. (Wi = 3.25 [IC 1.28-8.47]) y Morusalba (Wi = 2.95 [IC 1.26-8.13]); mientras que se observó un rechazo por las especies Schinus molle (Wi = 0.26 [IC 0.13-0.98]) y Pino (Wi = 0.11 [IC 0-0.35]). Por otra parte, el resto de las especies de árboles usadas no resultaron seleccionadas ni rechazadas (Wi = 1; Fig. 4).
discusiónEste es el primer estudio que evidencia el efecto letal de una tormenta severa sobre especies de aves urbanas en Sudamérica (Marzluff 2017). De acuerdo a nuestros resultados, se registraron cinco especies de aves afectadas, las cuales se caracterizan por estar adaptadas a ambientes urbanos (especies sinantró-picas) y presentan una alta abundancia en ciudades (Leveau 2013a). Se encontró un uso diferencial de los parches forestales y especies de árboles para pernoctar, y esto podría deberse al tamaño del parche y la diversidad de especies arbóreas que presentaron los dormideros (Evans et al. 2009, Chang y Lee 2016). Los eventos de mortandad en masa producidos por las tormentas severas, podrían afectar la dinámica poblacional de ciertas especies de aves que ocupan determinados hábitats en una ciudad (Newton 2007). La cobertura de parches forestales mostró una superficie relativamente baja (Juri y Chani 2005, Leveau 2013b), y esto debería tenerse en cuenta en el manejo y diseño de la configuración espacial del paisaje urbano, para contribuir a la supervivencia de las aves y evitar la pérdida de biodiversidad (Hansen y Urban 1992, Frank y Wissel 1998).
Figura 3. Diversidad de especies de árboles en parches forestales que resultaron dormideros de aves y aquellos que no, en términos de riqueza (q = 0), especies comunes (q = 1) y especies dominantes (q = 2) en la ciudad de Tulumaya, Mendoza, Argentina. Se muestra una porción de la curva de rarefacción interpolada (línea continua), una porción extrapolada (línea discontinua) y los intervalos de confianza al 95%.

Figura 4. Proporciones de uso y disponibilidad (eje izquierdo) e índice de selección de Manly (Wi; eje derecho) de las especies de árboles usadas como dormideros de aves en la ciudad de Tulumaya, Mendoza, Argentina. Se muestra Wi con un punto y sus intervalos de confianza del 95%. Referencias: Eucalipto = Eucalyptus spp.; Plátano = Platanus hispanica; Pino = Pinus spp.; Álamo blanco = Populus alba; Álamo negro = Populus nigra; Aguaribay = Schinus molle; Morera = Morus alba; Olmo = Ulmus spp.; Paraíso = Melia azedarach; Tamarindo = Tamarixgalica.
Los parches usados como dormideros resultaron con mayor diversidad de especies arbóreas, y la complejidad estructural y composición de la vegetación son factores determinantes de la ocupación de especies de aves en áreas urbanas (Melles et al. 2003, Fer-nández-Juricic 2004, Barbosa et al. 2011). Las especies seleccionadas para pernoctar (i.e. Plátano, Olmo, y Morera) son árboles exóticos de gran tamaño con cierta similitud estructural como el desarrollo de una copa amplia irregular en comparación a otras especies como Pinus spp. o Populus spp. (Martínez et al. 2013, Martínez et al. 2014, Martínez-Carretero 2022). Las características de esta arquitectura, influye en el número de perchas lo cual favorece una mayor disponibilidad de refugios para las aves (McClanahan y Wolfe 1993, Shiels y Walker 2003). Estos árboles, además, presentan mayor área foliar en comparación a especies de hojas compuestas como los algarrobos y aguaribays, o especies de hojas aciculares como los pinos (Martínez et al. 2014, Martínez-Carretero 2022). Dichos atributos estructurales, podrían brindar mayor protección contra condiciones climáticas adversas (precipitaciones, vientos, etc.) y depredadores como los gatos domésticos (Evans et al. 2009, Girini et al. 2014, Loss et al. 2015).
La Paloma Manchada y la Cotorra han colonizado numerosas ciudades y utilizan los árboles de gran porte para descansar y reproducirse, entre ellos Eucaliptos y Plátanos (Volpe y Aramburú 2011, Rodrí-guez-Maturino et al. 2018). Por otro lado, el Gorrión usa oquedades para anidar, e incluso de árboles de Plátano, Eucalipto y Olmo, y en zonas donde la disponibilidad de cavidades es limitada, puede ubicar sus nidos en el ramaje de los árboles o en nidos de otras especies de aves como las cotorras (Murgui 2011, Hernández-Brito et al. 2021). Si bien el ensamble de aves no mostró selección hacía el Eucalipto, este árbol fue la especie más abundante y usada para pernoctar. De esta manera, en relación a la cobertura del follaje, podría proporcionar refugios estables durante todo el año para las aves ya que es una especie perenne (Pérez 2004).
Las especies que perecieron por la tormenta, son aves granívoras, omnívoras, y oportunistas, de forma análoga a varias urbes del Neotrópico (Leveau 2013b, Montenegro-Pazmiño 2015). Además, las especies registradas se caracterizan por presentar poblaciones densas y estables en estos ambientes (Juri y Chani 2005). El tamaño poblacional les permitiría una mayor resiliencia y recuperación de la estructura comunitaria ante eventos meteorológicos extremos (Newton 2007). Las palomas podrían tener una alta mortandad debido a su elevada abundancia en la zona y su ecología comportamental (Calamari et al. 2011). Las mismas, suelen perchar en el canopeo de los árboles (obs. pers. Tallei y Benavidez), comportamiento que podría exponerlas al impacto del granizo y arrastre por vientos, en comparación a las aves medianas o pequeñas que se refugian debajo del dosel, a menor altura o en huecos (Fernández-Maldonado et al. 2017). A pesar de que las cotorras tienen mayor protección en sus nidos cerrados, ante vientos intensos los mismos son susceptibles al desprendimiento y caída debido a su gran tamaño y peso (Volpe y Aramburú 2011). Se ha encontrado que la mortalidad de las aves se incrementa en relación a la frecuencia de los eventos de vientos extremos (Louzao et al. 2019).
El menor registro de individuos muertos de especies de aves de menor tamaño como las torcacitas y gorriones, puede deberse a que los cuerpos pequeños resultan más fáciles de remover por animales de las urbes como perros, gatos, comadrejas (Didelphis albiventris), y ratas (Loss et al. 2015). Al atardecer, la de-tectabilidad de las especies de menor tamaño corporal en los dormideros, también puede resultar menor por las condiciones mínimas de luz (Bibby et al. 2000). Estas limitaciones, junto al restringido esfuerzo de muestreo, pueden sesgar las estimaciones de mortalidad de las especies como también del uso de hábitat en este trabajo (Husby 2016). Por otra parte, los dormideros pueden variar estacionalmente en relación a la fenología de la vegetación, al comportamiento de los ensambles de aves, a los hábitos de descanso y a la selección del hábitat que puede diferir entre especies (Pérez 2004, Jirinec et al. 2016, Stanley et al. 2021). Sin embargo, los trabajos que abordan los efectos de los eventos meteorológicos extremos suelen presentar varias limitaciones en el diseño de muestreo, y esto principalmente se encuentra asociado a la baja ocurrencia de los mismos y a las dificultades que conlleva cuantificar sus efectos en el momento en el que ocurren (Bailey y van de Pol 2016, Ummenhofer y Meehl 2017).
Aunque aún no hay mucho consenso sobre las proyecciones y modelos de los eventos climáticos extremos en Sudamérica (Grimm 2011), los mismos pueden representar un factor significativo de pérdida tanto de individuos como de hábitat para las aves. Como consecuencia del evento del presente estudio, Defensa Civil Mendoza intervino en más de 170 situaciones, entre ellas más de 100 árboles y ramas caídas en la ciudad de Tulumaya. Por otro lado, las poblaciones de numerosas especies de animales pueden ser resistentes al impacto de eventos extremos debido a la baja frecuencia de ocurrencia (Newton 2007). Sin embargo, dado que se proyecta un incremento en la frecuencia y severidad de estos eventos como consecuencia del cambio climático (Parry et al. 2007, Cai et al. 2014), es necesario comprender y caracterizar la respuesta de las poblaciones ante eventos extremos no solo para evaluar los impactos, sino para orientar los planes de mitigación y ordenación de las áreas verdes urbanas (Berget 2006).
Nuestros resultados evidencian que la tormenta afectó a cinco especies de aves urbanas, y en este sentido la supervivencia podría depender de sus rasgos de historias de vida (Dionne et al. 2008, Zuberogoi-tia et al. 2016) y las características estructurales y de composición que presentan sus hábitats (Evans et al. 2009). Debido a los escasos estudios en el Neotrópico sobre el efecto de las tormentas severas en las aves, identificar los impactos en las especies y sus hábitats podría contribuir a dilucidar los mecanismos que confieren resiliencia a las poblaciones (Ummenhofer y Meehl 2017). Esto es importante para planificar intervenciones sobre los espacios en la gestión del riesgo urbano (Bonier et al. 2007), ya que los espacios verdes no solo favorecen a la biodiversidad, sino que tienen efectos positivos en la salud humana (Chiesura 2004, Mitchell y Popham 2008, Goddard et al. 2017, Wang et al. 2017).
agradecimientos
Agradecemos por colaborar con el trabajo de campo a Eva Tallei. Este trabajo fue realizado con fondos del Grupo Águila Coronada (Naturalistas de Mendoza). ET y AB son becarios postdoctorales de CONICET.
Recibido: 20 de julio de 2022
Aceptado: 20 de diciembre de 2022

