











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkLa urbanización es una de las fuerzas de transformación más intensa de los ambientes naturales. A pesar del impacto negativo generalizado que ejerce este proceso sobre la fauna silvestre (Garitano Zavala y Gismondi 2003, Klem 2008, Loss et al. 2013, Sol et al. 2014) muchas aves han logrado adaptarse a vivir en ambientes antropizados e incluso han prosperado de forma exitosa haciendo uso de los recursos que ofrecen estos entornos modificados (Marzluff 2017). Como consecuencia, los centros urbanos proporcionan un hábitat con desafíos y oportunidades para distintas especies de aves silvestres (Rebolo et al. 2017).
Los Psittaciformes ocupan diversos ecosistemas, desde selvas tropicales y bosques templados hasta pastizales, sabanas y arbustales, en distintos continentes, aunque la mayor diversidad del grupo se concentra en América del Sur y Australasia (Forshaw y Cooper 1989, Toft y Wright 2015). Un número importante de especies del orden han colonizado ambientes urbanos dentro y fuera de sus áreas de distribución natural (Pruett-Jones 2021).
El Loro Barranquero (Cyanoliseus patagonus) es un psittaciforme neotropical que se distribuye principalmente en Argentina y en Chile (Rojas Martínez 2008, Masello et al. 2011) y se ha observado esporádicamente en Uruguay (Bucher y Rodríguez 1986). La abundancia del Loro Barranquero ha sufrido una considerable disminución desde principios del siglo XIX, que se adjudica a la pérdida y degradación de hábitat, a la captura para el mercado de mascotas, y a la persecución histórica por ser considerada plaga agrícola, aún sin evidencia científica que respalde este último escenario (Masello et al. 2006, 2011, Failla et al. 2008, Sánchez et al. 2016). Estos factores, en combinación con los requerimientos específicos del Loro Barranquero respecto de sitios de nidificación, de agua y recursos alimenticios, resultan en una distribución actual fragmentada y en variaciones geográficas significativas en su densidad, que puede resultar ocasionalmente abundante en algunos sectores de su área de distribución (Masello et al. 2011).
Las últimas estimaciones objetivas sobre la abundancia de Loro Barranquero en el extremo sur de la provincia de Buenos Aires fueron publicadas hace diez años por Grilli y colaboradores (2012), quienes destacaban la concentración de la especie en cercanías de la ciudad de Pedro Luro, Partido de Villarino, provincia de Buenos Aires. En los últimos años, se ha reportado un incremento aparente de su abundancia en otras ciudades y pequeñas localidades de esta región, acompañado de denuncias sobre daños en el tendido eléctrico y otros servicios, acumulación de suciedad en parques y paseos públicos y sonidos molestos, especialmente al atardecer, cuando los loros arriban a estas áreas urbanas para pernoctar en árboles o cableados, usando estos sitios como dormideros comunales (Sanchez Angonova 2019). El escenario de aparente incremento de las poblaciones de Loro Barranquero cobró relevancia regional, reflejándose en diferentes medios de comunicación que informaron aumentos de abundancia sin datos objetivos y sin considerar explicaciones alternativas, como modificaciones en el uso del hábitat o desplazamientos regionales de la especie (Bermúdez 2018, Diario La Tercera 2018, González 2018).
En ese contexto, y con el objetivo de aportar información que contribuya a clarificar la situación descripta, comparamos la abundancia del Loro Barranquero en el dormidero comunal de la ciudad de Bahía Blanca en los últimos nueve años y evaluamos las fluctuaciones estacionales en su abundancia.
métodosÁrea de estudio
La ciudad de Bahía Blanca se encuentra en el sudoeste de la provincia de Buenos Aires, Argentina (38°44'S, 62°16'O). De acuerdo con el censo realizado en 2010, la población cuenta con unos 301 572 habitantes. El Parque de Mayo es uno de los principales espacios verdes de la ciudad y cubre más de 60 ha rodeadas por barrios residenciales con predominio de construcciones de una planta o de pocos pisos (Fig. 1). El parque está forestado con ejemplares de Eucalipto Colorado (Eucalyptus camaldulensis), Ciprés (Cupressuss empervirens), Álamo Plateado (Populus alba), Pino Tos-quero (Pinus halepensis), Sófora (Styphnolobium japoni-cum), Palma Canaria (Phoenix canariensis) y Aguaribay (Schinus molle) (Speake 2015). Los loros utilizan desde hace décadas, y a lo largo de todo el año, un área de dormidero sobre los eucaliptos colorados de gran porte en este parque urbano (Pérez 2004). Cada tarde el Parque de Mayo recibe desde varios centenares a miles de loros, y, con excepción de un par de sitios muy menores, este dormidero concentra a casi la totalidad de los individuos en un radio de 20 km (Tella et al. 2014).
Diseño de estudio y censos de abundancia
En este estudio realizamos un total de 52 con-teos de loros en primavera, verano y otoño-invierno de los años 2014, 2018, 2019, 2020, 2021 y 2022. Entre febrero y diciembre de 2014 los censos fueron bimensuales. Desde julio de 2018 y hasta febrero de 2020, previo al cierre de los espacios públicos por la pandemia de Covid-19, los conteos se repitieron cada aproximadamente veinte días, y cada mes desde diciembre de 2021 hasta el 4 de julio de 2022. Contamos con la colaboración de más de 40 voluntarios rotativos que trabajaron en grupos de dos o tres, distribuidos en seis puntos fijos de observación ubicados a distancias de entre 300 y 1200 m en torno al dormidero. Cada grupo de observadores cubría una transecta relativamente perpendicular a las direcciones de arribo de los loros, de forma tal que todos ellos completaban un polígono de observación cerrado alrededor del parque (Canale 2015, Fig. 1). Registramos el número total de loros que volaron en dirección al dormidero comunal en franjas horarias consecutivas de 15 min de duración. Cada censo se extendió entre 45 y 60 min, desde el periodo previo a la puesta de sol hasta pasado el crepúsculo vespertino. Los grupos de observadores se mantenían comunicados y cesaban las tareas luego de pasados unos 15 minutos sin observaciones, lo que solía variar entre censos y resultaba en variaciones en su duración. Para evaluar las variaciones interanuales y estacionales del número de individuos que usaron el dormidero comunal de la ciudad de Bahía Blanca, comparamos los valores de abundancia de los censos mediante el test no paramétrico de Kruskal Wallis (Zar 1999). Los valores se expresan como promedios ± desvío estándar. Trabajamos con un nivel de significancia de 0.05.
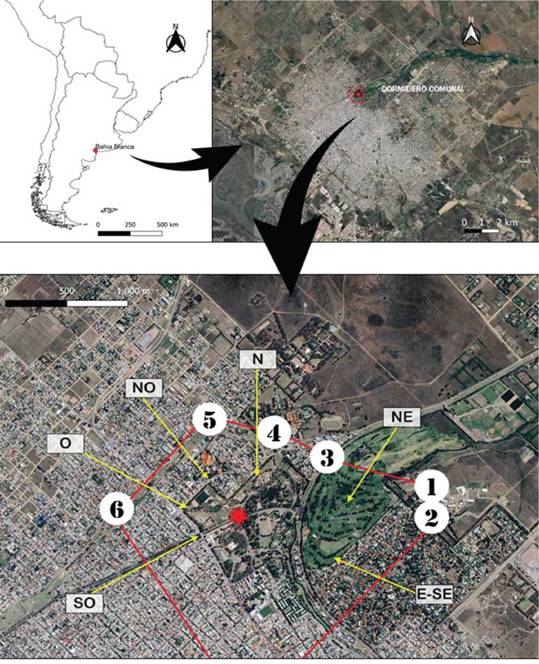
Figura 1. Dormidero de Loro Barranquero (Cyanoliseus patagonas) en la ciudad de Bahía Blanca, provincia de Buenos Aires, Argentina. La estrella marca el centro del área usada por la especie para pernoctar, los números señalan los puntos de observación de los conteos y las líneas rojas las transectas de intercepción. Las flechas indican además las direcciones de arribo de los loros.
Tabla 1. Número de ejemplares de Loro Barranquero que arribaron al dormidero del Parque de Mayo (Bahía Blanca) entre febrero de 2014 y julio de 2022. Entre paréntesis se indica el número de censos correspondiente a cada periodo.
Resultados
Detectamos diferencias marginalmente significativas en el número de loros que arribaron al dormidero comunal del Parque de Mayo a lo largo de todo el periodo de estudio (H = 11.03; gl = 5; P = 0.051). El mínimo de individuos (1767 loros) se registró en el censo del 31 de octubre de 2014 (primavera) mientras que el valor máximo (21 104 loros) se registró durante el conteo del 6 de agosto de 2019 (invierno,Tabla 1).
Las comparaciones interanuales no reflejaron un incremento significativo de la abundancia de loros entre las distintas primaveras (H = 3.85; gl = 3; P = 0.27), entre los veranos (H = 7.19; gl = 3; P = 0.065), ni entre las temporadas de otoño-invierno de los diferentes años estudiados, salvo para el otoño-invierno de 2019, que casi duplicó los valores correspondientes al mismo periodo del año 2014 (H = 6.02; gl = 3; P = 0.021).
La abundancia media de otoño-invierno fue de 10 693 ± 4248 loros (rango = 5082- 21 104 loros), la de la primavera de 3921 ± 1390 loros (rango = 1767 -6193 loros) y la del verano de 8813 ± 3108 loros (rango = 2684 - 12 972 loros; Fig. 2). En todos los años la abundancia siguió un mismo patrón estacional, con máximos en la temporadas de otoño-invierno y caídas en la primavera que resultaron estadísticamente significativas en 2014 y 2019 (P = 0.006) y marginalmente significativas para la primavera de 2018 (P < 0.06; Fig. 3).
Discusión
Este trabajo reúne estimaciones de abundancia de Loro Barranquero en un período de casi diez años. El número de loros que arribó al dormidero comunal en Bahía Blanca durante el otoño invierno de 2019 representa el máximo valor registrado hasta el momento en dicha ciudad, duplicando los valores correspondientes al mismo periodo del año 2014 en este mismo sitio, y superando en unas ocho veces al que reportan Grilli y colaboradores (2012) para un dormidero comunal en la ciudad de Pedro Luro. Este valor inusualmente alto coincidió con un aumento en la frecuencia de noticias locales publicadas por los medios de comunicación durante 2018, 2019 y 2020 que reportaron problemas relacionados con daños sobre instalaciones eléctricas, contaminación sonora, complicaciones sanitarias, aplicación de métodos para ahuyentar a los ejemplares de las áreas pobladas en distintas localidades del sudoeste de la provincia de Buenos Aires (Guercio 2019, La Nueva 2020), y con un reporte técnico sobre opciones para la mitigación de conflictos en la misma región (Sanchez Angonova 2019). Algunas notas periodísticas incluso reportaron interrupciones en el suministro eléctrico y eventos de tráfico ilegal de Loro Barranquero, en ambientes urbanos de otras provincias incluyendo Catamarca, Córdoba y San Juan (Puntal 2020, El An-casti 2021, Quiroga 2021).
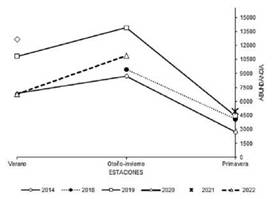
Figura 2. Abundancia estacional promedio de Loros Barranqueros arribando al dormidero comunal del Parque de Mayo, provincia de Buenos Aires, Argentina, entre 2014 y 2022.
Los resultados de nuestro trabajo y las observaciones menos sistemáticas citadas en el párrafo anterior parecerían apoyar la idea de un desplazamiento de loros hacia el dormidero urbano de la ciudad de Bahía Blanca, y eventualmente de otros centros urbanos de su área de distribución. Según los últimos censos realizados en la temporada otoño-invierno de 2022, esta situación se mantendría en la ciudad de Bahía Blanca, aunque con valores de abundancia ligeramente inferiores.
Resulta complejo postular cuál o cuáles podrían ser las causas de ese cambio, así como calificar su
carácter como temporario o permanente. En primer término, es muy razonable descartar un verdadero incremento en el tamaño poblacional de la especie, en tanto que los factores responsables de su retracción histórica no han hecho sino intensificarse en los últimos años. Las prácticas agrícolas son responsables de la pérdida de grandes extensiones de Monte nativo (Pezzola et al. 2004, Sánchez 2011, Zeberio 2012, Ga-bella 2015), de cuya vegetación autóctona dependen las poblaciones de Loro Barranquero y otras especies silvestres (Masello et al. 2011). Resulta mucho más probable, que el aumento en la abundancia de loros utilizando el dormidero comunal de Bahía Blanca en 2019 respecto de la situación cinco años antes, y los posibles incrementos en otras localidades de la zona reflejados en los medios de comunicación, obedezcan a desplazamientos regionales asociados, por ejemplo, con la búsqueda de los recursos alternativos o complementarios que les pudieran proveer las ciudades. Allí podrían encontrar sitios de pernocte y refugio particularmente seguros frente a los depredadores, como son las arboledas, redes de tendido eléctrico, luminarias y otras estructuras antrópicas (Masello 2021). El desmonte, las temporadas cíclicas de sequía y la consecuente baja disponibilidad de alimentos podrían estimular a las bandadas a buscar frutos y semillas de la flora urbana (Masello et al. 2006, Masello y Quillfeldt 2012), aprovechando además los granos
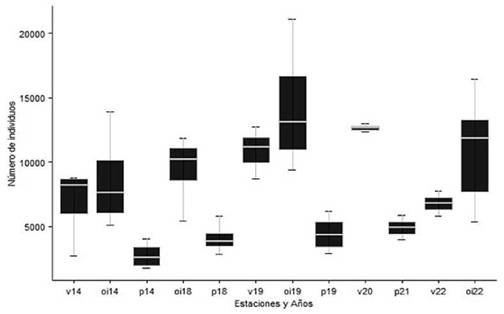
Figura 3. Número de Loros Barranqueros que arribaron al dormidero del Parque de Mayo, provincia de Buenos Aires, Argentina, entre febrero de 2014 y julio de 2022. En el eje X se indica cada año con sus respectivas estaciones, v: verano, p: primavera, o-i: otoño-invierno. Cada caja resume la información del total de censos desarrollados para la estación de cada año. Se muestran los valores máximo y mínimo y los cuartiles (primero, mediana y tercero). La línea blanca dentro de cada caja representa la mediana correspondiente.
de cereales derramados en los bordes de ruta y accesos a las ciudades (Sánchez et al. 2016). En particular durante las épocas de sequía, que resultan recurrentes en la región (Andrade et al. 2009, Casado y Campo 2019, Ferrelli 2020), los loros podrían aumentar su abundancia en áreas urbanas contiguas a zonas de regadío (Masello et al. 2011, 2015). Al respecto resulta interesante notar que el incremento local del número de loros que reportamos en este trabajo coincide con precipitaciones extremadamente bajas para la región (GBA, Gobierno de la provincia de Buenos Aires 2022). La rigurosidad de los efectos de la sequía y del desmonte son consistentes con eventos de mortandad masiva registrados durante las temporadas reproductivas 1998-1999 (Masello y Quillfeldt 2012) y 2020-2021 (Masello y Balbiano 2021), en la principal colonia reproductiva de la especie ubicada en el Balneario El Cóndor (Río Negro).
No debe descartarse en este punto la posibilidad de que la disponibilidad y/o la configuración de los remanentes de hábitats naturales para el Loro Barranquero estén alcanzando umbrales que disparen procesos demográficos o comportamentales (Fahrig 2002, 2013, Swift y Hannon 2010, Melo et al. 2018) y representen un nuevo estadio en la tendencia de declinación de la especie, aumentando la preocupación acerca de las perspectivas para su conservación.
A este escenario se suman las acciones de ahu-yentamiento desarrolladas en los dormideros de distintas localidades de la región, como Pedro Luro y Médanos en 2018, 2019 y 2020 (Infocielo 2018, La Arena 2020), que podrían influir sobre los desplazamientos de la especie y explicar, al menos en parte, el notable incremento que comenzó a observarse en el dormidero de la ciudad de Bahía Blanca en 2019. En este punto es importante destacar el desafío que representa la conservación de una especie que, pese a su retracción histórica, muestra variaciones regionales y locales de abundancia que la hacen ver como un ave abundante y con poblaciones en aumento frente a la población general, situación que se refleja claramente en los medios de comunicación locales.
Resulta particularmente interesante observar cómo los cambios estacionales de abundancia de la especie en Bahía Blanca se complementan con las fluctuaciones registradas en el Balneario El Cóndor, donde el número de loros alcanza sus valores máximos durante la temporada reproductiva (Masello et al. 2006). En esa misma época del año, Bahía Blanca y el resto de las ciudades y localidades del sudoeste bonaerense, donde las barrancas naturales y los sitios antrópicos aptos para anidar resultan limitantes, registran un descenso en la cantidad de individuos (Tella et al. 2014, Canale 2015).
Desde el punto de vista metodológico, los con-teos en dormideros están asociados con una serie de limitaciones a la hora de estimar la abundancia (Bi-bby et al. 1992, Casagrande y Beissinger 1997, Dénes et al. 2018, Zulian et al. 2020, 2022). Para minimizar los posibles efectos que estos pudieran tener sobre nuestros resultados, tomamos una serie de decisiones que podrían ayudar a estandarizar esfuerzos similares con esta y con otras especies de comportamiento y características similares. Por un lado, ubicamos los puntos de observación en áreas abiertas, de alta visibilidad, claramente delimitadas a partir de elementos salientes del paisaje, de manera de reducir el riesgo de dobles conteos entre grupos de observadores. Evitamos días de lluvia y de viento intenso para eludir condiciones climáticas que pudieran reducir la visibilidad y/o interferir en el horario de arribo de los loros. Ajustamos el inicio de los censos mediante observaciones en los días previos, comenzando 15 min antes de la aparición de los primeros individuos. Para evitar errores por diferencias en el entrenamiento de los observadores, cada grupo contó con al menos un observador con experiencia previa y en la mayoría de las ocasiones los conteos fueron realizados por dos o más voluntarios por punto.
Más allá de eso, el tamaño de los animales (42 cm; Narosky e Yzurieta 2010), el vuelo bajo (entre 15 y 50 m por encima de los observadores) y su comportamiento ruidoso en vuelo, facilitó nuestras tareas y ayudó a evitar posibles omisiones. Por otra parte, el foco de nuestro trabajo era determinar variaciones estacionales e interanuales en la abundancia de la especie, por lo que el esfuerzo se concentró en la precisión de los conteos, más que en su exactitud.
Los resultados obtenidos en este trabajo destacan la relación entre distintos núcleos de distribución de la especie y la consecuente necesidad de emprender acciones de conservación a nivel regional. Se vuelve evidente, además, la importancia de mantener evaluaciones objetivas de la densidad de la especie que comprendan ambientes naturales y antrópicos en temporadas de cría y de invernada para discriminar entre cambios en el uso del territorio y variaciones reales en su abundancia poblacional. Se observa también la importancia de completar la identificación de los dormideros de la región, estandarizando los métodos de con-teo para aumentar la eficiencia de estas estimaciones. Finalmente, y si bien está clara la imperiosa necesidad de conservar y restaurar los ambientes naturales que proveen recursos clave para esta especie, estos esfuerzos deben complementarse con estrategias de convivencia que reduzcan los conflictos que derivan de su asociación con los centros urbanos.
agradecimientosAgradecemos a los voluntarios por su compromiso y participación en los censos de abundancia que permitieron el desarrollo de esta investigación: Agustín Álvarez, Andrés Cervino, Natalia Contreras Salvatico, Xoana Fernandez, Franco Farías Toledo, Elias Forconi, Marcela Frechero, Laura Fritz, Carolina Giménez, Silvana Giunchiglia, Jonathan Giuliani, Sebastián Giunti, Ana Belén Gómez Llopis, Laura Haag, Braian Janco-vich, Jesica Liberati, Gabriela León, María Victoria Le-dezma Martínez, Joscelyn Locati, Fernando López, Pamela Malmoria, Leandro Marban, Evangelina Mattos, Jessica Montiel, Paula Monzón, Andrés Miguel Oses Ríos, Laura Otero, Marina Páez, Rodrigo Emanuel Pérez, Brenda Pizzano, Abel Pontoriero, Nicole Quaglia, Carla Rayen Barrera, Martin Saavedra, Sabrina Salas, Alberto Scorolli, Nicolás Segatori, Romina Tejerina, Camilo Traini, Paula Valverde y Germán Villalba. Al COA Loica Pampeana por facilitarnos los binoculares para los conteos. A los tres revisores que realizaron valiosos aportes a este manuscrito. Este trabajo fue financiado con fondos del CONICET y de la Universidad Nacional del Sur.
Recibido: 1 de julio de 2022
Aceptado: 8 de noviembre de 2022

